











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La familia Eleotridae, se componen de alrededor de 150 especies de distribución mundial, que habitan en aguas marinas, salobres y dulces, Dormitator latifrons es representante de esta familia, conocido como dormilón gordo del Pacífico, chopopo, chame, puyeque o popoyote (Nordlie y Haney 1993). Es un pez originario de climas templados y subtropicales, tiene valor comercial en las costas de Ecuador, además que se le puede encontrar en ríos, manglares y en lugares tan extremos como charcas o fuera del agua mientras haya humedad (Vicuña 2010, López-Huerta et al. 2018). Una característica de esta especie es su alta resistencia fisiológica que le permite sobrevivir en lugares donde se presentan concentraciones bajas de oxígeno disuelto, amplio intervalo de temperatura (Castro-Rivera et al. 2005), variaciones de salinidad (Chang 1984), encontrándose de forma frecuente en aguas salobres (Yáñez-Arancibia y Díaz-González 1977).
En su medio natural D. latifrons puede experimentar grandes variaciones de salinidad, pero se tiene poca información sobre la capacidad para enfrentar el estrés osmótico (Todd 1975). La presión osmótica interna de los peces varía dependiendo en el ambiente acuático en el que se desarrollen; los peces de agua dulce poseen una concentración osmótica superior a la de su medio, mientras que en los peces marinos las sales, en los fluidos corporales, se encuentran más diluidas que el agua de mar (Christensen et al. 2017). La osmorregulación es la forma en la que los organismos pueden controlar o mantener estable su medio interno, cuando se exponen a diversas salinidades ambientales, algunos peces solo toleran salinidades similares en las que residen (Kültz 2015). A las especies que tiene la capacidad de sobrevivir dentro de ciertos intervalos de salinidades se les denomina eurihalinas y estas pueden ser de dos tipos, las que toleran y se adaptan a fluctuaciones rápidas de la salinidad externa y las que realizan migraciones en algún momento de su ciclo vital entre medios dulceacuícolas y marinos (Nordlie y Haney 1998). Gran parte del trabajo sobre las respuestas osmorreguladoras se ha realizado principalmente en salmónidos (Serrano et al. 2011). Basado en lo anterior, en este trabajo se evaluó la tolerancia fisiológica al cambio de salinidad, bajo condiciones de laboratorio, de D. latifrons, con el objetivo de aportar conocimiento sobre la capacidad de osmorregulación bajo condiciones controladas, y establecer las condiciones ambientales en las que puede ser mantenida en cultivo.
Materiales y métodos
Para el desarrollo del estudio se colectaron 200 organismos juveniles de D. latifrons (Laguna El Quelele, Nayarit, México), con peso 10.0 ± 2.0 g y talla 13.1 ± 2.2 cm, los cuales se transportaron en un contenedor de 1 000 L con aireación constante al Laboratorio de Calidad de Agua y Acuicultura Experi mental (LACUIC) del Centro Universitario de la Costa de la Universidad de Guadalajara, en Puerto Vallarta, Jalisco, México. Los peces se transfirieron a un estaque de 1 700 L para cuarentena y mantenimiento a salinidad de 22 ups (unidades prácticas de salinidad), misma salinidad a la que se encontraban en el medio natural. Durante este periodo se suministró un ali mento balanceado comercial con 35% proteínas y 8% lípidos, aireación constante y temperatura de 24.0 ± 1.0 °C.
Una vez transcurrido el periodo de cuarentena y adaptación al alimento balanceado, se seleccionaron 100 organismos al azar y se colocaron en dos unidades experimentales (UE) de 450 L (50 organismos por UE) con peso y longitud promedio de 8.5 ± 1.0 g y 9.5 ± 1.0 cm respectivamente. Previo a la colocación de los organismos se realizó un ajuste a la salinidad para mantener salinidades de 0.3 y 33 ups. La modificación de la concentración salina del agua se realizó de manera paulatina incrementando y/o disminuyendo 0.5 ups por hora hasta alcanzar la salinidad deseada partiendo de la salinidad de aclimatación. Durante el periodo del cambio salino se registró la sobrevivencia.
Durante 30 días los peces se mantuvieron a las salinidades ajustadas y se alimentaron con el alimento mencionado previamente, aireación constante y recambio de agua del 20% semanal. De los tratamientos de 0.3 y 33 ups se seleccionaron al azar seis organismos con peso y longitud promedio de 10.3 ± 0.5 g y 11.5 ± 1.0 cm respectivamente, los cuales se colocaron de forma individual en acuarios de 20 L, para luego medir la frecuencia ventilatoria branquial (FVB) por medio de los movimientos operculares por minuto, para obtener la frecuencia ventilatoria branquial basal por minuto (FVB basal).
Para determinar la capacidad de los peces de soportar cambios bruscos de salinidad, se seleccionaron al azar tres organismos que se encontraban en el tratamiento de 0.3 ups y se colocaron de forma individual en acuarios de 40 L, aforados a 20 L de agua a 15 ups, la cual se obtuvo de la mezcla de agua de mar (33 ups) con agua dulce. Los peces se introdujeron de forma directa a la salinidad ajustada, y se mantuvieron por 5 minutos (tiempo de estabilización) para posteriormente proceder a medir la FVB por 60 segundos. Las medidas se realizaron a las 1, 4, 24 y 48 h con un contador manual. La misma metodología se llevó a cabo para determinar la FVB de los peces transferidos de 0.3 a 25 y de 0.3 a 33 para los peces acondicionados a 33 ups y transferidos a 25, 15 y 0.3 ups.
Análisis estadístico
Para determinar las diferencias entre los tratamientos, se realizó una prueba de homogeneidad para después realizar un análisis de varianza. El nivel de significancia se estableció con una p < 0.05. Cuando se detectaron diferencias estadísticas, se utilizó una prueba post hoc de Dunnett para identificar diferencias estadísticas entre tratamientos. Los análisis se realizaron con el programa estadístico GraphPad Prism® 6.01.
Resultados y discusión
En el medio natural los organismos acuáticos tienen la capacidad de trasladarse de un lugar a otro hasta encontrar las condiciones fisiológicas adecuadas, cuando las variables fisicoquímicas del agua cambian de manera negativa para su óptimo desarrollo. Pero en condiciones de cultivo los organismos están confinados en espacios restringi dos y no pueden desplazarse hacia mejores condiciones ambientales y se genera un estrés que puede desencadenar la aparición de enfermedades o la muerte. El estrés es una condición en la cual el equilibrio dinámico u homeostasis es afectado, amenazado o perturbado por estímulos internos o externos (Corredor y Landines 2009). Los cuales pueden producir respuestas en las que interviene una serie de reacciones bioquímicas y fisiológicas que le permiten recuperar su homeostasis y adaptarse a una nueva situación. Lo que resulta en mayor estado de alerta y movilización de energía (Barreto y Volpato 2004, Blancas-Arroyo et al. 2014).
La FVB en los en peces se utiliza como indicador de estrés o alerta al reconocimiento visual de un depredador (Barreto et al. 2003), como indicador de sobrevivencia de peces o de estrés en ambientes con altos niveles de amonio (Blancas-Arroyo et al. 2014). También se ha reportado que la FVB es una respuesta muy sensible a la perturbación y una técnica no invasiva que no requiere equipos sofisti cados para evaluar el bienestar o estados de estrés en los peces (Barreto y Volpato 2004). Al respecto se observó que en condiciones de agua dulce con 0.3 ups, la FVB en juveniles de D. latifrons tuvo un promedio de 50.7 ± 4.2 ventilaciones por minuto, pero al modificar la salinidad, se tuvieron diferen cias estadísticas significativas entre tratamientos (p ≤ 0.05) (Figura 1). Los tratamientos de 0.3-15 y 0.3-33 ups tuvieron un promedio de 75.3 ± 5.7 y 103.3 ± 20.2 ventilaciones por minuto, respectivamente; durante el primer minuto de haber sido expuestos al choque osmótico con diferencias estadísticas significativas en la FVB basal (Figura 1a). Después de una hora de la transferencia los peces, solo se observaron diferencias estadísticas entre el tratamiento 0.3-33 ups (Figura 1b). Para las horas subsecuentes la FVB de ningún tratamiento presentó diferencias estadísticas (Figura 1c, d y e).
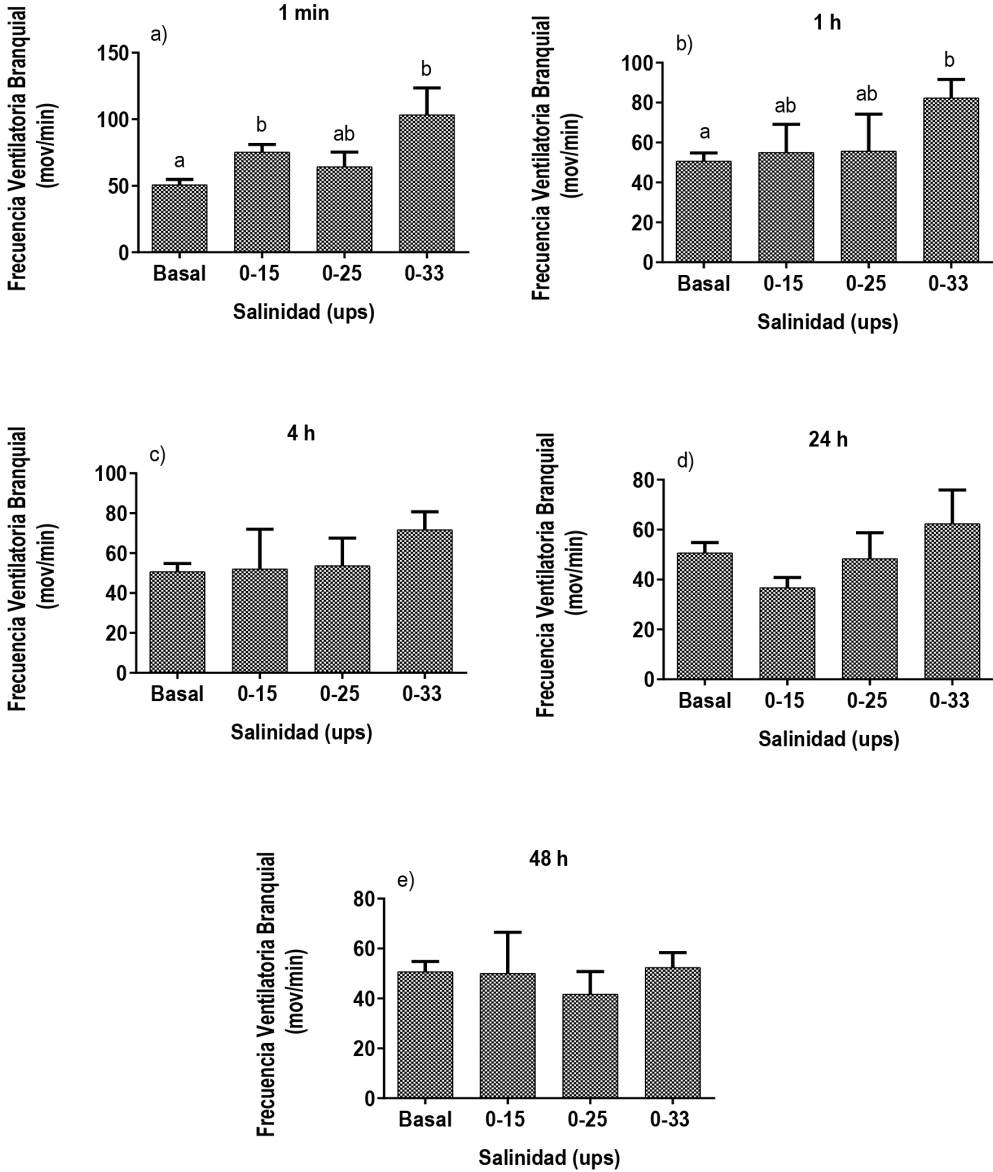
Figura 1: Frecuencia Ventiladora Branquial (FVB) de juveniles de D. latifrons transferidos de agua dulce hacia agua salada (0-15, 0-25, 0-33 ups), a), b), c), d) y e) FVB de los juveniles D. latifrons expuestos a los tratamientos con respecto al tiempo. Tratamientos con letras diferentes demuestran diferencias significativas (p < 0.05).
Sobre la capacidad fisiológica de D. latifrons para enfrentar los choques osmóticos, se reporta que el efecto de la salinidad de organismos capturados en agua dulce y transferidos a agua salada a concentraciones de 35, 40, 42, 45 y 50 ups, tuvieron una salinidad media letal de 42 ups (Chang 1984). Con respecto a la respuesta osmoadaptativa de D. latifrons, se reporta una supervivencia elevada en la transferencia directa de diferentes concentraciones de agua dulce al agua de mar (Becerra 1997). Lo que indica que este pez es capaz de hipoosmorregu- lar, lo que sugiere su carácter anfídromo, por lo que es capaz de emigrar desde y hacia medios marinos durante todo su ciclo de vida.
En relación con la FVB de los peces transferidos de agua salada (33 ups) ha agua dulce (0.3 ups), se observaron diferencias significativas. Los tratamientos 15-0.3, 25-0.3 y 33-0.3 ups tuvieron una FVB promedio de 79.9 ± 9.0, 94.3 ± 9.3 y 84.7 ± 7.4 ventilaciones por minuto, respectivamente; al primer minuto de haber sido expuestos al agua dulce, con diferencias significativas (Figura 2a). A los 60 min se encontraron diferencias estadísticas significativas (p ≤ 0.05) con los tratamientos15-0.3 y 33-0.3 ups con una FVB de 64.3 ± 4.0 y 79.7 ± 1.5 ventilaciones por minuto, respectivamente (Figura 2b). Por otra parte, se encontraron diferencias significativas a las 4, 24 y 48 h, en el tratamiento 33-0.3 ups con respecto a la FVB basal con valor de 67.0 ± 5.6 a las 4 h y de 58.3 ± 7.1 ventilaciones por minuto a las 24 y 48 h de haber sido expuestos al cambio (Figura 2c, d y e).
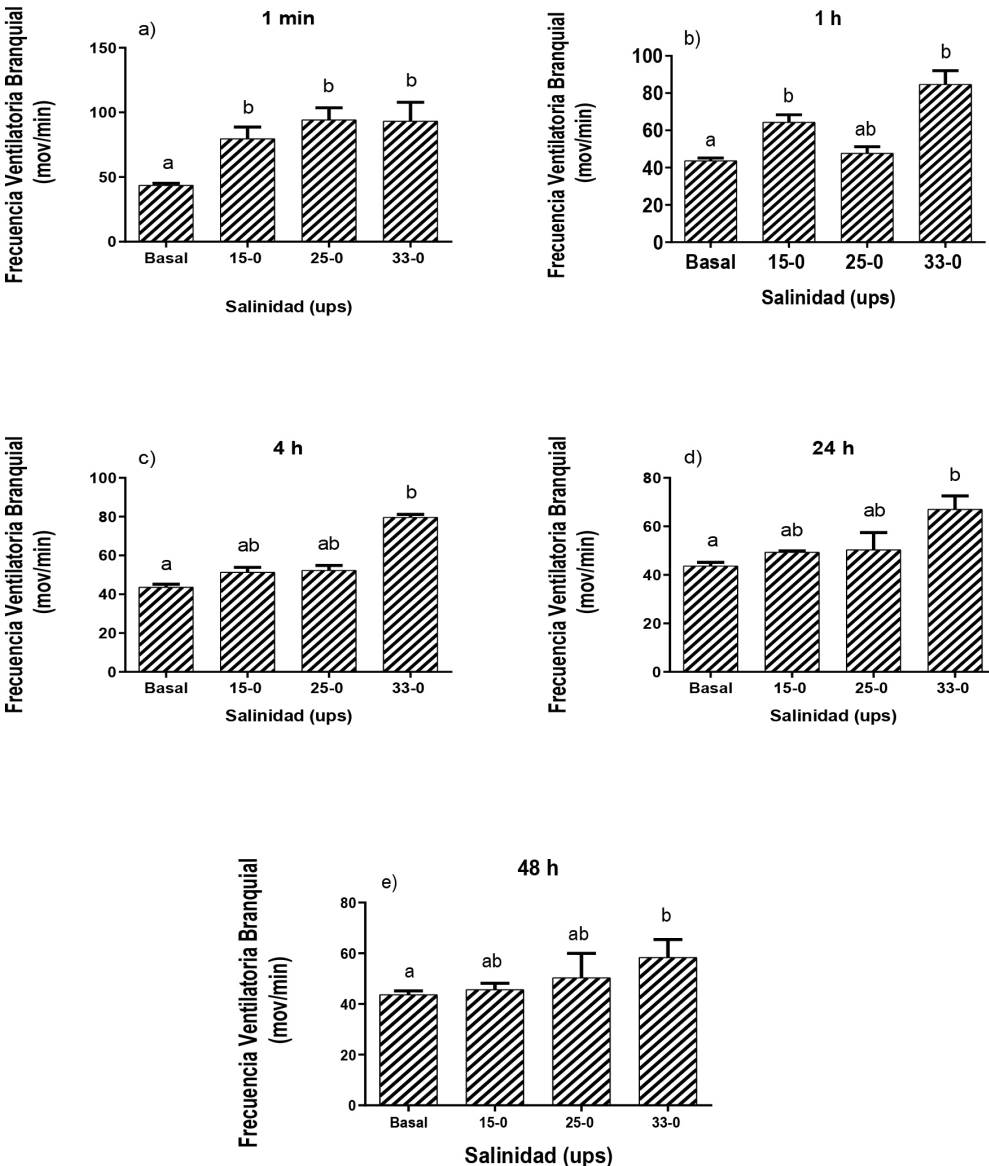
Figura 2: Frecuencia Ventilatoria Branquial (FVB) de juveniles de D. latifrons transferidos de agua salada hacia agua dulce (15-0, 25-0, 33-0 ups), a), b), c), d) y e) FVB de los juveniles D. latifrons expuestos a los tratamientos con respecto al tiempo. Tratamientos con letras diferentes demuestran diferencias significativas (p < 0.05).
Durante el proceso de aclimatación a distintas salinidades como en todos los casos de cambio salino se presentó un 100% de sobrevivencia. Lo que sugiere una alta capacidad de D. latifrons adaptarse a concentraciones salinas variables incluso soportar cambios bruscos. Al respecto, se sabe que algunos peces eurihalinos tienen la capacidad fisiológica de adaptarse más rápido a los cambios en la salinidad del agua, compensando la situación acelerando el proceso de equilibrio osmótico (Bœuf y Payan 2001). Pero en el caso de que el proceso de adaptación no se lleve a cabo de manera rápida, el organismos entra en una fase de resistencia, en la que libera corticoides adrenales que pueden derivar en situaciones letales provocando la muerte de los organismos (Marrero-Hernández 2008). Al respecto, Nordlie y Haney (1993) evaluaron las tolerancias de salinidad y las concentraciones osmóticas de plasma para D. maculatus especie homóloga de D. latifrons en las costas del Atlántico americano, encontrando un límite de tolerancia entre 75 y 80 ups, con sobrevivencia del 47% de los peces sometidos a 80 ups. Sobre la tolerancia a la salinidad se sabe que la capacidad de un pez para aclimatarse a diferentes niveles de salinidad ambiental depende de su capaci dad para regular la absorción y la excreción de iones, y mantener su equilibrio hidromineral (Zhao et al. 2011, Blancas-Arroyo et al. 2014). Los juveniles de D. latifrons fueron capaces de sobrevivir durante 30 días, tanto bajo condiciones de agua dulce (0.3 ups) como marinas (33 ups), sin aparente estrés; incluso bajo condiciones hiperosmóticas, cuando los peces retornaron a la FVB basal desde la primera hora, lo que confirma su carácter eurihalino.
Los resultados del presente estudio coinciden con los resultados de Nordlie y Haney (1993), con relación a las concentraciones secuenciales a las que fueron expuestos y la capacidad de regresar a la FVB basal al ser transferidos de agua salada ha agua dulce y viceversa con un 100% de sobrevivencia. Por lo que D. latifrons en un pez que muestra alta capacidad fisiológica de adaptación a ambientes con cambios salinos, por lo que se confirma su carácter eurihalino y su importancia ecológica al poder ocupar diversos nichos tróficos. Además, tales características lo hacen un buen candidato para ser cultivado en agua dulce y agua salobre. Por lo que se requiere ampliar los estudios sobre el efecto de la salinidad en el crecimiento, costo energético y fisiológico de D. latifrons, para establecer los márgenes osmóticos óptimos en los que puede desarrollarse de manera adecuada y sin estrés.

