











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El cultivo de chile verde representa para México una producción de 3.24 millones de t, donde el 58.36% se produce en los estados de Sinaloa (757 769.051), Chihuahua (682 084.871) y Zacatecas (450 098.95 t) (SIAP 2020). El principal factor que limita la producción y ocasiona o genera bajos rendimientos del chile es la pudrición del sistema radicular, la cual está asociada con Rhizoctonia solani, Fusarium oxysporum, Verticillium sp., Phytophthora sp. y Pythium sp. (Pérez et al. 2017, Reyes et al. 2021). En la actualidad, el manejo de la enfermedad se basa en el control químico y el uso de tecnologías que conducen a la inestabilidad en los ecosistemas naturales, además, que genera resistencia a moléculas activas de fungicidas de diferentes mecanismos de acción (Sang et al. 2018, De Ramón et al. 2019). Una alternativa factible es el uso de diferentes especies de Trichoderma que interactúan en beneficio de las plantas, al mejorar su crecimiento y desarrollo, además de su alta actividad antagónica a través de la competencia espacial y micoparasitismo (Salazar et al. 2015). También se han reportado enzimas y efectores que se involucran en el reconocimiento y la degradación de la pared celular del huésped frente a R. solani, P capsici, Sclerotinia sclerotiorum y Botrytis cinérea (Atanasova et al. 2018, Andrade et al. 2019). Por lo anterior el objetivo del trabajo fue determinar la compatibilidad entre cuatro especies de Trichoderma y su efecto en la incidencia causada por Fusarium oxysporum y Rhizoctonia solani y en el rendimiento del cultivo del chile (Capsicum annuum L.).
Materiales y métodos
Ubicación de los experimentos
El ensayo se realizó en la Universidad Autónoma Agraria Antonio Narro (UAAAN) en Saltillo, Coahuila; en el Laboratorio de Microbiología del Departamento de Parasitología y el trabajo de campo en una parcela experimental del campo El Bajío de la misma Universidad, donde se reporta la presencia de marchitez del chile causada por Fusarium oxysporum y Rhizoctonia solani (Espinoza et al. 2019).
Material biológico
Los microorganismos antagónicos fueron proporcionados por el cepario de Microbiología de la UAAAN. Las cepas evaluadas fueron Trichoderma asperellum (TA), T harzianum (THZ), T lignorum (TL) y T. yunnanense (TY); las cuales fueron activadas en el medio de cultivo papa-dextrosa-agar (PDA) suplementado con 3 g L-1 de extracto de malta.
Compatibilidad entre especies de Trichoderma
Las cepas de Trichoderma spp. se activaron en cajas Petri de 90 mm con medio de cultivo Papa-Dextrosa-Agar (PDA), transcurridos 10 días se realizaron confrontaciones entre dos, tres y cuatro especies. Se establecieron 8 tratamientos, para confrontar dos especies se sembraron en puntos equidistantes (TA-TL, TA-THZ, TA-TY y TY-TL) con tres especies al sembrarlas en los puntos de intersección de un triángulo equilátero (TA-TLTHZ, TY-TA-TL y TA-TY-THZ) y las cuatro especies se sembraron en los cuatro puntos cardinales (TA-TL-THZ-TY) de la caja Petri con el medio PDA y se incubaron a 27 ± 2 °C. Se realizaron observaciones cada 12 h, una vez hubo contacto entre las especies confrontadas se midió cada especie confrontada y cada una de ellas por separado, para lo cual, se utilizó un Vernier digital y se determinó la compatibilidad usando la metodología de Sueke et al. (2010). Ocho días después de la siembra se contabilizaron las esporas con una cámara de Neubauer en un microscopio compuesto Motic BA210E y se midió el ancho de barrera en el punto de contacto entre las especies de Trichoderma confrontadas.
Ensayo en campo
Se sembraron semillas de chile serrano de la variedad Tampiqueño 74 en charolas de poliestireno, transcurridos 40 días después de la siembra (dds), plántulas de 10 cm de altura se trasplantaron a doble hilera en camas de 1.5 m, con riego por goteo y acolchado plástico. La fertilización se realizó antes de la siembra, aplicando al fondo del surco la dosis de 600 kg ha-1 de la mezcla física T-17 (17-17-17), la cual tenía una composición de fosfato monoamónico granulado (11-52-00), urea (46-00-00) y nitrato de potasio (12-00-46). Con base en los resultados de compatibilidad in vitro se evaluaron los tratamientos de especies de Trichoderma por separado (TL, THZ, TA), dos (TA-TL, THZ-TL y THZ-TA), tres (THZ-TA-TL) y un testigo absoluto. Las aplicaciones se realizaron al drench a los 7, 21, 51 y 81 dds, con una solución de conidias de 1X106. El experimento se estableció en un diseño de bloques al azar con tres repeticiones. Transcurridos 85 días dds se evaluaron las variables producción por planta, número de frutos por planta, además, los parámetros de incidencia y severidad de la enfermedad como se describe en la investigación de Espinoza et al. (2019).
Resultados y discusión
Compatibilidad de especies de Trichoderma
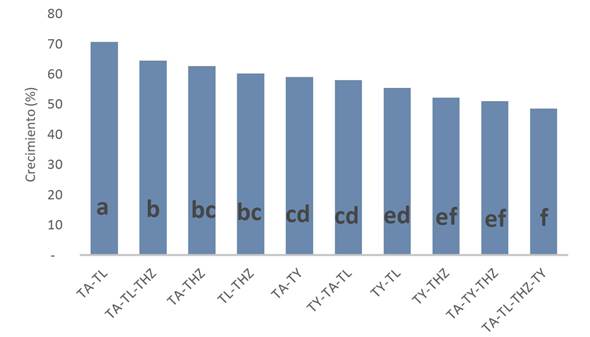
Al confrontar las diferentes cepas de Trichoderma, se encontró que T. lignorum (TL) produjo altas concentraciones de conidios por mL (4.42 x 107), seguido por la combinación TA-TL (2.04 x 107) y TY-TL (1.46 x 107). Las mejores combinaciones con tres especies fue TA-TY-THZ y TA-TL-THZ en concentraciones de 9.92 x 106 y 9.28 x 106, respectivamente; además se observó que al combinar las cuatro cepas la esporulación se redujo significativamente (Tabla 1). La compatibilidad en el crecimiento (Figura 1) muestra que existe diferencia estadística significativa (p < 0.05), donde TA-TL, TA-TL-THZ, TA-THZ y TL-THZ mostraron compatibilidad del crecimiento del 60 al 71%. Al realizar las mediciones y observaciones de la barrera formada cuando se confrontan las diferentes especies de Trichoderma (Figura 1) se pudo constatar que existe compatibilidad de TA-TL, TA-THZ y TA-TL-THZ, con ancho de barrera de 0.05, 0.06 y 0.08 cm, respectivamente. Lo que demuestra que TA-TL y TA-TL-THZ son compatibles por su buen comportamiento en las variables de esporulación, compatibilidad en el crecimiento y sin efecto antagónico al crecer en el mismo espacio. La condición de la formación de barrera de defensa, competencia por el espacio y esporulación, indican que el efecto encontrado se asocia a la producción de metabolitos secundarios, además el contenido mineral del medio de cultivo y la afinidad entre especies puede mantener su capacidad de esporulación (Ortuño et al. 2013). El desarrollo micelial de los microorganismos que comparten un mismo espacio responden a principios ecológicos, donde la interacción funcional se observa cuando la contribución de un miembro de la comunidad a una función a nivel comunitario depende de la presencia o ausencia de otras especies (Sánchez et al. 2019). También se sabe que microorganismos compatibles mejoran el suelo desde su composición físico-químico, contenido de materia orgánica, pH, conductividad eléctrica, nitrógeno, fósforo, potasio y calcio (Villacís et al. 2016), además promueven el crecimiento vegetal, controlan enfermedades y se aumenta el rendimiento de los cultivos (Cano 2011).
Tabla 1 Esporulación en placa de especies de Trichoderma solos y en combinación.
| Combinación | Esporas mL-1 | Combinación | Esporas mL-1 |
| TL | 4.42 x 107 a * | TA-THZ | 5.45 x 106 def |
| TA-TL | 2.04 x 107 b | TA-TY | 5.38 x 106 ef |
| TY-TL | 1.46 x 107 bc | TY-TA-TL | 5.02 x 106 ef |
| TL-THZ | 1.39 x 107 bcd | TA-TL-THZ-TY | 4.94 x 106 ef |
| TA | 1.29 x 107 bcde | TY-THZ | 4.63 x 106 ef |
| TA-TY-THZ | 9.92 x 106 cdef | THZ | 2.72 x 106 f |
| TA-TL-THZ | 9.28 x 106 cdef | TY | 1.66 x 106 f |
*Especies de Trichoderma solas o en combinación producen la misma cantidad de conidios estadísticamente (P > 0.05). TA: Trichoderma asperellum, THZ: T. harzianum, TL: T. lignorum, y TY: T. yunnanense
Incidencia y severidad de la marchitez del chile
En la incidencia de la enfermedad (Tabla 2) se encontraron diferencias estadísticas marcadas (p < 0.05), expresando el testigo altos porcentajes (47.69%) de la enfermedad. Mientras que el tratamiento THZ-TA-TL expreso baja incidencia (13.67%), pero la severidad fue estadísticamente igual, lo que sugiere que la aplicación de las tres especies de Trichoderma protege a la planta de la infección, pero no tiene efecto en plantas enfermas. La incidencia y severidad de la marchitez del chile se reducen con la aplicación de diferentes especies de Trichoderma, al respecto, Sánchez et al. (2019) indican que al confrontar a especies de Trichoderma spp. con Pythium ultimum hay antagonismo in vitro del fitopatógeno, además, en condiciones de campo se reduce la sintomatología en raíz y área foliar. La combinación de micoorganismos compatibles desencadenan e incrementan la respuesta de defensa en la planta comparado cuando son suministrados microbios solos (Rodríguez et al. 2019, Morato y Cardona 2021), por lo que esta alternativa de control biológico suprime eficazmente los patógenos y es amigable con el medio ambiente (Nishad et al. 2020).
Tabla 2 Comportamiento de plantas de chile en producción de frutos en campo bajo diferentes tratamientos de especies de Trichoderma solos o en combinación aplicados al drench.
| Tratamiento | Pi | EE | DPi | EE | Ps | EE | DPs | EE | Pp | EE | Ipp | EE | Fp | EE | IFp | EE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| THZ-TA-TL | 13.67b* | 1 | 71.34a | 7 | 9.67a | 1 | 57.53a | 5 | 737.30ab | 52 | 70.55a | 5 | 113.67ab | 12 | 72.80a | 18 |
| THZ | 34.52ab | 4 | 29.22a | 4 | 16.43a | 5 | 39.20a | 4 | 630.00ab | 58 | 47.54a | 6 | 97.44ab | 7 | 48.14a | 10 |
| TA | 32.74ab | 5 | 31.35a | 1 | 7.50a | 1 | 58.88a | 7 | 678.00ab | 74 | 63.64a | 15 | 99.22ab | 16 | 59.21a | 10 |
| TL | 20.97ab | 1 | 56.04a | 11 | 9.12a | 1 | 50.02a | 13 | 754.20ab | 37 | 74.75a | 10 | 112.56ab | 13 | 71.92a | 12 |
| THZ-TA | 32.41ab | 1 | 33.66a | 3 | 10.65a | 1 | 41.62a | 2 | 478.00ab | 33 | 14.11a | 2 | 70.89ab | 6 | 14.17a | 2 |
| THZ-TL | 28.91ab | 1 | 55.98a | 2 | 6.05a | 1 | 6.85a | 4 | 769.80ab | 45 | 78.02a | 5 | 97.67ab | 7 | 48.95a | 6 |
| TA-TL | 21.76ab | 2 | 55.98a | 2 | 7.63a | 1 | 58.14a | 4 | 802.20a | 46 | 86.21a | 3 | 116.22a | 14 | 79.14a | 5 |
| TESTIGO | 47.69a | 6 | - | - | 18.24a | 3 | - | - | 434.40b | 26 | - | - | 65.78b | 5 | - | - |
Pi: porcentaje de incidencia de la enfermedad; EE: error estándar; Dpi: disminución del porcentaje de incidencia de la enfermedad; Ps: porcentaje de severidad de la enfermedad; DPs: disminución del porcentaje de severidad de la enfermedad; Pp: producción por planta; IPp: incremento de producción por planta; Fp: frutos por planta; IFp: incremento de frutos por planta; TA: Trichoderma asperellum; THZ: T. harzianum; TL: T. lignorum; TY: T. yunnanense. *Tratamientos con letras diferentes entre columnas indican diferencias estadísticas de acuerdo con la prueba de Tukey (P < 0.05).
Producción y número de frutos por planta
En la producción de frutos por planta se encontraron diferencias estadísticas significativas (p < 0.05), presentando la mezcla TA-TL la mayor producción (802.2 g planta-1), seguido de THZ-TL (769.8 g planta-1), con respecto al testigo los tratamientos incrementaron la producción en 86 y 78%, ya que el testigo tuvo un rendimiento de 434.4 g planta-1. Para el número de frutos por planta, se encontraron diferencias significativas (p < 0.05), presentando las mezclas TA-TL 116.22, THZ-TA-TL 113.67 y TL 112.56, mientras que el testigo tuvo 65.78 frutos por planta. Pero los incrementos de producción fueron estadísticamente iguales, pero se encontraron incrementos del 14 al 79% con respecto al testigo (Tabla 2). Al respecto, Alejo et al. (2015) comprobaron que los aislamientos de Trichoderma spp. provenientes de patosistemas silvestres y agrícolas tuvieron un efecto promotor de crecimiento en plantas de Capsicum chinense Jacq., por otra parte, estudios con chile dulce demuestran que la aplicación de T. asperellum estimula el crecimiento de las plántulas en la etapa de almácigo, lo que mejoró la elongación de la raíz, la biomasa de la raíz y el área foliar (Brenes et al. 2019). Lo que se puede atribuir a que las especies de Trichoderma que interactúan en la rizosfera sintetizando hormonas en la raíz de la planta, lo que puede interferir con el desarrollo y alterar de forma positiva el desarrollo de las plantas (Nieto et al. 2017, Li et al. 2019). También se ha comprobado que hay una relación armónica y ecológicamente compatible al aplicar microorganismos con productos bioactivos, ya que estos permiten un adecuado estado nutricional de las plantas y son una alternativa a explotar en el sector agrícola (Terry et al. 2005).
Se encontró compatibilidad de Trichoderma lignorum con T. asperellum y T. harzianum, en laboratorio y campo, ya que en condiciones in vitro no se antagonizan, comparten espacio y permiten la esporulación. Se tiene una respuesta favorable a la inoculación de Trichoderma lignorum y sus combinaciones con T. asperellum y T. harzianum, con evidencia de aumento del rendimiento y sanidad de las plantas de chile serrano Tampiqueño 74 sensible a la enfermedad en condiciones de campo.

