











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los micronutrientes son esenciales para el crecimiento y desarrollo normal de las plantas a pesar de la baja concentración en comparación con los macronutrientes. El rango dinámico de concentraciones internas óptimas de micronutrientes en las plantas requiere una regulación estricta para que sus requisitos se cumplan en diferentes condiciones ambientales (Castro et al. 2018). El boro (B) es un nutriente esencial para las plantas debido a que participa en la formación, estabilidad y función de la pared y la membrana celular, transporte de electrones de la membrana plasmática, metabolismo de los carbohidratos, metabolismo de fenol y auxinas, elongación y división celular, alargamiento de las raíces, fijación de nitrógeno y la asimilación de nitratos (Marschner 1995, Arunkumar et al. 2018, Brdar-Jokanović 2020, García et al. 2020). El B es absorbido por las plantas en forma de H3BO3 y H2BO3 (Arunkumar et al. 2018). Sin embargo, el B tiene un rango estrecho entre deficiencia y toxicidad (Brdar-Jokanović 2020, García et al. 2020). La toxicidad del B en las plantas está relacionada a la calidad del agua de riego utilizada y a los suelos con concentraciones excepcionalmente altas en B (Chatzissavvidis y Antonopoulou 2020). La toxicidad de B en el medio de crecimiento es una restricción para la productividad de los cultivos (El-Shazoly et al. 2019) , debido a que causa deterioro de las funciones celulares y de los procesos fisiológicos y bioquímicos de las plantas (Chatzissavvidis y Antonopoulou 2020).
El estrés abiótico es uno de los principales factores limitantes que influyen en el crecimiento y la productividad de las especies vegetales (Atif et al. 2019). El calcio (Ca) es un elemento esencial necesario para el crecimiento y desarrollo de las plantas en condiciones de estrés y sin estrés, ya que éste no solo cumple la una función importante en la estabilidad de la pared y la membrana celular, sino que también actúa como un segundo mensajero involucrado en varios procesos celulares (White y Broadley 2003, Aldon et al. 2018, Thor 2019). La variación intracelular de la concentración de Ca libre se encuentra entre los primeros eventos que siguen a la percepción de la planta del cambio ambiental (Aldon et al. 2018). El estrés, generalmente, conlleva a un incremento de Ca libre en el citoplasma de las células, el cual genera una expresión genética que activa respuestas bioquímicas que da a la planta la posibilidad de adaptarse a condiciones adversas de diferente naturaleza (Aldon et al. 2018). En cuanto la interacción entre el B y Ca, la información disponible es limitada y la naturaleza de esta compleja interacción aún se desconoce claramente. Por lo tanto, el objetivo del presente trabajo fue determinar si la aplicación de diferentes concentraciones de Ca puede aliviar la toxicidad de B en plantas de pimiento inducido por altas concentraciones de B presentes en la solución nutritiva.
Materiales y métodos
Localización
La presente investigación se llevó a cabo de julio a diciembre del 2019 en un invernadero del Departamento de Horticultura de la Universidad Autónoma Agraria Antonio Narro. Durante el experimento se presentó una temperatura promedio diurna de 30 °C, humedad relativa promedio de 55% y radiación fotosintéticamente activa incidente de 450 μmol m-2 S-1.
Material vegetal
Se utilizaron semillas pimiento amarillo cv. shir F1, las cuales se sembraron el 10 de julio en charolas de poliestireno de 200 cavidades, utilizando como sustrato peat moss. El trasplante se realizó a los 40 días después de la siembra en contenedores de polietileno negro con un volumen de 10 L. El sustrato utilizado para el trasplante fue compuesto de una mezcla de peat moss y perlita (70/30%, v/v).
Tratamiento
Los tratamientos consistieron en cinco concentraciones de B (0.5, 1.5, 3.0, 4.5 y 6.0 ppm) y tres concentraciones de Ca (7, 9 y 11 meq L-1), la combinación de estos dio un total de 15 tratamientos. Como fuente de boro y calcio se utilizaron ácido bórico y nitrato de calcio. Las diferentes concentraciones de Ca se obtuvieron a partir de las modificaciones de la solución Steiner (1961), variando únicamente la cantidad de Ca y el resto de los macronutrientes se mantuvieron constantes. Además, en cada tratamiento se agregaron los siguientes micronutrimentos (mg L-1): 5 Fe, 0.11 Zn, 0.65 Mn, 0.02 Cu y 0.05 Mo. Para la formulación de las soluciones nutritivas se consideró las propiedades químicas del agua de riego. El pH de las soluciones nutritivas se ajustó a 6.0 ± 0.1 con H3PO4 al 85% y HNO3 al 55%. Tres días después del trasplante se iniciaron los riegos con las soluciones correspondientes a cada tratamiento. Los riegos se realizaron en forma manual y la frecuencia dependió del requerimiento hídrico de las plantas, aplicando el volumen suficiente de la solución nutritiva para mantener una fracción de lixiviación del 20%.
Variables agronómicas evaluadas
A los 108 días después del trasplante, se evaluó la altura de planta midiendo desde la base hasta la parte apical. Los diferentes órganos de la planta se separaron, para luego lavar el sistema radicular con agua de la llave y agua destilada para eliminar el exceso de sustrato, posteriormente, se determinó la longitud radicular. La raíz, tallo y hoja se colocó de forma separada en bolsas de papel estraza y posteriormente se introdujeron en un horno de secado a temperatura de 65 °C durante 72 h, para después registrar la biomasa seca de cada órgano en una balanza analítica. El peso seco total se obtuvo de la sumatoria del peso seco de cada órgano evaluado.
Fotosíntesis
Para determinar la fotosíntesis se utilizó un equipo portátil (LI-6800XT, Lincoln, Nebraska). Las lecturas se efectuaron al inicio de la floración. La evaluación se realizó en hojas bien desarrolladas y completamente sanas, evaluando este parámetro entre las 12:00 y 13:00 h tomando una lectura por hoja en dos hojas por planta.
Concentración y contenido nutrimental
Los tejidos secos se digestaron en una mezcla de 2:1 de H2SO4: HClO4 y 2 mL de H2O2 al 30%. La determinación de N se realizó por el procedimiento de Micro-Kjeldahl (Bremmer 1996); mientras que, las concentraciones de fósforo (P), potasio (K), Ca, magnesio (Mg) y B se determinaron con espectrómetro de emisión de plasma acoplado inductivamente (ICP-AES, model Liberty, VARIAN, Santa Clara, CA) (Soltanpour et al. 1996). Una vez obtenido la concentración nutrimental, se utilizó la biomasa seca de los distintos órganos para obtener el contenido nutrimental por planta.
Análisis estadístico
El diseño experimental utilizado fue el de bloques completos al azar con un arreglo factorial de 5x3, con cuatro repeticiones en cada tratamiento. Los datos obtenidos se sometieron a un análisis de varianza (ANOVA) y la comparación de medias fue de acuerdo con la prueba de Tukey (α ≤ 0.05) utilizando el programa estadístico SAS (Statistical Analysis Systems) versión 9.0.
Resultados
Los parámetros de fotosíntesis, altura de planta, longitud de raíz, peso seco de raíz, peso seco de tallo, peso seco de hoja y peso seco total fueron afectados por la concentración de B en la solución nutritiva; mientras que, a excepción del peso seco de tallo, estos mismos parámetros fueron afectados por las concentraciones de Ca así como por la interacción de ambos factores evaluados (Tabla 1). Debido a las bajas temperaturas presentadas el experimento no pudo llegar a la etapa de producción.
Tabla 1 Efecto de la concentración de B y Ca en la solución nutritiva sobre la fotosíntesis, altura de planta, longitud de raíz, peso seco de raíz (PSR), peso seco de tallo (PST), peso seco de hoja (PSH) y peso seco total (PST) de plantas de pimiento amarillo cv. shir F1.
| B | Fotosíntesis | Altura de planta | Longitud de raíz | PSR | PST | PSH | PST |
|---|---|---|---|---|---|---|---|
| (ppm) | (μmol m-2 s-1) | (cm) | (g planta-1) | ||||
| 0.5 | 3.77b | 59.28a | 43.33a | 5.33b | 13.05b | 21.39c | 39.78b |
| 1.5 | 3.98ab | 57.22a | 41.50abc | 7.00a | 15.33a | 22.50b | 44.83a |
| 3 | 4.05a | 56.89a | 42.00ab | 6.11ab | 14.00ab | 24.11a | 44.00a |
| 4.5 | 3.98ab | 53. 44b | 41.06bc | 5.67b | 13.22b | 24.17a | 43.05a |
| 6 | 3.49c | 50.56c | 39.67c | 5.44b | 12.67b | 24.83a | 42.94b |
| Anova p ≤ Ca (meqL-1) | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| 7 | 3.84ab | 55.27b | 43.47a | 5.47b | 13.20a | 22.16b | 40.70b |
| 9 | 3.97a | 57.43a | 40.37b | 6.27a | 13.77a | 24.27a | 44.30a |
| 11 | 3.76b | 53.73b | 40.70b | 6.00ab | 14.00a | 23.76a | 43.76a |
| Anova p ≤ | 0.004 | 0.001 | 0.001 | 0.036 | 0.118 | 0.001 | 0.001 |
| Interacción | 0.001 | 0.006 | 0.001 | 0.026 | 0.111 | 0.001 | 0.001 |
| CV % | 4.03 | 3.21 | 3.72 | 13.80 | 7.68 | 3.07 | 4.54 |
La fotosíntesis incrementó al aumentar la concentración de B, con un máximo con 4.5 ppm de B, presentando este comportamiento cuando se aumentó la concentración de Ca en la solución nutritiva a 9 y 11 meq L-1; mientras que al adicionar 7 meq L-1 de Ca la fotosíntesis decreció cuando la concentración de B fue mayor a 1.5 ppm (Figura 1A ). La altura de planta se redujo conforme incrementó la concentración de B en la solución nutritiva, pero este detrimento fue memos pronunciado al incrementar la concentración de Ca en la solución nutritiva de 9 meq L-1 (Figura 1B ). La longitud de raíz fue superior con 3 ppm de B con 7 meq L-1 de Ca en la solución nutritiva; mientras que al adicionar 9 y 11 meq L-1 de Ca la longitud de raíz decreció con más de 0.5 ppm de B (Figura 1C).
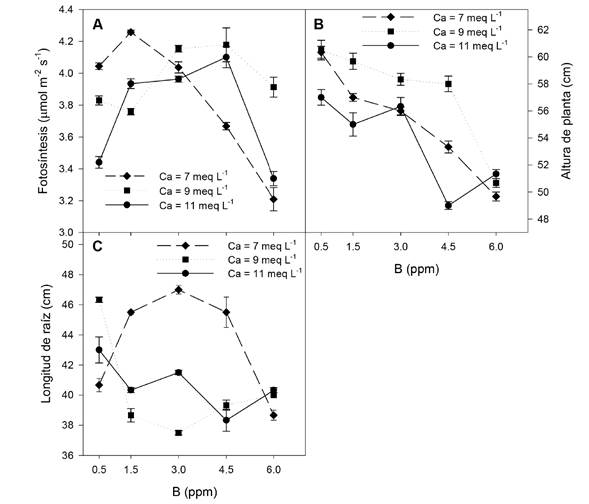
Figura 1 Efecto de las concentraciones de B y Ca en la solución nutritiva sobre la fotosíntesis (A), altura de planta y longitud de raíz de plantas de pimiento amarillo cv. shir F1. Las barras indican el error estándar de la media.
El peso seco de raíz fue mayor con 1.5 ppm de B en la solución, mientras que valores inferiores o superiores a esta concentración el peso seco de raíz decreció, este mismo comportamiento se observó cuando la concentración de Ca en la solución nutritiva fue de 7 y 9 meq L-1. En tanto que al incrementar la concentración de Ca a 11 meq L-1 el peso seco de raíz fue mayor con 3 ppm de B (Figura 2A). El peso seco de hoja, en general, incrementó al aumentar la concentración de B, principalmente cuando se adicionó 7 y 9 meq L-1 de Ca en la solución nutritiva; pero, al incrementar la concentración de Ca a 11 meq L-1 el peso seco de hoja fue mayor al adicionar una concentración de 1.5 y 4.5 ppm de B (Figura 2B). Similar comportamiento se observó en el peso seco total (Figura 2C).
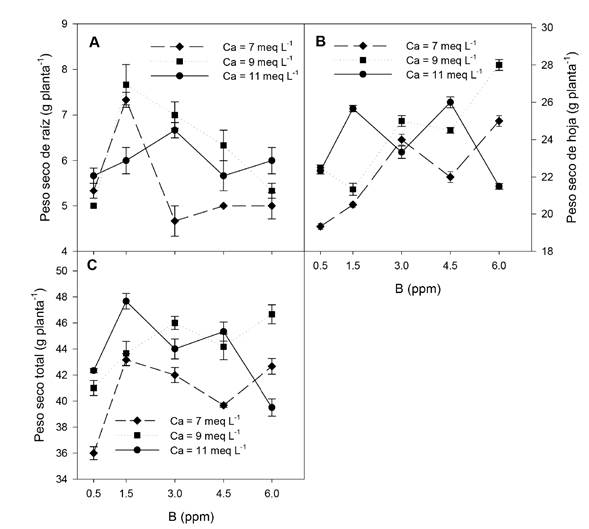
Figura 2 Efecto de las concentraciones de B y Ca en la solución nutritiva sobre el peso seco de raíz y hoja de plantas de pimiento amarillo cv. shir F1. Las barras indican el error estándar de la media.
El contenido de N, P K, Ca, Mg y B en las plantas de pimiento fue afectado de forma significativa por las concentraciones de B en la solución nutritiva; con excepción del contenido total de Mg, el contenido total de estos iones en las plantas de pimiento fue afectado por las concentraciones de Ca en la solución nutritiva. También el contenido total de N, P, K, Ca, Mg y B fue influenciado de forma significativa por la interacción de los dos factores evaluados (Tabla 2).
Tabla 2 Efecto de las concentraciones de B y Ca en la solución nutritiva sobre el contenido total de nitrógeno (N), fosforo (P), potasio (K), calcio (Ca), magnesio (Mg) y boro (B) en las plantas de pimiento amarillo cv. shir F1.
| B | N | P | K | Ca | Mg | B |
|---|---|---|---|---|---|---|
| (ppm) | (mg planta-1) | |||||
| 0.5 | 871.48c | 143.506b | 953.01b | 531.26b | 184.34e | 2.69e |
| 1.5 | 997.22a | 170.407a | 1093.19a | 637.67a | 232.77c | 4.88a |
| 3 | 986.78ab | 166.016a | 1000.12b | 618.48a | 260.41a | 4.19c |
| 4.5 | 944.14b | 165.251a | 1062.16a | 615.24a | 243.80b | 3.57d |
| 6 | 958.52ab | 163.045a | 1055.37a | 543.04b | 208.95d | 4.49b |
| Anova p ≤ | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| Ca (Meq L-1) | ||||||
| 7 | 925.62b | 153.160c | 1011.79b | 576.60b | 223.864a | 4.10a |
| 9 | 970.70a | 168.575a | 1069.40a | 599.66a | 225.104a | 4.09a |
| 11 | 940.56ab | 163.201b | 1017.12b | 591.15ab | 229.216a | 3.70b |
| Anova p ≤ | 0.010 | 0.001 | 0.001 | 0.022 | 0.087 | 0.001 |
| Interacción | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| CV (%) | 4.03 | 3.42 | 3.52 | 3.66 | 2.94 | 3.42 |
Medias con letra diferentes indican efectos significativos según la prueba de comparación múltiple de Tukey con p ≤ 0.05. Anova = análisis de varianza. Interacción = B x Ca. CV = coeficiente de variación.
El mayor contenido de N en las plantas de pimiento se presentó en aquellas plantas que se les proporcionó 1.5 ppm de B y 7 meq L-1 de Ca, con una concentración diferente de B el contenido total de N decreció; pero cuando se incrementó la concentración de Ca en la solución a 9 y 11 meq L-1 el mayor contenido de N se obtuvo con 3 ppm de B, mientras que con una concentración diferente de B el contenido de N fue menor, aunque el detrimento fue menos pronunciado que con 7 meq L-1 de Ca (Figura 3A). De igual manera, el contenido total de P y K en las plantas de pimiento fue mayor con 1.5 ppm de B al adicionar 7 y 9 meq L-1 de Ca, pero con una concentración inferior o superior de B el contenido de estos iones decreció, con un detrimento menos marcado con 9 meq L-1 de Ca (Figura 3B y C). El contenido de Ca total en las plantas de pimiento fue mayor con 1.5 y 3 ppm de B y 7 meq L-1 de Ca; mientras que, al incrementar la concentración de Ca a 9 meq L-1 el contenido total de Ca fue mayor con 1.5 ppm de B, superando al contenido total alcanzado con 7 meq L-1 de Ca; en tanto que, al incrementar la concentración de Ca a 11 meq L-1 el contenido total de Ca fue superior con 4.5 ppm de B (Figura 3D).
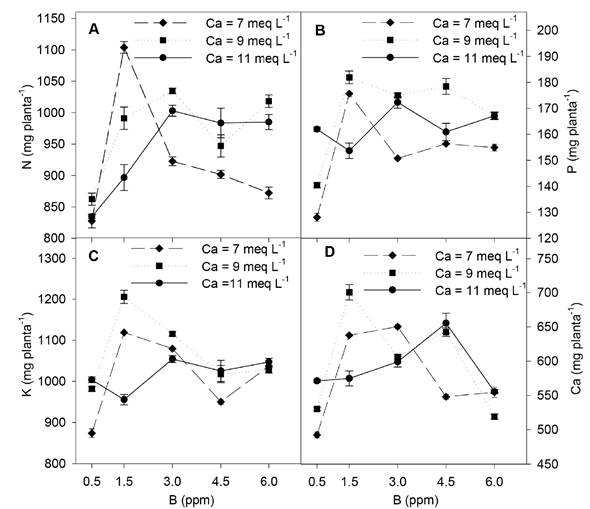
Figura 3 Efecto de las concentraciones de B y Ca en la solución nutritiva sobre el contenido total de nitrógeno (N), fósforo (P), potasio (K) y calcio (Ca) en plantas de pimiento amarillo cv. shir F1. Las barras indican el error estándar de la media.
El contenido total de Mg en las plantas de pimiento incrementó al aumentar la concentración de B en la solución nutritiva, observando este comportamiento con las diferentes concentraciones de Ca adicionados a la solución nutritiva. Pero con concentraciones de 7 y 11 meq L-1 de Ca el contenido máximo de Mg se observó con 3 ppm de B, pero al adicionar 9 meq L-1 de Ca el mayor contenido de Mg se presentó con 4.5 ppm de B (Figura 4A). Por otro lado, el contenido total de B en las plantas fue mayor con 1.5 y 3 ppm de B al adicionar únicamente 7 meq L-1 de Ca en la solución nutritiva; pero, al incrementar la concentración de Ca a 9 meq L-1 el mayor contenido total de B se presentó con 1.5 ppm de B, con contenidos diferentes el contenido total de B decreció; mientras que, al incrementar la concentración de Ca a 11 meq L-1 el contenido de B fue mayor con 1.5 y 6 ppm de B (Figura 4B).

Discusión
El B es un micronutriente esencial para el crecimiento y desarrollo óptimo de las plantas debido a que ejerce funciones importantes a nivel fisiológico y metabólico (Marschner 1995, Arunkumar et al. 2018). Pero cuando la concentración de B excede el valor óptimo determinado puede resultar tóxico para las plantas (Chatzissavvidis y Antonopoulou 2020). Los requerimientos de B pueden variar entre especies y/o cultivares, algunas especies pueden tolerar hasta 0.5 mg L-1, mientras que otras pueden llegar a toleran hasta 4 mg L-1 (Batar et al. 2009, Princi et al. 2015, García et al. 2020). En este trabajo se observó que el B en altas concentraciones (mayor a 0.5 ppm) en la solución nutritiva afecta negativamente el crecimiento de las plantas de pimiento, ya que, en general, se observó un detrimento de la altura de planta y longitud de raíz al emplear concentraciones altas de este ion. Relacionado a lo anterior, Yermiyahu et al. (2008) indicaron que el mayor crecimiento de las plantas de pimiento cv. Maor se obtuvo con 0.093 mM de B; mientras que, el mayor crecimiento de pimiento cv. Celica se obtuvo con 0.05 mM. Sin embargo, la toxicidad sobre la altura de planta se redujo al incrementar la concentración de Ca en la solución nutritiva a 9 meq L-1. El Ca al ser un mensajero secundario ante condiciones de estrés (Gao et al. 2019, Thor 2019), pudo influir en la mitigación de la toxicidad de B en las plantas de pimiento. Siddiqui et al. (2013) indicaron que al incrementar la concentración de Ca se alivió la toxicidad del B en plantas de rábano, debido a que el incremento del Ca disminuyó los niveles de malondialdehído y peróxido de hidrógeno y la fuga de electrolitos y mejoró las actividades de las enzimas antioxidantes. En tanto que, en la longitud de la raíz no se observó el efecto positivo del Ca al incrementar su concentración en la solución nutritiva. Esta respuesta alcanzada pudo deberse a que la concentración de B es inferior en las raíces (García et al. 2020), ya que el B se transporta rápidamente a los órganos superiores y se acumula allí (Camacho et al. 2008).
Se ha indicado que las alta concentración de B afecta negativamente el proceso fotosintético, ya que este ion daña el contenido de clorofila, la velocidad de transporte de electrones, la eficiencia del uso de CO2, el fotosistema II, la subunidad de H+-ATPasa y algunas actividades relacionadas con las enzimas Rubisco y la NADP-GAPDH (Han et al. 2009, Chen et al. 2014, Mesquita et al. 2016, Landi et al. 2019, Riaz et al. 2019). La reducción de la fotosíntesis debido a las altas concentraciones de B se vio en este trabajo; sin embargo, cuando se incrementó la concentración de Ca en la solución nutritiva a 9 y 11 meq L-1 el efecto tóxico del B se observó hasta con 6 ppm en comparación a las plantas a las que se les adicionó 7 meq L-1 de Ca. Tomando en cuenta que el Ca ayuda a las plantas a protegerse contra la peroxidación lipídica, fuga de electrolitos y contenido de H2O2 producidos ante diferentes condiciones adversas del medio (Siddiqui et al. 2013), al incrementar la concentración de Ca en la solución nutritiva la planta de pimiento pudo incrementar la absorción de Ca para mitigar en mayor medida el estrés causado en el proceso fotosintético al adicionar una concentración alta de B, comparado al adicionar únicamente 7meq L-1 de Ca.
Por otro lado, Marschner (1995) menciona que la producción de biomasa de un cultivo se determina en gran medida por la fotosíntesis, ya que aproximadamente el 90% del peso seco de una planta se considera derivado de los productos formados durante este proceso. En este trabajo se observó que el efecto de las altas concentraciones de B en la solución nutritiva sobre la fotosíntesis también se vio reflejado en la producción de biomasa, donde la mayor actividad de esta variable fisiológica se relacionó con los tratamientos donde se obtuvo la mayor producción de biomasa seca de raíz, hoja y total, principalmente al adicionar 9 y 11 meq L-1 de Ca, resultado que pudo relacionarse a que, además de su participación como segundo mensajero, el Ca cumple funciones en la biogénesis de la pared celular, división, expansión y organización celular (Kolling et al. 2019), por lo que al aumentar la concentración de Ca se logró generar un mayor desarrollo vegetal lo que se reflejó en una mayor biomasa seca. Relacionado a lo anterior, Turan et al. (2009) y Metwally et al. (2018) reportaron que la aplicación de altos niveles de Ca alivió parcialmente los síntomas de toxicidad de B sobre la producción de biomasa seca de brotes y raíz de plantas de trigo y canola. Además, de acuerdo al comportamiento de la longitud radicular y peso seco de raíz, se observó que al adicionar 9 y 11 meq L-1 de Ca y concentraciones superiores a 0.5 ppm de B en la solución nutritiva las plantas de pimiento modificaron la arquitectura radicular produciendo un sistema de raíces más ramificadas al reducir el crecimiento de la raíz primaria, ya que a pesar de tener una longitud radicular inferior se obtuvo mayor peso seco radicular con estos tratamientos.
Una dosis inadecuada de B afecta el estado nutrimental de la planta (Sarafi et al. 2017). Sarafi et al. (2018b) indican que para obtener un crecimiento óptimo de las plantas y una absorción máxima de los nutrimentos, la concentración de B en la solución nutritiva en el cultivo de pimiento no debe exceder 1 mg L-1. Por su parte, Princi et al. (2015) y Ferreira et al. (2020) indican que el exceso de B puede afectar la absorción N por las raíces, esto debido a que hay una alteración de su metabolismo. En este trabajo se observó una reducción del contenido total de N en las plantas de pimiento al incrementar la concentración de B en la solución nutritiva, principalmente cuando se adicionó únicamente 7 meq L-1 de Ca; sin embargo, al incrementar la concentración de Ca (9 y 11 meq L-1) en la solución nutritiva el contenido total de N en las plantas presentó un menor detrimento a altas concentraciones de B, demostrando que al aumentar la concentración de Ca se mantiene el proceso metabólico de la pared celular y el transporte de iones, tal como es mencionado en otros estudios (Tariq y Mott 2007, Sarafi et al. 2018a). En este experimento se observó que el contenido total de P en las plantas de pimiento fue afectado por las concentraciones empleadas de B, lo que concuerda con Sarafi et al. (2018b), quienes informaron una reducción de la absorción total de P en plantas de pimiento al aumentar los niveles de B en la solución nutritiva; pero, de igual manera que el N, el contenido total de P en las plantas fue menos afectado por el exceso de B al incrementar la concentración de Ca en la solución nutritiva (9 y 11 meq L-1).
Asimismo, el contenido total de K en las plantas de pimiento también presentó una reducción al aumentar la concentración de B en la solución nutritiva. Sarafi et al. (2018b) reportaron una reducción en la absorción de K al aumentar la concentración de B en la solución nutritiva, indicando que esta respuesta pudo deberse posiblemente a la reducción del crecimiento de la planta o de las raíces. Sin embargo, el efecto benéfico del incremento de la concentración de Ca en la solución sobre el efecto inhibidor del exceso de B en el contenido nutrimental únicamente se observó con 9 meq L-1, en tanto que, al incrementar el Ca a 11 meq L-1 se observó un efecto negativo en el contenido total de K en las plantas de pimiento. La respuesta anterior puede atribuirse a que el K presenta antagonismo con Ca (Ranade-Malvi 2011).
De la misma forma, se observó un detrimento del contenido total de Ca en las plantas de pimiento al aumentar la concentración de B en la solución nutritiva, principalmente con 7 y 9 meq L-1 en la solución nutritiva; mientras que, al aumentar la concentración de Ca en la solución a 11 meq L-1 el contenido total de Ca incrementó, con un máximo con 4.5 ppm de B; sin embargo, cuando se empleó una concentración de 9 meq L-1 de Ca y 4.5 ppm de B en la solución nutritiva se observó una recuperación del contenido total de Ca, obteniendo semejante contenido total de Ca que el mayor contenido obtenido con 11 meq L-1 de Ca. El Ca es un nutriente mineral esencial en las plantas, el cual realiza la función de mensajero secundario ante condiciones de estrés (White y Broadley 2003, Rahman et al. 2016). La señalización del Ca a condiciones desfavorables se lleva a cabo mediante una acumulación de éste en el citosol, para posteriormente conllevar a una reacción en cadena obteniéndose como resultado la activación de factores de trascripción para la expresión de genes de defensa (Thor 2019). Por lo que, la respuesta del contenido total de Ca en este trabajo ante concentraciones altas de B pudo deberse a que la planta al estar bajo estrés por las concentraciones altas de B tuvo mayor absorción y acumulación de Ca; sin embargo, esta respuesta dependió de la disponibilidad de Ca en el medio de crecimiento, por lo que el mayor contenido total de Ca con concentraciones altas de B se presentó con concentraciones de 9 y 11 meq L-1 de Ca en la solución nutritiva. El efecto adverso de la toxicidad de B en el contenido total de Mg en las plantas de pimiento se observó en este trabajo; sin embargo, no se obtuvo una respuesta positiva del incremento de la concentración de Ca en la solución nutritiva.
A pesar de que en este trabajo se incrementó la concentración de B en la solución nutritiva, se observó que cuando la concentración de B fue mayor a 1.5 ppm el contenido total de B en las plantas de pimiento se redujo, alcanzando un mínimo con 4.5 ppm de B, con detrimento al incrementar la concentración de Ca en la solución (9 y 11 meq L-1); pero, cuando la concentración de B se incrementó a 6 ppm se observó una recuperación del contenido total de B en las plantas. El menor contenido total de B en las plantas de pimiento alcanzado con una concentración de 4.5 ppm de B y 9 y 11 meq L-1 de Ca concuerda con el incremento del contenido total de Ca en las plantas con estos tratamientos, por lo tanto, el incremento del contenido total de Ca reguló la absorción de B cuando éste se encontraba en altas concentraciones en el medio de crecimiento. Relacionado a lo anterior, se ha indicado que la aplicación de Ca puede reducir la absorción de B en plantas de rábano, canola y trigo, debido a que disminuye la permeabilidad de la membrana de célula vegetal a este ion (Turan et al. 2009, Siddiqui et al. 2013, Metwally et al. 2018).
Conclusiones
Concentraciones superiores a 1.5 ppm de B afectan de forma negativa la fotosíntesis, crecimiento, producción de biomasa y absorción de nutrientes en las pantas de pimiento morrón. Pero al incrementar la concentración de Ca en la solución nutritiva a 9 y 11 meq L-1, las plantas de pimiento pueden tolerar concentraciones mayores de B. Además, cuando la concentración de Ca en la solución nutritiva es mayor a 7 meq L-1 el contenido total de B en el tejido vegetal se reduce; por lo tanto, el Ca regula la absorción de B cuando este se encuentra en altas concentraciones en el medio de crecimiento.

