











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La fragmentación antropogénica del hábitat altera la geometría espacial, la conectividad y el grado de aislamiento de los fragmentos remanentes. El cambio de uso de suelo asociado a estas modificaciones afecta los procesos ecológicos a diferentes niveles, desde el comportamiento de los individuos y la dinámica de las poblaciones, hasta la composición y estructura de las comunidades y los flujos de materia y energía (Trilleras et al. 2015). Los bosques tropicales albergan el 70% de la biodiversidad mundial. Pero se estima que en la región tropical se pierde aproximadamente 15 millones de hectáreas de bosque maduro por año, de las cuales el 60% corresponden a la fragmentación derivada de prácticas agrícolas, pecuarias y forestales, urbanización, minería, tala furtiva e incendios (Cristóbal et al. 2014).
Entender cómo se modifican los patrones ecológicos en hábitats fragmentados es clave para comprender el impacto que tiene la perturbación antropogénica sobre la dinámica de los ecosistemas y escalas (Ries et al. 2004). Por ello es fundamental evaluar los cambios en la estructura de las comunidades y, en consecuencia, en la red de interacciones entre las diferentes especies que las componen (Suzán et al. 2008). El uso de bioindicadores es una herramienta útil para conocer el grado de integridad de un ecosistema mediante el monitoreo de especies de vertebrados e invertebrados con altas abundancias, y que además sean ecológica, taxonómica y tróficamente diversas. Las especies bioindicadoras tienen un papel importante dentro del ecosistema y responden de forma cuantitativa y predecible a cambios en el entorno, por lo que el análisis de métricas ecológicas como abundancia y diversidad de especies puede ayudar a detectar cambios ambientales (Russo y Jones 2015).
Los murciélagos son el segundo orden de mamíferos con mayor diversidad de especies en el mundo, comprendiendo más de 1 300 especies (Voigt y Kingston 2016). En México este orden está conformado por 140 especies, de las cuales 16 son endémicas (Ceballos y Arroyo-Cabrales 2012, Ramírez-Pulido et al. 2014). La elevada dependencia que presentan algunas especies a hábitats específicos, el grado de especialización en su alimentación y el uso de refugios, las hace susceptibles a las modificaciones del paisaje, por lo que han sido propuestas como bioindicadores del estado de conservación de bosques húmedos neotropicales (Medellín et al. 2000, Russo y Jones 2015). La importancia de este grupo de mamíferos se debe, entre otros factores, a la diversidad de procesos ecológicos en los que interviene, como la polinización de plantas y dispersión de semillas que son importantes para la regeneración de selvas y bosques tropicales, además de desempeñarse como control biológico de poblaciones de insectos (Kunz et al. 2011).
El interés por el estudio del papel de los murciélagos como hospederos de una gran diversidad de endoparásitos, ectoparásitos y diversos agentes patógenos ha aumentado en los últimos años. Entre los ectoparásitos asociados a los murciélagos en América se han registrado principalmente a las familias Nycteribiidae, Streblidae, Spinturnicidae, Spelaeorhynchidae, Ixodidae, Argasidae e Ischnopsyllidae (Colín-Martínez y García-Estrada 2018, Colín-Martínez et al. 2018, Cuxim-Koyoc et al. 2018, Czenze y Broders 2011, González-Acuña et al. 2018). Los ácaros pertenecientes a las familias Spinturnicidae y Spelaeorhynchidae, específicos de murciélagos, presentan adaptaciones que responden a una forma de vida ectoparasitaria que incluyen hábitos hematófagos. Los dípteros de las familias Nycteribiidae y Streblidae son los ectoparásitos más abundantes y mejor adaptados de los murciélagos (Dittmar et al. 2006).
En distintos estudios, el hospedero suele considerarse como hábitat para el parásito, ya que el cuerpo del hospedero proporciona una amplia gama de estímulos ambientales y recursos y juega un papel importante en la configuración de la historia de vida, la ecología y el comportamiento de los parásitos (Presley 2010). Sin embargo, el nivel de infestación de un hospedero por un ectoparásito particular depende no sólo de las características del hospedero, sino también del hábitat y del tipo de refugio del murciélago (Patterson et al. 2007, Pilosof et al. 2012). Las condiciones ambientales afectan la dinámica de las poblaciones de parásitos, así como los ensambles e interacciones a nivel de comunidad entre parásitos y hospederos. Distintas especies de parásitos que son sensibles a las modificaciones del hábitat del hospedero han sido reconocidas como posibles bioindicadores (Mackenzie y Hemmingsen 2014, Patterson et al. 2007, Patterson et al. 2008).
En trabajos recientes se ha estudiado el efecto de la perturbación antropogénica sobre la prevalencia de ectoparásitos de murciélagos orientados a las interacciones entre el ambiente, el hospedero y los parásitos. La modificación de estas interacciones puede desencadenar procesos de selección en los ectoparásitos, modificando la especificidad filogenética hacia sus hospederos por ausencia o presencia de estos (Dittmar et al. 2015). A su vez incrementa la probabilidad de encontrar hospederos accidentales en sitios con distintos grados de perturbación (Poulin et al. 2011). Por ello, resulta necesario considerar las condiciones ambientales, las variables climáticas, la presencia de actividades antropogénicas y la fragmentación de los hábitats en el estudio de las comunidades de ectoparásitos (Bolivar-Cimé et al. 2018, Hernández-Martínez et al. 2018). El objetivo del presente trabajo fue evaluar la composición y la filoespecificidad de comunidades de ectoparásitos asociados a murciélagos en tres tipos de paisajes agropecuarios en el estado de Veracruz.
MATERIALES Y MÉTODOS
Descripción del área de estudio
El trabajo se desarrolló en el centro y sur del estado de Veracruz. La ubicación geográfica del estado ofrece un entorno de características tropicales, que se modifican por la influencia de las cadenas montañosas con climas que varían de muy calientes a muy fríos. La precipitación anual oscila entre 500 mm en algunos lugares del norte y alrededor de 4 000 mm en el sur (Estrada-Contreras et al. 2015). Se distinguen dos épocas: 1) secas, de marzo a mayo con precipitación menor a 100 mm por mes, y 2) lluvias, de junio a febrero con precipitaciones superiores a 200 mm por mes.
Sitios de estudio
Los ocho sitios de muestreo se ubican en los municipios de Tlapacoyan, Martínez de la Torre, Úrsulo Galván, Puente Nacional y San Andrés Tuxtla. Cada sitio corresponde a una unidad de producción pecuaria con bovinos de doble propósito y la distancia mínima entre cada uno fue de al menos 2 km. Las unidades de producción, que en general se encuentran establecidas bajo un sistema ganadero de pastoreo controlado, representan un gradiente de paisajes con diferente nivel de transformación (Tabla 1). Para fines de este estudio, se consideraron tres categorías de paisaje: pastizal, silvopastoril y acahual. Se consideró como pastizal inducido a aquellos sitios cuyo paisaje estuviera dominado por la presencia de gramíneas forrajeras; silvopastoril a los sitios en los que una combinación de gramíneas forrajeras, leguminosas arbóreas y arbustivas dominara el paisaje; y acahual a los sitios con paisajes dominados por vegetación secundaria derivada de una selva alta perennifolia que incluyera la presencia de árboles, arbustos, herbáceas y gramíneas.
Tabla 1 Sitios de muestreo y coordenadas geográficas.
| Sitio | Paisaje | Municipio | Coordenadas |
|---|---|---|---|
| Clarín | Pastizal 1 | Tlapacoyan | 20o 02 03 Norte 97o 06 14 Oeste |
| Soledad | Silvopastoril 1 | Martínez de la Torre | 20o 02 08 Norte 97o 04 39 Oeste |
| Encinar | Acahual 1 | Tlapacoyan | 19o 54 43 Norte 97o 12 10 Oeste |
| CBTA 17 | Silvopastoril 2 | Úrsulo Galván | 19o 24 34 Norte 96o 22 33 Oeste |
| Utrera | Acahual 2 | Puente Nacional | 19o 17 06 Norte 96o 31 11 Oeste |
| Perico | Pastizal 2 | San Andrés Tuxtla | 18o 39 42 Norte 95o 12 55 Oeste |
| Divino | Pastizal 3 | San Andrés Tuxtla | 18o 38 32 Norte 95o 11 36 Oeste |
| Ruiz | Acahual 3 | San Andrés Tuxtla | 18o 40 13 Norte 95o 07 34 Oeste |
Captura de murciélagos
El trabajo de campo se realizó de septiembre a noviembre de 2016, meses que corresponden a la época de lluvias, y de marzo a mayo de 2017, en la época de secas. Para capturar los murciélagos se utilizaron ocho redes de niebla de 12 m de largo por 2.5 m de altura en cada sitio de muestreo por noche. Por cada sitio, se muestreó durante dos noches en lluvias y dos noches en secas, con un esfuerzo de muestreo de 7 h/red por noche para un total de 28 h/red por sitio de muestreo considerando ambas temporadas. La colocación de las redes fue pareada y la distancia mínima entre cada grupo de redes fue de 50 metros para evitar competencia entre ellas. Cada red se revisó con un periodo no mayor a 30 minutos entre una liberación y otra para evitar perturbar el área de captura y minimizar el tiempo en que los murciélagos quedaran enmallados en la red. Una vez capturado, cada individuo fue mantenido dentro de un saco de tela de manera individual para posteriormente obtener los valores morfométricos necesarios para su identificación taxonómica. La identificación hasta nivel de especie se realizó con la guía de campo de Medellín et al. (2008). La captura y manejo de los organismos se realizó cumpliendo las directrices de la Sociedad Americana de Mastozoología para el uso de mamíferos silvestres en investigación (Sikes y Bryan 2016).
Captura e identificación ectoparásitos
Los murciélagos capturados se sometieron a inspección visual sistematizada, en la que se capturaron todos los ectoparásitos que se detectaron a simple vista. Para localizar los ectoparásitos, la búsqueda se realizó de la cabeza a los miembros pélvicos con énfasis en regiones como orejas, propatagio, plagiopatagio y uropatagio. La captura de ectoparásitos se llevó a cabo con la ayuda de pinzas entomológicas en los murciélagos. Una vez obtenida la muestra se liberó a cada individuo. Las muestras se colocaron en tubos de 2 ml con aproximadamente 1.5 ml de alcohol al 70% para fijarlos y se mantuvieron a temperatura ambiente (Bezerra et al. 2016). En el laboratorio, los ácaros presentes en las muestras se separaron para someterlos a un proceso de aclarado con lactofenol a temperatura ambiente (de Almeida et al. 2017) por aproximadamente 45 días. Una vez aclarados se retiraron de los tubos y de manera directa se realizó el montaje en medio CMC-10. Todas las muestras montadas se colocaron en charolas de metal y se realizó la desecación del medio con ayuda de una incubadora a 50 °C durante una semana (Kazak et al. 2017). Para una mejor conservación de los montajes se realizó el sellado de los cubreobjetos con un anticorrosivo formulado a base de resinas alquidálicas y cromato de zinc. Para la identificación taxonómica de los ectoparásitos se utilizaron microscopios estereoscópico y óptico (Wenzel 1976).
Análisis de datos
Todos los análisis se realizaron con la ayuda del software libre R v. 3.4.3. La descripción de la diversidad de especies de hospederos y ectoparásitos se realizó con el índice de entropía de Shannon. Con los valores de diversidad de hospederos y ectoparásitos se calculó el coeficiente de correlación de Pearson. Se generaron curvas de rango-abundancia para describir la estructura de la comunidad de especies de ectoparásitos por sitio de muestreo y por especie hospedera. A la variable de la abundancia relativa se le realizó la prueba de Shapiro-Wilk para conocer la distribución de los datos. Se realizó un análisis de covarianza (ANCOVA) considerando en nuestro modelo como variable respuesta a la abundancia relativa de los ectoparásitos, como variable explicativa al tipo de paisaje y como covariable al rango de las especies ectoparásitas (Abella-Medrano et al. 2015). La diversidad beta de las comunidades de ectoparásitos y hospederos de los ocho sitios se cuantificó con el índice de Morisita-Horn. Se utilizó el coeficiente de correlación de Pearson para analizar la relación entre las abundancias relativas de todos los pares de especies ectoparásitas de las especies de murciélagos generalistas (López-Pérez et al. 2018).
Para determinar la especificidad filogenética de los ectoparásitos, es decir el grado de asociación parásito-hospedero explicado por la filogenia de los hospederos, se comparó el efecto de la diversidad filogenética, entendido como el agrupamiento filogenético (AF) de las especies de murciélagos utilizadas por una especie parásita determinada contra un modelo nulo basado en el efecto del tamaño estandarizado de la muestra total (Poulin et al. 2011). El árbol filogenético utilizado como base para realizar el cálculo de AF fue el súper-árbol filogenético de mamíferos publicado por Bininda-Emonds (2007).
RESULTADOS
Se registraron murciélagos de las familias Phyllostomidae y Mormoopidae, con un total de 23 especies, de las cuales las más abundantes fueron Artibeus jamaicensis, A. lituratus y Sturnira hondurensis. Dichas especies estuvieron presentes en todos los sitios de muestreo. Se capturaron 944 individuos, de los cuales 428 fueron portadores de al menos un ectoparásito; la prevalencia total de ectoparásitos fue de 45.34% (IC 42.16 - 48.51%). Se realizó la identificación taxonómica de 1 433 ectoparásitos, 995 dípteros de la familia Streblidae, 432 ácaros de la familia Spinturnicidae y seis ácaros de la familia Spelaeorhynchidae, para un total de 35 especies. Las especies más abundantes fueron Trichobius costalimai (Streblidae) y Periglischrus iheringi (Spinturnicidae). Para los estréblidos, se registraron especies de las tres subfamilias registradas para el Nuevo Mundo. La riqueza de ectoparásitos fue de 32 especies para la familia Phyllostomidae y cuatro para la familia Mormoopidae; entre ambas familias se compartió el registro de Periglischrus ojastii (Tabla 2). Las especies hospederas con mayor número de asociaciones de especies ectoparásitas fueron A. jamaicensis, A. lituratus y S. hondurensis.
Tabla 2 Número de individuos capturados de cada especie de ectoparásito por especie hospedera y por tipo de paisaje.
| Ectoparásito | ATE | Hospedero |
|---|---|---|
| P S A | ||
| Anastrebla mattadeni | 0 0 1 | Artibeus jamaicensis |
| 0 0 1 | Carollia perspicillata | |
| 0 0 1 | Glossophaga soricina | |
| Aspidoptera delatorrei | 0 6 0 | Sturnira hondurensis |
| 2 5 0 | Sturnira parvidens | |
| Aspidoptera falcata | 0 2 0 | Artibeus lituratus |
| 1 1 6 | Sturnira hondurensis | |
| 0 0 2 | Sturnira parvidens | |
| Aspidoptera phyllostomatis | 56 4 39 | Artibeus jamaicensis |
| 0 0 2 | Phyllostomus discolor | |
| 0 0 1 | Platyrrhinus helleri | |
| 1 2 1 | Sturnira hondurensis | |
| 0 1 1 | Sturnira parvidens | |
| Megistopoda aranea | 34 23 100 | Artibeus jamaicensis |
| 0 4 1 | Artibeus lituratus | |
| 0 0 1 | Phyllostomus discolor | |
| 0 0 1 | Platyrrhinus helleri | |
| 2 0 3 | Sturnira hondurensis | |
| Megistopoda próxima | 0 0 10 | Artibeus jamaicensis |
| 0 0 1 | Glossophaga commissarisi | |
| 4 8 29 | Sturnira hondurensis | |
| 5 6 8 | Sturnira parvidens | |
| Metelasmus pseudopterus | 1 0 2 | Artibeus jamaicensis |
| Nycterophilia coxata | 0 0 2 | Artibeus jamaicensis |
| 0 0 1 | Leptonycteris yerbabuenae | |
| Nycterophilia mormoopsis | 0 1 0 | Mormoops megalophylla |
| Paratrichobius dunni | 2 0 0 | Dermanura tolteca |
| Paratrichobius longicrus | 0 2 1 | Artibeus jamaicensis |
| 12 27 6 | Artibeus lituratus | |
| 0 1 4 | Sturnira hondurensis | |
| Paratrichobius lowei | 0 1 0 | Artibeus lituratus |
| Paratrichobius salvini | 1 0 1 | Platyrrhinus helleri |
| Paratrichobius sanchezi | 3 0 5 | Platyrrhinus helleri |
| 0 0 1 | Uroderma bilobatum | |
| Periglischrus caligus | 0 0 5 | Glossophaga commissarisi |
| 0 0 5 | Glossophaga soricina | |
| Periglischrus herrerai | 0 0 4 | Artibeus jamaicensis |
| 0 1 1 | Artibeus lituratus | |
| 0 0 1 | Sturnira parvidens | |
| Periglischrus iheringi | 132 15 132 | Artibeus jamaicensis |
| 16 42 18 | Artibeus lituratus | |
| 7 0 0 | Dermanura tolteca | |
| 0 0 1 | Dermanura watsoni | |
| 1 0 4 | Platyrrhinus helleri | |
| 4 0 0 | Uroderma bilobatum | |
| Periglischrus ojastii | 0 0 1 | Pteronotus parnellii |
| 0 1 8 | Sturnira hondurensis | |
| 0 4 4 | Sturnira parvidens | |
| Periglischrus torrealbai | 0 0 25 | Phyllostomus discolor |
| Speiseria ambigua | 0 0 4 | Carollia perspicillata |
| 0 0 1 | Carollia sowelli | |
| Spelaeorhynchus praecursor | 0 0 4 | Artibeus jamaicensis |
| 0 0 2 | Carollia perspicillata | |
| Strebla harderi | 0 0 1 | Carollia perspicillata |
| Strebla hertigi | 0 0 1 | Artibeus jamaicensis |
| 0 0 2 | Carollia sowelli | |
| 0 0 9 | Phyllostomus discolor | |
| Strebla wiedemanni | 2 0 13 | Desmodus rotundus |
| Trichobioides perspicillatus | 0 0 54 | Phyllostomus discolor |
| Trichobius assimilis | 17 9 54 | Artibeus jamaicensis |
| 6 5 0 | Artibeus lituratus | |
| 2 0 0 | Sturnira hondurensis | |
| Trichobius caecus | 3 0 8 | Pteronotus parnelli |
| Trichobius costalimai | 0 0 187 | Phyllostomus discolor |
| Trichobius galei | 0 0 1 | Pteronotus davyi |
| Trichobius handleyi | 0 0 6 | Glossophaga commissarisi |
| 0 3 2 | Glossophaga soricina | |
| Trichobius joblingi | 5 0 18 | Carollia perspicillata |
| 0 0 9 | Carollia sowelli | |
| Trichobius keenani | 0 0 3 | Glossophaga commissarisi |
| 1 2 15 | Glossopha soricina | |
| Trichobius parasiticus | 6 0 91 | Desmodus rotundus |
| Trichobius sphaeronotus | 0 0 2 | Leptonycteris yerbabuenae |
| Trichobius tuttlei | 0 1 1 | Artibeus jamaicensis |
| 0 0 1 | Phyllostomus discolor | |
| Total | 326 177 930 |
(ATE = Abundancia total de ectoparásitos, P = Pastizal, S = Silvopastoril, A = Acahual)
El esfuerzo de muestreo de ectoparásitos representó una cobertura de la riqueza estimada de 99.5% (35 especies), por lo que la curva de acumulación de especies de ectoparásitos tendió a alcanzar la asíntota (Figura 1). El sitio con los valores de riqueza más altos, tanto en especies de murciélagos como de ectoparásitos, fue el Acahual 3, además de ser el sitio con mayor diversidad de hospederos. El sitio con el valor de diversidad de especies de ectoparásitos más alto fue el Acahual 1 (Tabla 3). El acahual fue el tipo de paisaje que obtuvo los valores más altos de diversidad de especies de murciélagos y ectoparásitos. Cabe resaltar que el pastizal, que es el paisaje con mayor transformación antropogénica, presentó valores más altos de diversidad de hospederos y ectoparásitos en comparación con el paisaje silvopastoril. A través del cálculo del coeficiente de correlación de Pearson, se detectó una relación positiva, marginalmente significativa (r = 0.67, p = 0.06), entre los valores de diversidad de especies de hospederos y de diversidad de especies de ectoparásitos por sitio de muestreo.

Tabla 3 Análisis de la diversidad de hospederos y ectoparásitos por tipo de paisaje.
| Sitio | Riqueza de especies | Diversidad de especies (Shannon) | ||
|---|---|---|---|---|
| Murciélagos | Ectoparásitos | Murciélagos | Ectoparásitos | |
| Pastizal 1 | 9 | 7 | 1.54 | 1.57 |
| Pastizal 2 | 8 | 10 | 1.04 | 1.36 |
| Pastizal 3 | 11 | 12 | 1.93 | 1.89 |
| Silvopastoril 1 | 4 | 10 | 0.96 | 1.92 |
| Silvopastoril 2 | 6 | 11 | 1.28 | 1.93 |
| Acahual 1 | 11 | 20 | 2.03 | 2.45 |
| Acahual 2 | 11 | 16 | 1.31 | 1.81 |
| Acahual 3 | 18 | 23 | 2.09 | 2.24 |
En el paisaje de tipo pastizal se registraron 17 especies de ectoparásitos, de las cuales Paratrichobius dunni fue exclusiva para este paisaje. Para el paisaje de tipo silvopastoril se registraron 15 especies, dos de ellas exclusivas: Nycterophilia mormoopsis y P. lowei. Para el paisaje de tipo acahual se registraron un total de 11 especies exclusivas y presentó el valor más alto de riqueza de especies ectoparásitas (31 especies; Tabla 2). De todas las especies hospederas, únicamente A. jamaicensis, A. lituratus, S. hondurensis y S. parvidens fueron capturados en todos los sitios de muestreo. Con base en los valores de abundancia relativa, se consideró a A. jamaicensis, A. lituratus y S. hondurensis como las especies mejor representadas en el muestreo (Figura 1). A. jamaicensis obtuvo los valores más altos de diversidad de especies de ectoparásitos, seguido por S. hondurensis y en tercer lugar A. lituratus. La comunidad de ectoparásitos para los 209 hospederos de la especie A. jamaicensis estuvo integrada por 645 ectoparásitos, de tres familias: Streblidae (10 especies), Spinturnicidae (dos especies), Spelaeorhynchidae (una especie), con la mayor riqueza de ectoparásitos asociados. Las especies ectoparásitas más abundantes en los tres tipos de paisaje fueron Periglischrus iheringi (Spinturnicidae), que dominó en acahual y pastizal, Trichobius assimilis (Streblidae) y Megistopoda aranea (Streblidae), esta última la más abundante en silvopastoril. En el caso de la composición de la comunidad de ectoparásitos asociados con A. lituratus, de los 66 hospederos parasitados se obtuvieron 142 ejemplares pertenecientes a dos familias: Streblidae (cinco especies) y Spinturnicidae (dos especies). Las especies más abundantes para los tres tipos de paisaje fueron Periglischrus iheringi (Spinturnicidae) y Megistopoda aranea (Streblidae). La comunidad de ectoparásitos de los 39 hospederos de la especie S. hondurensis estuvo compuesta por 80 ectoparásitos de dos familias: Streblidae (siete especies) y Spinturnicidae (una especie). Megistopoda proxima (Streblidae) fue la más abundante en los tres tipos de paisaje. La composición de las comunidades de ectoparásitos en los hospederos generalistas de hábitat se muestra en la Figura 2.

Para el ANCOVA sólo se consideró a A. jamaicensis como especie hospedera, debido a que fue la única que presentó más de una especie de ectoparásito en cada sitio de muestreo. La interacción entre el tipo de paisaje y el rango de las especies ectoparásitas demostró un efecto estadísticamente significativo sobre la abundancia relativa de los ectoparásitos (F = 7.034 y p = 0.005). Los resultados de la diversidad beta (Morisita-Horn) entre las comunidades de ectoparásitos (por tipo de paisaje) muestran en general una alta similitud para cada especie hospedera generalista. Las comunidades de ectoparásitos de S. hondurensis presentaron valores altos de similitud de acuerdo con su composición, agrupándose en la misma rama, sin importar el tipo de paisaje (Figura 3). En cuanto a la relación entre las abundancias relativas de especies ectoparásitas con las especies hospederas generalistas de hábitat, la más fuerte fue una relación negativa entre la abundancia relativa de Paratrichobius longicrus y la abundancia relativa de Periglischrus iheringi en A. lituratus (r = -0.64yp< 0.0001).
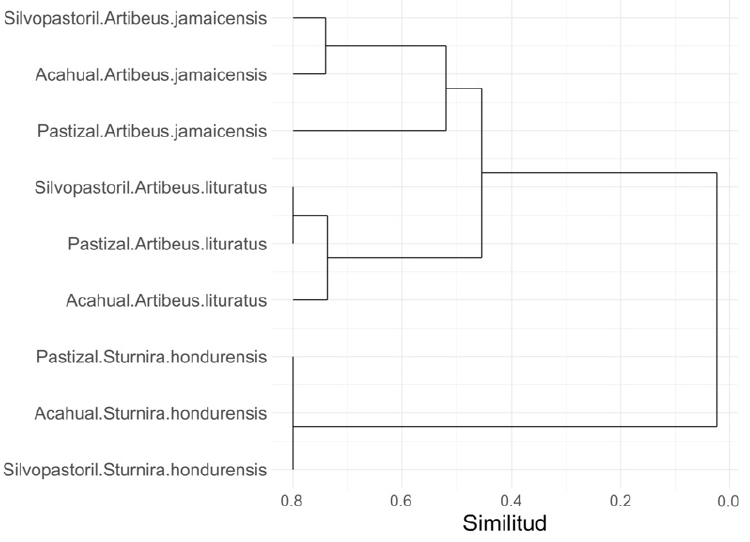
Figura 3 Dendograma de similitud de Morisita-Horn de las comunidades de ectoparásitos por tipo de paisaje de los tres hospederos generalistas (A. jamaicensis, A. lituratus y S. hondurensis).
La comparación de los valores de AF de las siete especies de estréblidos representados en todos los tipos de paisaje. Los valores de AF calculados por frecuencia muestran que la mayoría de los ectoparásitos tienen una elevada asociación filogenética. Sin embargo, los valores de AF fueron similares en los tres tipos de paisaje, es decir, no se observó diferencia significativa entre las medias de los valores de AF en estas siete especies de ectoparásitos de murciélagos muestreados en diferentes tipos de paisaje. A excepción de Trichobius assimilis cuyo valor de AF mostró un patrón de asociación filogenética significativamente distinto entre acahual y los sitios silvopastoril y pastizal.
DISCUSIÓN
En las últimas cuatro décadas, los ecosistemas del estado de Veracruz han sido modificados sustancialmente por las actividades antropogénicas, entre las que destaca la ganadería por su amplia distribución y por el grado de transformación que imprime sobre los paisajes naturales a través del establecimiento de potreros y zonas de cultivo. Estas modificaciones en la estructura del paisaje no sólo impactan las características de la vegetación, sino que, además alteran la dinámica de las comunidades de fauna, tanto de vertebrados como de invertebrados. Por lo tanto, los cambios en la composición de las comunidades reflejan el efecto que tiene la modificación de la estructura del paisaje en regiones con alta actividad pecuaria (Gonçalves et al. 2017, Newbold et al. 2015).
Si bien la relación positiva entre la diversidad de hospederos y la diversidad de ectoparásitos resulta lógica al considerar que conforme aumenta la captura de especies de murciélagos, se incrementa la probabilidad de acumular más hospederos y, por ende, mayor cantidad de especies ectoparásitas (Krasnov et al. 2004), es importante estudiar los efectos de la composición de las comunidades de especies hospederas sobre la composición de las comunidades de ectoparásitos asociados. La composición de las comunidades de especies hospederas en paisajes transformados, junto con sus patrones de distribución y de asociación con ciertos parásitos, ha sido ampliamente investigada. Sin embargo, existen pocos trabajos que presten atención a la composición, a la especificidad filogenética y a los patrones de organización de las comunidades de parásitos, especialmente las de ectoparásitos en murciélagos, en paisajes modificados por el hombre (Tello et al. 2008).
Las comunidades de hospederos y parásitos son sensibles a las condiciones del ambiente externo, incluida la perturbación antropogénica (Pilosof et al. 2012). Por ello, variables como la abundancia, riqueza, diversidad y el grado de infestación de los parásitos, pueden responder a cambios en el tamaño de las colonias, la masa corporal de los hospederos y factores asociados a la fragmentación del hábitat. Sin embargo, sólo se ha demostrado el efecto de la fragmentación del hábitat sobre la riqueza de especies de helmintos y virus asociados a murciélagos (Gay et al. 2014, Rico-Chavez et al. 2015).
El análisis de la diversidad y riqueza de las comunidades de ectoparásitos muestra que los valores más altos se registraron en sitios conformados principalmente por acahual. Los acahuales suelen estar asociados a paisajes más heterogéneos que se caracterizan por la presencia de vegetación secundaria, fragmentos remanentes de bosque maduro y zonas de cultivo o potreros (Pérez-Nicolás et al. 2018). Al respecto Froeschke et al. (2013) encontraron que la riqueza de especies de ectoparásitos fue más alta en sitios fragmentados que en sitios conservados, atribuyendo este patrón a la heterogeneidad del paisaje. Pero las comunidades de ectoparásitos en nuestro trabajo están compuestas en gran medida por las mismas especies independientemente de las características del paisaje. De manera similar, Frank et al. (2016), reportan que los ensambles de ectoparásitos en las comunidades de murciélagos de su estudio fueron muy similares en composición entre los sitios que muestrearon, los cuales presentan diferente grado de deforestación. Se ha demostrado que la composición del paisaje tiene un gran peso en el valor ecológico del mismo ya que provee de diversos recursos, como cobertura y alimento a las especies que transitan entre los pastizales, ayudando a mantener la diversidad del área (Daily et al. 2014).
Las especies ectoparásitas Periglischrus iheringi, Trichobius assimilis, Trichobius tuttlei, Aspidoptera phyllostomatis y Megistopoda aranea fueron compartidas entre los tres diferentes tipos de paisaje agropecuario del estudio. Dado que las especies de murciélagos generalistas de hábitat dominan en paisajes fragmentados, la estructura de la comunidad de los ectoparásitos asociados a estos sitios reflejó tal dominancia. Sin embargo, las comunidades de ectoparásitos de las tres especies de murciélagos generalistas de hábitat no mostraron diferencias significativas en cuanto a su composición por tipo de paisaje y únicamente se observó una variabilidad en cuanto a las abundancias relativas de las especies ectoparásitas. De manera similar, Frank et al. (2016) determinaron que la cobertura forestal no es un predictor significativo para el análisis de la abundancia y la composición de las especies ectoparásitas asociadas a murciélagos.
En estudios realizados en Chiapas y Yucatán se reportaron valores de prevalencia de ectoparásitos por sitio entre 11 y 85% (Cuxim-Koyoc et al. 2018, Tlapaya-Romero et al. 2015). La prevalencia de murciélagos parasitados en el presente trabajo fue del 45.34%. Estas diferencias pueden estar relacionadas con las condiciones ambientales de cada localidad (temperatura, humedad, altitud y precipitación), el esfuerzo de muestreo (específicamente la cantidad de murciélagos capturados) y las variaciones en las comunidades de hospederos (Lourenço et al. 2016). Por otro lado, Patterson et al. (2007) sugieren que los hábitos de refugio pueden impactar significativamente en los niveles de parasitismo. La prevalencia, la abundancia y la riqueza de especies de ectoparásitos en los hospederos aumenta en especies hospederas que forman colonias de muchos individuos y que se refugian en sitios que mantienen la temperatura y humedad ideales para completar el ciclo de vida de los parásitos, como es el caso de las especies de murciélagos que se refugian en cuevas. En el presente estudio, las especies de murciélagos Centurio senex, Chiroderma salvini, Chiroderma villosum, Dermanura phaeotis y Micronycteris microtis no presentaron ectoparásitos. Estos casos, en los que no existen registros de ectoparasitismo, pueden deberse a los hábitos de cada especie, por ejemplo, el constante recambio del refugio o simplemente el pequeño tamaño de sus colonias.
Algunos estudios en murciélagos han descrito relaciones negativas entre diferentes especies de ectoparásitos a través del análisis de la correlación entre sus densidades (Tello et al. 2008). En el estudio se encontró fuerte relación negativa entre la abundancia relativa de los ectoparásitos Paratrichobius longicrus (Trichobiinae) y Periglischrus iheringi (Spinturnicidae) en la especie hospedera Artibeus lituratus. Esta relación podría ser el resultado de interacciones competitivas entre estas especies, ya que ambas tienden a ocupar las superficies del hospedero desprovistas de pelo. Este tipo de interacciones podría presentarse entre otras especies, pues se ha documentado que las especies de ectoparásitos tienen preferencia por zonas específicas de la superficie del cuerpo de los murciélagos. Por ejemplo, las especies de la subfamilia Streblinae tienen preferencia por superficies del cuerpo cubiertas de pelo, mientras que las especies que pertenecen a la subfamilia Trichobiinae tienden a ubicarse con mayor frecuencia en las membranas de las alas al igual que las especies de ácaros de la familia Spinturnicidae (Ter Hofstede y Fenton 2005).
La especificidad filogenética, representada mediante el valor de AF, no mostró cambios significativos entre los distintos paisajes. Sin embargo, se considera que podría existir un efecto sobre la especificidad filogenética de los ectoparásitos por efectos ambientales (Dick y Dittmar 2014). Otros estudios mencionan que las especies hospederas preferidas por las moscas parásitas no corresponden con las especies consideradas adecuadas, entendiéndose a éstas últimas como las especies de hospedero en las que el parásito tiene el mayor éxito parasitario (Wenzel et al. 1966). Con tiempo y oportunidades de adaptación suficientes, el hospedero más adecuado podría convertirse en algún momento en el hospedero preferido en un área delimitada, lo que destaca que estos procesos se encuentran en constante dinamismo, con variantes espaciales y temporales.
Conocer los efectos de la transformación ambiental en las comunidades de sus ectoparásitos es primordial para el análisis y comprensión del impacto que generan las actividades antropogénicas en los agroecosistemas. En consecuencia, la consideración de factores ambientales que afecten la riqueza, abundancia y diversidad de los ectoparásitos, así como la especificidad filogenética con sus hospederos, es esencial para profundizar en el estudio de la relación hospedero-parásito. Este trabajo es pionero en la incorporación de características de paisajes fragmentados por la actividad pecuaria en el análisis de la composición de comunidades de ectoparásitos y su relación filogenética con especies de vertebrados de importancia ecológica como los murciélagos. De manera similar a lo planteado por Froeschke et al. (2013), por lo que las inferencias precisas sobre el efecto de las transformaciones del hábitat en las abundancias de los parásitos sólo serán posibles cuando las historias de vida del hospedero y parásito se estudien e involucren a fondo en el análisis.
CONCLUSIONES
La composición y configuración del paisaje pueden proporcionar información importante sobre las relaciones entre murciélagos y ectoparásitos en sitios perturbados por la actividad pecuaria, un ecosistema que anteriormente se había estudiado con poca frecuencia. El nivel de transformación de los paisajes agropecuarios definido por una variable categórica tiene un efecto sobre la composición de las comunidades de ectoparásitos asociadas con A. jamaicensis. El acahual, es el paisaje con menor transformación antropogénica, presentó los valores más altos de diversidad de especies de murciélagos y ectoparásitos. Los patrones de especificidad de hospedero parecen responder en mayor medida a la asociación parásito-hospedero. Por lo tanto, la composición de las comunidades de ectoparásitos depende en mayor medida de las especies hospederas presentes en la región de estudio. Se necesitan estudios que profundicen en la caracterización cuantitativa de paisajes antropizados debido a la actividad pecuaria, a fin de no depender de un sitio conservado de referencia, para facilitar las comparaciones entre sitios con paisajes transformados por actividades antropogénicas.

