











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La producción mundial de tomate (Solanum lycopersicum L.) en 2019 fue de 180 766329 t (FAOSTAT 2021) y es el segundo cultivo de hortalizas más importante a nivel mundial después de la papa (Costa y Heuvelink 2018). Con respecto a la variedad Mariela de tomate, tiene altos rendimientos, frutos grandes y un comportamiento adecuado frente a los virus, Alternaría solani (Cooke) Wint. y Phytophthora infestans (Mont.) de Bary (Moya et al. 2000). De acuerdo con Al-Qurainy (2009) hay aproximadamente 400 especies de plantas de al menos 45 familias que son hiperacumuladoras de metales. Con respecto al tomate, es una planta que se ha utilizado en algunos estudios como planta hiperacumuladora, para la descontaminación de suelos con alta concentración de metales (Kumar et al. 2009, Bizily et al. 1999), ya que acumula hasta alrededor de 100 mg kg-1 peso seco de Cd, hasta 1 000 mg kg-1 peso seco de Ni, Cu, Co, Cr y Pb, y hasta 10 000 mg kg-1 peso seco de Zn y Mn (Bizily et al. 1999).
El plomo (Pb) es uno de los metales pesados más tóxicos incluso en concentraciones muy bajas que puede acumularse en diferentes partes induciendo la inhibición del crecimiento de las raíces, crecimiento subdesarrollado, ennegrecimiento del sistema radicular y clorosis (Sharma y Dubey 2005). Al respecto, Akinci et al. (2010) explica que en plantas de tomate la elongación de la raíz y altura de la planta; biomasa fresca y seca de raíces, brotes y hojas; área foliar fueron negativamente afectadas por el aumento de las concentraciones de plomo (75, 150 and 300 mg L·1).
El sistema que regula a las especies reactivas de oxígeno (EROs) para mantenerlas a un nivel no dañino en las células vegetales incluye una red de antioxidantes no enzimática y enzimática (Sachdev et al. 2021), esta última incluye enzimas como la superóxido dismutasa (SOD), glutatión reductasa (GR), catalasa (CAT), entre otras (Hasanuzzaman et al. 2012, Kaur et al. 2019). Teniendo en cuenta que el nivel de metales pesados en los suelos agrícolas aumenta año tras año y que el tomate es una hortaliza muy consumida en el mundo, además del poco conocimiento que existe acerca de las respuestas de las enzimas del estrés oxidativo a nivel bioquímico y molecular en plantas de tomate que crecen en suelos contaminados con plomo, el objetivo de este trabajo fue analizar la actividad de algunas enzimas del estrés oxidativo a nivel bioquímico y molecular en plantas de tomate. cv. Mariela con diferentes concentraciones de PbAc2, lo que permitirá una mejor comprensión de esas respuestas en este cultivo.
Materiales y métodos
Las semillas de tomate cv. Mariela se sembraron en bandejas que contenían una mezcla 1: 1 (v / v) de vermiculita suplementada con NPK (nitrógeno, fósforo y potasio) 10:10:10 (g) y un producto comercial (Plantmax HT Eucatex, Sao Paulo, Brasil) al 2% (p/v) y se colocaron en condiciones de invernadero en la Escuela Superior de Agricultura de la Universidad de Sao Paulo, Brasil. Las plántulas se trasplantaron una vez que aparecieron las primeras hojas verdaderas a macetas de 1 L (unidad experimental) con arena y tres concentraciones de Acetato de plomo (PbAc2; 0, 50 y 100 mg kg-1) con tres repeticiones cada una. Estas concentraciones de plomo se utilizaron considerando que las concentraciones de este metal son 40, 70 y 100 mg kg-1 para suelos arenosos, limosos y arcillosos, respectivamente (BMJV 2017).
A las plantas de tomate se les aplicaron 50 mL de las soluciones de PbAc2 aplicándosele a cada maceta 0, 50 o 100 mg kg-1 de PbAc2 dependiendo del tratamiento con una frecuencia de dos veces por semana a partir del día del trasplante hasta el inicio de la fase reproductiva (16 veces en 60 días); cuando las aplicaciones fueron completas, se recolectaron hojas y tallos representando la fase vegetativa de crecimiento. Terminada la recolección, los órganos vegetativos se lavaron con agua desionizada para almacenarse por separado a -80 °C hasta su análisis.
Determinación de la actividad de las enzimas antioxidantes
Para el estudio de la actividad enzimática, las muestras se manejaron a 4 °C. Las muestras de hojas y tallos por separado se homogeneizaron en un mortero con tampón de potasio 100 mM (pH 7,5) que contenía ácido etilendiaminotetraacético (EDTA) 1 mM, DL-ditiotreitol 3 mM y 5% (p/v) de polivinilpirrolidona en proporción de 2: 1 (volumen de tampón / peso fresco); luego se centrifugaron 30 min a 10.000 x g y el sobrenadante se almacenó en alícuotas a -80 °C (Azevedo et al. 1998).
La actividad de la GR (EC 1. 8. 1. 7) se analizó por espectrofotocolorimetría a 30 °C en una mezcla con 3 mL de tampón de fosfato de potasio 100 mM (pH 7.5) con ácido nitrobenzoico 1 mM, GSSG (disulfuro de glutatión) 1 mM, NADPH 0,1 mM (fosfato de dinucleótido de nicotinamida y adenina en forma reducida) y finalmente se añadió el extracto vegetal. El valor de reducir el GSSG se controló mediante el aumento de la absorbancia a 412 nm durante 2 minutos (Gomes-Junior et al. 2006). La actividad de la GR se determinó en μmol min mg-1 de proteína.
La actividad de la SOD (EC 1.15.1.1) se determinó monitoreando la inhibición de la reacción fotoquímica del nitro azul tetrazolio (NBT) (Beyer y Fridowich 1987), donde una unidad de actividad SOD se definió como la cantidad de enzima necesaria para causar el 50% de inhibición de la reducción del NBT monitoreada a 560 nm. La actividad de la SOD se midió en μmol min mg-1 de proteína. El análisis de la actividad de las enzimas en estudio se realizó con un espectrofotocolorímetro modelo Ray Leigh UV-2601.
Análisis de la concentración de las proteínas solubles totales
La concentración de las proteínas solubles totales se analizó por el método de Bradford (1976) con albúmina bovina (BSA) como estándar.
Análisis estadístico
El diseño experimental utilizado fue bloques al azar con tres concentraciones de Pb (0, 50 y 100 mg kg-1 PbAc2) y se realizaron tres repeticiones con 10 unidades experimentales en total. Se utilizó un ANOVA de clasificación simple y una prueba de Tukey (p ≤ 0.05) para comparar las medias entre los tratamientos. Se utilizó el programa estadístico SAS versión 9.4.
PCR en Tiempo Real
Un mini kit de plantas GeneEluteTM (Sigma-Aldrich®) se utilizó para la extracción del ARN total. La concentración del ácido nucleico se determinó con un espectrofotómetro NanoDrop (ND-1000, Thermo Scientific, EE. UU.). La síntesis del ADNc se llevó a cabo con el sistema SuperScriptTM (Invitrogen life technologies) y se llevó a cabo una PCR (Thermocycler Applied Systems 2720, EE. UU.) con un ciclo a 42 °C durante 5 min, 50 °C durante 50 min y 70 °C durante 15 min.
Se estudió la expresión de tres genes involucrados en las respuestas a estrés abiótico en cultivos de interés (IFR, TCTP y SOD). La PCR en tiempo real (RT-PCR) se realizó con cebadores que fueron diseñados con el software Primer Express 2.00 (Applied Biosystems) partiendo de secuencias recuperadas de la base de datos NCBI (Centro Nacional de Información Biotecnológica). En el caso del gen TCTP, se seleccionó de una biblioteca de tabaco de ADNc (Hai-Tao et al. 2005) porque su papel comienza a ser informado como relevante en el metabolismo de metales (Tabla 1). La RT-PCR se realizó utilizando Platinum® SYBR® Green qPCR Super MIX-UDG (Invitrogen), con placas de 48 pocillos. El programa de PCR incluyó un ciclo a 95 °C durante 15 min, 40 ciclos a 95 °C durante 15 min, 60 °C durante 60 min, un ciclo a 95 °C durante 15 min y 60 °C durante 60 min. Para el análisis de los datos generados se utilizó una cuantificación comparativa CT (ΔΔCT) utilizando el software StepOne.
Tabla 1 Diseño de los cebadores para el PCR en Tiempo Real.
| Largo (pb) | Tm | Secuencia del cebador | |
|---|---|---|---|
| SOD | |||
| Cebador Izquierdo | 20 | 59.93 | 5’ ATGATGGAACTGCCACCTTC 3’ |
| Cebador Derecho | 20 | 59.75 | 5’ TAGCAGCTTTAGCCCTGGAG 3’ |
| IFR | |||
| Cebador Izquierdo | 20 | 58.65 | 5’ TGCTCAGTACATGTGGGTCA 3’ |
| Cebador Derecho | 20 | 60.16 | 5’ GGATTCTCTTGGCATCCTGA 3’ |
| TCTP | |||
| Cebador Izquierdo | 20 | 60 | 5’ AGCTGGTCCCTCGTTTTCTT 3’ |
| Cebador Derecho | 20 | 60 | 5’ GCACCTTGAACAACCCACTT 3’ |
| Ubiquitina | |||
| Cebador Izquierdo | 21 | 58 | 5’ AGAGGAAGGGTTTGAAGCAGG3’ |
| Cebador Derecho | 21 | 59 | 5’ TTTGCCAGCTTGAACATCCAT3’ |
Resultados y discusión
El análisis de las proteínas solubles totales a diferentes concentraciones de PbAc2 mostró diferencias significativas en las hojas (Figura 1a) y en los tallos (Figura 1b ) de las plantas de tomate cv. Mariela. Este parámetro en las hojas fue menor a 50 mg kg-1 donde hubo 68% menos proteína que en el control, y a 100 mg kg-1 hubo 58% menos proteína que en el control bajo la influencia de PbAc2, como se muestra en la Figura 1A . Este resultado podría deberse a que la fotosíntesis se vio afectada, específicamente la máquina fotosintética por su relación con las proteínas. Al respecto Sharma y Dubey (2005) informan que el proceso de fotosíntesis se ve muy afectado por la toxicidad del Pb, además que el contenido de proteínas disminuye cuando se acumula el Pb (hojas) y a medida que la concentración del metal aumenta (Mohan y Hosetti 1997).
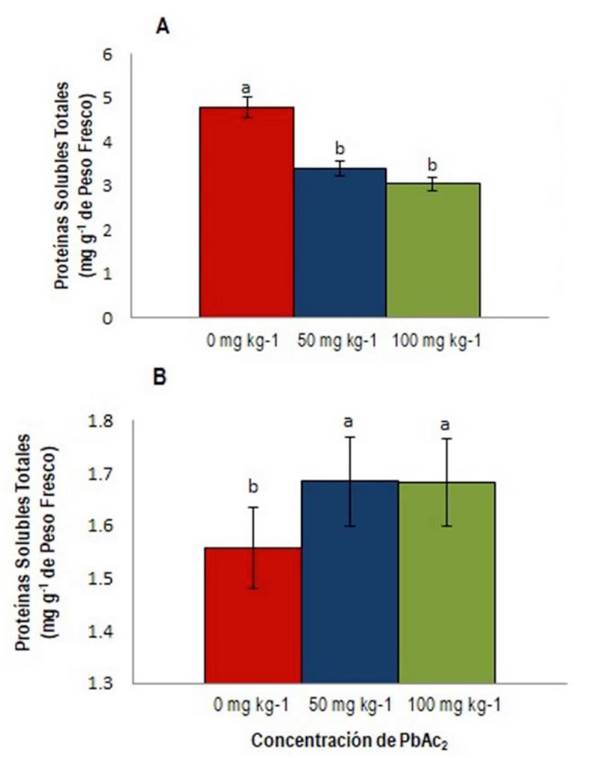
Figura 1 Proteínas Solubles Totales (mg g-1 de PF); A) Proteínas en hojas; B) Proteínas en tallos de S. lycopersicum cv. Mariela. Letras diferentes indican diferencias significativas (p ≤ 0.05).
En cuanto al aumento en los tallos de las proteínas solubles totales en las plantas tratadas con PbAc2 esto podría deberse a una saturación en los mecanismos de retención del Pb en el tallo, teniendo en cuenta que es un órgano de transporte por lo cual su capacidad como reservorio del metal es finita (Abreu et al. 2016).
La actividad de la GR con los tratamientos utilizados fue mayor (133. 3%) en hojas a 50 mg kg-1 sin diferencias significativas entre el control y 100 mg kg-1 de PbAc2 (Figura 2a ). Los valores encontrados difieren de los informados por Usman et al. (2020) en en Tetraena qatarensis donde la actividad de la GR en toda la planta aumentó con la concentración de Pb (0, 25, 50 y 100 mg L-1). La menor actividad de la GR encontrada en las hojas a 100 mg kg-1 de PbAc2 no difiere significativamente del control pudiendo implicar la inhibición de su actividad a esta concentración. Normalmente en condiciones de estrés la actividad de la enzima debe aumentar como parte de los procesos antioxidantes (Gill et al. 2013). Este resultado coincide con el obtenido por Khan et al. (2020) en plantas de algodón (Gossypium hirsutum L.) tratadas con 0 y 500 μM de Pb donde en las hojas medias no hubo diferencias significativas entre el control y el tratamiento de glutatión (GSH) más Pb.
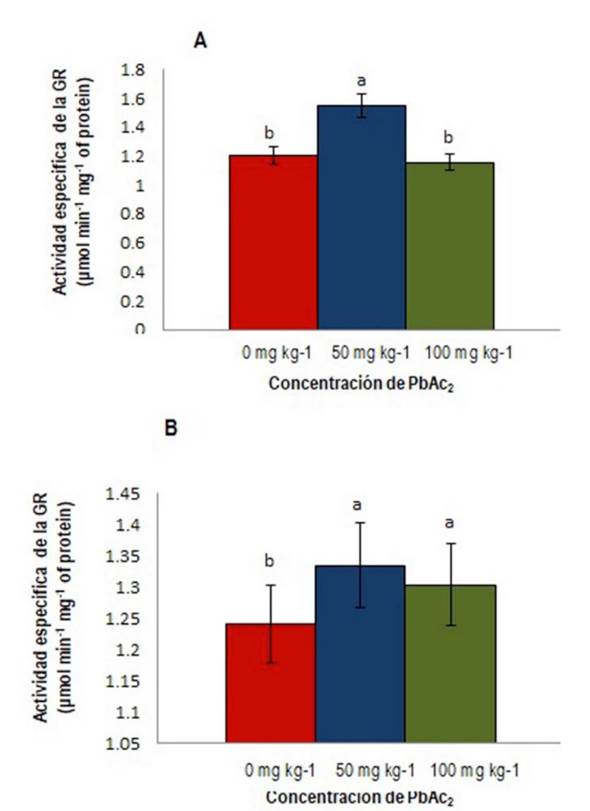
Figura 2 Actividad específica de la enzima GR (µmol min-1 mg-1 de proteína); A) GR en hojas, B) GR en tallos, de S. lycopersicum cv. Mariela. Letras diferentes indican diferencias significativas (p ≤ 0.05).
En los tallos, la actividad de la enzima aumentó un 108.9% a 50 mg kg-1 y un 105.6% a 100 mg kg-1 de PbAc2 (Figura 2b ). El Pb en el suelo se mueve desde las raíces hasta las hojas, pasando por el tallo en su camino hacia las hojas; en los tallos la GR comienza su función como enzima antioxidante al eliminar el H2O2 que se produce principalmente en los cloroplastos para mantener el estado redox de la célula (Asada 1992), lo que pudiera explicar su mayor actividad con las concentraciones de PbAc2 utilizadas.
La actividad de la SOD fue significativamente menor al compararla con el control bajo las dos concentraciones de PbAc2 en ambas partes de la planta estudiadas (Figura 3 a, b ). La disminución de la actividad de la SOD en presencia de las dos concentraciones de PbAc2 utilizadas y las dos partes de la planta analizadas coincide con los resultados obtenidos en plántulas de manglar (Bruguiera gymnorrhiza) (Tao et al. 2012), esquejes de J. curcas (Shu et al. 2011) y en plantas de cáñamo (Xia et al. 2019). Según Hasanuzzaman et al. (2012), la SOD es la enzima que actúa como la primera línea de defensa porque dismutasa el radical superóxido (O2 -) convirtiéndolo en H2O2. La menor actividad de esta enzima en esta investigación podría indicar una disminución de la función de dismutar el radical superóxido. El resultado encontrado en la actividad de la SOD por Machín-Suárez et al. (2017) en el tomate cv. Amalia bajo las mismas concentraciones de PbAc2 fue diferente al resultado de este estudio, lo que indica que en una misma especie de planta las respuestas enzimáticas varían entre cultivares. De manera general la actividad de ambas enzimas en la variedad de tomate Mariela tuvieron un comportamiento diferente pues la GR mostró una mayor actividad en hojas en la concentración de 50 mg kg-1 de PbAc2 y en tallos aumentó en ambas concentraciones mientras que la SOD disminuyó en ambos órganos y en las dos concentraciones estudiadas. Aunque las dos son enzimas antioxidantes están asociadas con la regulación de las EROs en plantas, cada una tiene una función muy diferente lo que explica este comportamiento. El estrés abiótico provocado por la aplicación de diferentes concentraciones de Plomo, inhibió la actividad enzimática de la SOD y estimuló la actividad enzimática de la GR en hojas y tallos, lo que mejoró el mecanismo de protección basado en la regulación de las actividades enzimáticas y compartimentación de ambas enzimas.
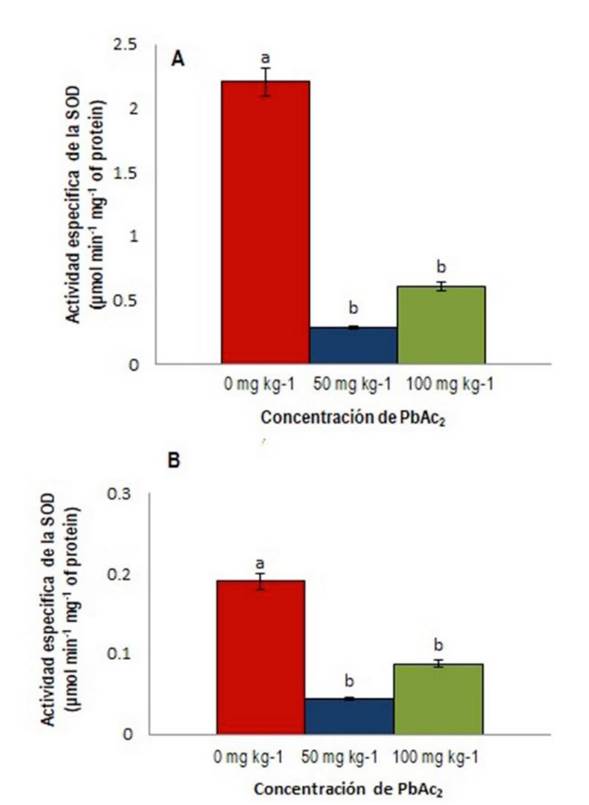
Figura 3 Actividad específica de la enzima SOD (µmol min-1 mg-1 de proteína); A) SOD en hojas, B) SOD en tallos, de S. lycopersicum cv. Mariela. Letras diferentes indican diferencias significativas (p ≤ 0.05).
Se analizaron los genes IFR, TCTP y SOD para comprender mejor la respuesta de defensa de la planta contra el estrés oxidativo (Figura 4). La ubiquitina se utilizó como gen de control debido a su expresión basal estable (Jain et al. 2006). Para todos los genes analizados, la expresión fue mayor bajo estrés por Pb con diferencias significativas al compararlas con el control. La expresión de la TCTP en hojas en comparación con el control fue 600% mayor a 50 mg kg-1 y 675% mayor a 100 mg kg-1 sin diferencias significativas entre los dos niveles de PbAc2; en los tallos la expresión fue 300% mayor a 50 mg kg-1 y aumentó 800% a 100 mg kg-1, en este caso con diferencias significativas entre todos los tratamientos. En plantas, según Bommer (2017), la TCTP parece tener una función protectora frente a estreses fitotóxicos, como la presencia de metales pesados, lo que puede explicar la alta expresión de este gen bajo estrés por Pb en ambas partes de la planta de tomate cv. Mariela. También se ha informado que algunos metales pesados en plantas regulan de forma positiva la TCTP bajo diversas condiciones de estrés celular (Schmidt et al. 2007). En los tallos esta expresión génica no fue diferente entre los tratamientos.
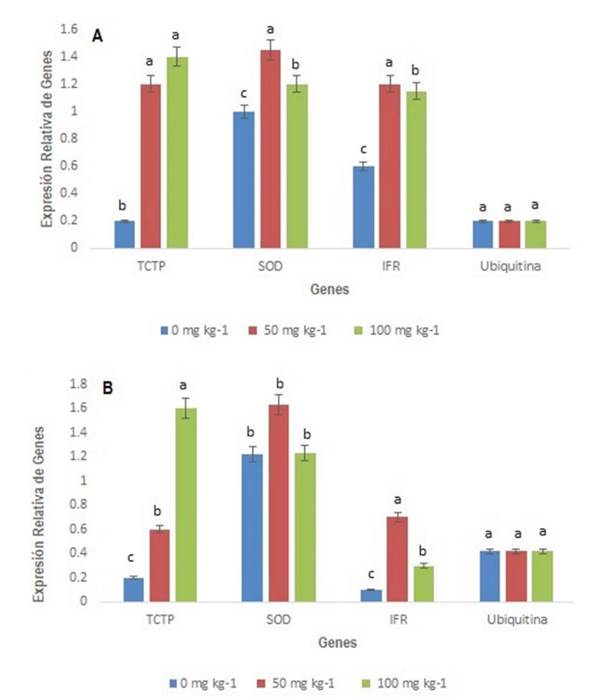
Figura 4 Expresión relativa de la Proteína Tumoral Controlada Transcripcionalmente (TCTP), la Superóxido Dismutasa (SOD) y la Isoflavona Reductasa (IFR) en S. lycopersicum cv. Mariela, crecida con diferentes concentraciones de Pb (PbAc2) (0, 50, 10 mg kg-1), A) Hojas, B) Tallos. Letras diferentes indican diferencias significativas (p ≤ 0.05).
La expresión del gen de la SOD en hojas y tallos fue superior a 50 mg kg-1 de PbAc2 lo cual coincide con lo obtenido en plantas de tomate cv Micro-Tom donde la expresión de la SOD fue mayor a la menor concentración de PbAc2 (Pérez et al. 2013), y difiere del estudio realizado en plantas de tomate cv. Amalia donde la expresión de la SOD aumentó con las dos concentraciones de PbAc2 utilizadas (Machín-Suárez et al. 2017). Los resultados sugieren que el perfil de la transcripción genética no está relacionado con la actividad enzimática bajo estrés por Pb y que este incremento en la expresión del gen de la SOD pudiera contrarrestar el efecto a nivel bioquímico en cuanto a la disminución de la actividad de la enzima.
La expresión del gen IFR fue mayor a 50 mg kg-1 en hojas y tallos en 250% y 400% respectivamente. Para el tratamiento de 100 mg kg-1 en hojas aumentó en 240% y en los tallos 200% mayor que el testigo con diferencias significativas entre los tratamientos. La expresión del gen IFR fue mayor en hojas y tallos en las dos concentraciones trabajadas de PbAc2, siendo diferente este resultado al obtenido en dos cultivares de arroz (Oryza sativa L.) Hennong 315 y Shendao 47 donde la expresión del gen de la IFR fue baja en ambos cultivares, lo que significa que el estrés por metales pesados disminuyó la expresión del gen (Wang et al. 2020). Resultados similares a los obtenidos en esta investigación fueron informados por Pérez et al. (2013) en plantas de tomate cv-Micro-Tom. La mayor expresión del gen IFR tal vez se deba a que los flavonoides pueden funcionar como quelantes de metales (Korkina 2007) y también protegen a las células contra el estrés oxidativo (Rusak et al. 2005). Debido a que hay poca información sobre la expresión del gen IFR en plantas, este trabajo es uno de los pocos que relaciona a este gen con el estrés inducido por Pb.
En plantas de tomate cv. Mariela, la toxicidad del Pb induce la actividad de la enzima GR, una de las enzimas importantes del sistema de defensa antioxidante y reduce la actividad de la SOD en ambas concentraciones de PbAc2 ensayadas. El análisis molecular indica mayor expresión de los tres genes estudiados (SOD, TCTP, IFR) bajo estrés por Pb, lo que sugiere que estos resultados pueden aplicarse en programas de mejoramiento genético de plantas de tomate, para obtener plantas con mayor tolerancia al Pb y/o con mejores capacidades de acumulación de este metal pesado.

