











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La fenología se refiere a la sincronización de los eventos biológicos recurrentes, al estudio de los mismos en relación con las fuerzas bióticas y abióticas que los causan, así como a la interrelación entre las fases de dichos eventos para la misma o diferentes especies (Lieth 1974). La fenología es un indicador importante del impacto de los cambios del clima sobre los ecosistemas terrestres (Richardson et al. 2013, Migliavacca et al. 2015), por lo que en los últimos años se ha renovado el interés por los estudios fenológicos, dada su relevancia para el estudio del cambio global (White et al. 2009, Cerdeira Morellato et al. 2016).Actualmente se acepta que la influencia entre la biósfera y la atmósfera es recíproca: el aumento de la temperatura del planeta y la mayor frecuencia de eventos extremos, como sequías e inundaciones, afectan la fenología de los biomas modificando las fechas de brotación, floración y fructificación de las plantas y la duración de las estaciones de crecimiento (Richardson et al. 2013), incluso la actividad fotosintética (Gamon et al. 2016).Esto a su vez modifica las características de las superficies naturales (rugosidad, humedad, albedo, entre otras), afectando el intercambio de agua, energía y CO2 (Arora y Boer 2005, Peñuelas et al. 2009). Al mismo tiempo, todos estos cambios alteran el microclima en los ecosistemas y tienen un efecto sobre la competencia entre los individuos y la estructura de las poblaciones (Richardson et al. 2013).
En el comportamiento de todo ecosistema existe una clara dependencia del crecimiento de la vegetación con la variación de las condiciones climatológicas y éstas a su vez están asociadas al relieve. Los factores ambientales que caracterizan un lugar pueden clasificarse en topográficos, climáticos y de suelo. La latitud, longitud y elevación pueden ser medidas indirectas del clima regional, mientras que la pendiente y la orientación indican el clima local (Socha 2008). El efecto de la elevación se manifiesta con dos fenómenos: la temperatura se reduce en las zonas altas, y con ella la evapotranspiración y la humedad se acumula en las zonas bajas (Gallardo- Cruz et al. 2009). La orientación es un factor que puede modificar las variables esenciales para el funcionamiento de las plantas, como la cantidad y calidad de radiación incidente, las temperaturas y la frecuencia de heladas (Torres et al. 2012); también influye en el balance hídrico, lo que condiciona la diversidad y estructura de la vegetación (Gallardo-Cruz et al. 2009). Por otra parte, la diversidad y densidad de vegetación disminuye con la pendiente (Nadal-Romero et al. 2014).
Inferir la información fenológica a partir de datos satelitales tiene la ventaja de asegurar repetitividad y cobertura espacial y temporal de las observaciones, en todos los ecosistemas del planeta (Peñuelas et al. 2009). El estudio del desarrollo espacio-temporal de las coberturas vegetales a partir de datos de teledetección se conoce como Land Surface Phenology; se relaciona con la fenología de las plantas, pero es diferente al enfoque con datos de campo, ya que considera el pixel (con todo lo que pueda contener) como unidad de medida (White et al. 2009, Zhang et al. 2017). La primera propuesta de representación del comportamiento fenológico de la vegetación a nivel global fueron los mapas mensuales de vegetación ECOCLIMAP (Masson et al. 2003).
Otra forma de representar la variación espacio-temporal de la vegetación, es a partir de la evolución estacional del índice de área foliar (Arora y Boer 2005), que corresponde al número de capas de superficie foliar por unidad de suelo (LAI: Leaf Area Index) y es considerado una variable climática esencial para entender la interacción entre las superficies naturales y la atmósfera, según el Global Climate Observing System (Fang et al. 2013). La primera serie de LAI fue CYCLOPES y su versión mejorada es la serie GEOV1 (Baret et al. 2013); ambas se desarrollaron dentro del proyecto Copernicus (antes GEOLAND) del programa Global Monitoring for Environment and Security de la Unión Europea, con 0.05o de resolución (1 km en el ecuador) y pasos de 10 días, a partir de datos SPOT/VGT (Smets et al.2013). Alcántara et al. (2013) indican que la información fenológica contenida en la serie de LAI de cada pixel de 1 km se puede extraer a partir del cálculo de variables fenológicas anuales (agregación anual o AAT por Annual Aggregated Time series), como son el inicio, final, mitad y duración de la estación de crecimiento de la vegetación, el máximo, el mínimo y la amplitud del LAI y su valor acumulado en ese periodo de tiempo (Valderrama-Landeros et al. 2014).Con las series de LAI se han podido obtener las climatologías de LAI, que son mapas globales del promedio del LAI en un conjunto de varios años, para cada pixel de 1 km. A nivel global destaca la climatología GEOCLIM-LAI, calculada a partir de la serie GEOV1 (1999 - 2010) (Verger et al. 2016). La utilización de climatologías de LAI en los modelos climáticos globales, en comparación con el uso de un valor de LAI constante, ha contribuido a mejorar las estimaciones fenológicas (Verger et al. 2016). La representación del ciclo anual de la vegetación con una climatología puede servir como línea de base para establecer la condición normal de un ecosistema (Verger et al. 2013).
Los estudios fenológicos revisten especial interés en las zonas protegidas, como la Reserva de la Biósfera Mariposa Monarca (RBMM), donde los cambios en la fenología del bosque pueden afectar la supervivencia de las mariposas, cuya población ha disminuido en los últimos años. Se sabe que las mariposas son susceptibles a los cambios de temperatura y humedad (Brower et al. 2011), en particular a las heladas nocturnas (Barve et al. 2012), eventos que podrían aumentar como consecuencia del cambio climático (Oberhauser y Peterson 2003, Islas-Báez et al. 2015). Existen estudios de seguimiento fenológico de algunas especies vegetales de la RBMM (Cornejo-Tenorio e Ibarra-Manríquez 2007); sin embargo, no se ha realizado una caracterización fenológica del bosque a partir de las bases globales de LAI. El objetivo del trabajo fue caracterizar la fenología de la RBMM, considerando los diferentes tipos de coberturas presentes en la zona, a partir del análisis de su serie GEOV1 (2000 - 2015), y establecer la relación de dependencia de algún aspecto de la fenología con el clima local, representado éste indirectamente a través de la elevación y la exposición al sol de cada kilómetro cuadrado de la reserva.
Materiales y métodos
Zona de estudio
La Reserva de la RBMM, con una superficie de 56 259 ha, es una de las áreas protegidas de bosque templado más importantes de México, por ser el refugio invernal de la mariposa Danaus plexippus L.; además se caracteriza por una gran diversidad de plantas vasculares y una particular biogeografía,que incluye una combinación única de elementos de origen holártico y neotropical a una altitud elevada en una zona tropical (Cornejo-Tenorio et al. 2003). Se encuentra entre los estados de Michoacán y México, en la provincia fisiográfica Eje Neovolcánico, al extremo sur de la Altiplanicie Mexicana, al norte de la depresión del Balsas. Se trata de un sistema montañoso discontinuo, con fuertes pendientes y elevaciones de 1 800 a 3 640 m, con clima templado subhúmedo (lluvias en verano, precipitación media anual de 830 mm y temperatura media anual de 15.7oC), que constituye un área importante de captación fluvial (Giménez de Azcárate et al. 2003). .
La vegetación característica es bosque templado, con bosque de pino-encino (Pinus - Quercus) en las zonas de menor elevación, y oyamel (Abies religiosa (Kunth) Schltdl. & Cham.) en las de mayor, siendo este último el hábitat característico de la mariposa monarca; asimismo, se encuentran manchones de pastizales y zonas arbustivas, además de parcelas agrícolas. Los terrenos son en su mayoría propiedad de comunidades indígenas y ejidos, con niveles significativos de pobreza. A pesar de la categoría de reserva, en la zona son frecuentes los casos de tala ilegal; también ha habido problemas ambientales (vientos y deslaves) que han llevado a la tala autorizada de grandes cantidades de madera, con afectaciones al ecosistema (Leverkus et al. 2017).
Mapa de vegetación
Como referencia de la distribución de los tipos de vegetación de la zona de estudio, se utilizó el mapa de cobertura de la RBMM (Serie cartográfica Monarca: Volumen I. Vegetación y cubiertas de suelo) en versión impresa, que se digitalizó, georreferenció y reclasificó con el fin de resumir los ocho tipos de cobertura del mapa, en cuatro: oyamel (oyamel y oyamel perturbado), pino (pino-encino, encino, pino-encino perturbado y encino perturbado), arbustos (pastizal inducido y arbustos secundarios) y cultivos (cultivos y plantaciones). Se aplicaron sucesivos filtros para eliminar artefactos y pixeles aislados; el resultado se convirtió a la proyección (UTM WGS84) y resolución de la serie de tiempo de LAI de GEOV1.Para cada tipo de cobertura se obtuvo el número de pixeles, valor que constituye una aproximación de la superficie que ésta ocupa. El proceso se realizó con el programa ENVI, versión 4.8.
Datos de clima
Se utilizaron los datos globales de temperatura y precipitación de WORLDCLIM, versión 1 (Hijmans et al. 2005), recortados a la zona de estudio. Esta base de datos es el primer modelo climático en forma de superficies climatológicas de alta resolución (1 km, los anteriores eran de 6 km o más) y consiste en mapas mensuales (superficies climáticas), con resolución de 1 km, correspondientes a las temperaturas media, mínima y máxima promedio de 1950 al 2000, registradas por estaciones meteorológicas en tierra e interpoladas espacialmente con ayuda de un modelo digital de elevación(no se consideran orientaciones ni pendientes); las temperaturas máximas y mínimas se estiman como la temperatura media, más o menos la mitad del intervalo medido de temperaturas. Se calculó el promedio de cada variable meteorológica, para cada mes y cada tipo de vegetación, a partir del mapa de cobertura de referencia, utilizando los programas ENVI,versión 4.8 y MatLab, versión R2015a.
Datos de relieve
A partir del continuo de elevaciones de la zona de estudio del Instituto Nacional de Estadística y Geografía de México (INEGI), convertido a resolución de 1 km, se obtuvo la elevación, la pendiente y la orientación de cada pixel de 1 km. Para la orientación, se consideraron cuatro regiones: Norte (315o, 45o), Este (45o, 135o), Sur (135o, 225o) y Oeste (225o, 315o). Para cada tipo de vegetación, según el mapa de referencia, se estableció la distribución de las elevaciones y pendientes, así como de las orientaciones. A partir de la fórmula de Oke (Bennie et al. 2006) se calculó el índice de radiación, Ri, de cada pixel, como Ri = cos β cos Z + sin β sin Z cos (Ω - Ωs ), donde β es el ángulo de la pendiente, Z es el zénit solar (ángulo del Sol con la vertical), Ω es el azimut solar (ángulo del Sol con la dirección norte), Ωs es la orientación del terreno. Ri estima la proporción de irradianza potencial directa interceptada por una ladera, para una posición solar (cenital y azimutal) dada, y es un indicador de su nivel de iluminación. En este estudio se consideró la posición del Sol en el solsticio de invierno, con el fin de resaltar las diferencias de iluminación entre las laderas orientadas al Norte y al Sur.
Datos de LAI
Se utilizaron los datos de LAI generados de forma global por el programa europeo de monitoreo de la superficie terrestre Copernicus (Smets et al. 2013). Se trata de imágenes con una resolución espacial de 0.05o (1 km en el ecuador) y resolución temporal de 10 días, correspondientes al periodo 1999-2015 (36 imágenes por año, 612 datos por pixel), que cubren la superficie de México. Se recortó el área de estudio (100.16◦ - 100.37◦ W y 19.32◦ - 19.75◦ N) y se le aplicó una máscara del límite de la RBMM a 1 km. Los datos se ordenaron cronológicamente (repitiendo el último año y tomando el primero (1999) como margen inicial), formando una matriz de LAI (49 x 24 x 648). Los huecos de información, debidos principalmente a la presencia de nubes, se rellenaron calculando interpolaciones lineales en el tiempo. Se calculó la climatología de la RBMM como el promedio interanual del LAI de cada pixel para cada fecha de la serie (de 1999 a 2015, con pasos de 10 días); asimismo se obtuvo la desviación estándar asociada.
Variables fenológicas
Con el fin de obtener las variables fenológicas de cada pixel para cada año, se llevó a cabo el análisis de la serie completa con el programa TIMESAT (Jöhsson y Eklundh 2004), considerando un filtro adaptativo Savitsky-Golay, un tamaño de ventana del 20%, una estación por año y dos iteraciones. Para cada pixel y cada año de la serie (2000 a 2015), se obtuvieron 11 variables fenológicas: inicio, fin, longitud y mitad de la estación de crecimiento, mínimo, máximo y amplitud de LAI, derivadas izquierda y derecha, integral extendida e integral corta (Valderrama-Landeros et al. 2014). Los valores muy grandes (mayores de 36.5 para inicio y mitad de estación, y mayores de 50 para final y longitud de estación) o negativos no se tomaron en cuenta. Para cada una de las 11 variables fenológicas calculadas, en cada pixel se calculó el promedio interanual de la variable; para todos los pixeles de un mismo tipo de cobertura se calculó el promedio de los valores promedio de cada variable. También se calculó la desviación estándar asociada a este promedio general de cada variable para cada cobertura; este valor representa la variabilidad espacial de la variable dentro de la zona correspondiente a cada tipo de vegetación. Adicionalmente para cada cobertura se realizó un análisis de regresión múltiple entre los valores promedio de las variables fenológicas y la elevación y el índice de radiación, elementos que integran las características abióticas del lugar: temperatura, precipitación, insolación y humedad; se utilizó la función fitlm, con el método de Robust Fit del programa Matlab, para determinar el coeficiente de determinación, R2, así como la significancia del modelo y de los parámetros de regresión.
Resultados
La cobertura más abundante en la RBMM es el pino, con 216 pixeles, que corresponden al 37.43% de la zona de estudio (aproximadamente 20 000 ha),
seguido por el oyamel con 183 pixeles (31.72%, cerca de 17 000 ha), los cultivos con 107 pixeles (18.54%, 9 900 ha) y finalmente los arbustos, con 71 pixeles (12.31%, 6 500 ha). Para todas las coberturas el LAI es mínimo en los meses de enero a mayo; a partir de este momento, durante la etapa de crecimiento vegetativo, el LAI aumenta, hasta alcanzar el valor máximo entre agosto y octubre (Figura 1).
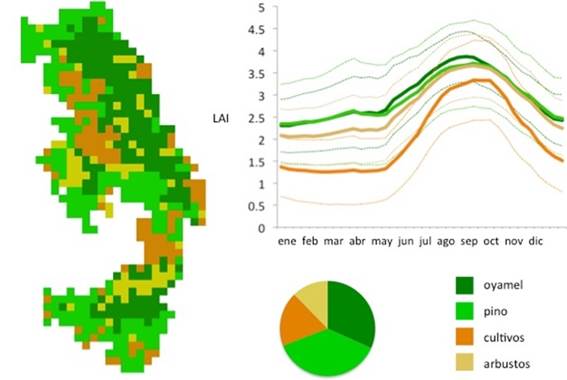
Figura 1 Mapa de vegetación de la RBMM a 1 km con 4 clases (oyamel, pino, cultivos, arbustos) (tamaño de píxel 1 km2). Distribución porcentual de la superficie ocupada por cada clase. Promedio y desviación estándar interanual del valor de índice de área foliar (LAI) de cada clase (climatología).
Cada ciclo anual se compone de una época de poca lluvia (secas) entre octubre y mayo, con el mínimo en el mes de marzo, y una época de lluvias entre junio y septiembre, donde la precipitación puede superar los 200 mm. La precipitación en el mes más seco (marzo) es mayor en la zona de oyamel que para las demás coberturas, mientras que en la época de lluvias hay menos variabilidad (Figura 2). En cuanto a las temperaturas (Figura 3), las medias varían entre 8 y 14 oC, presentándose las más bajas entre los meses de noviembre a febrero y las más altas en marzo y abril. Las temperaturas mínimas más bajas (1 oC) se producen entre noviembre y febrero, mientras que los valores de temperatura mínima más altos se alcanzan en la época de lluvias, entre junio y septiembre (8 oC). Las temperaturas máximas son mayores en la época de secas (21 oC) y menores durante el invierno (18 oC). Entre los tipos de cobertura, el oyamel siempre presenta las temperaturas más bajas, mientras que las zonas de pino y de cultivos presentan las más altas; en particular, la temperatura mínima en el mes más frío (enero), es más baja en la zona de oyamel.
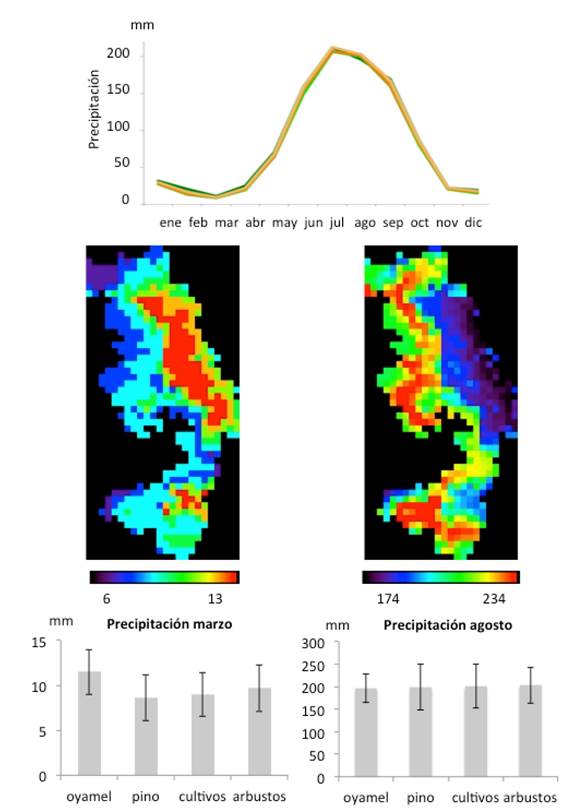
Figura 2 Precipitación promedio de cada tipo de cobertura en la RBMM (oyamel, pino, cultivos, arbustos); mapa de distribución de la precipitación en el mes más seco (marzo) y más lluvioso (agosto) (tamaño de píxel 1 km2), y promedio y desviación estándar de la precipitación interanual de cada cobertura en esos meses (datos de WORLDCLIM).
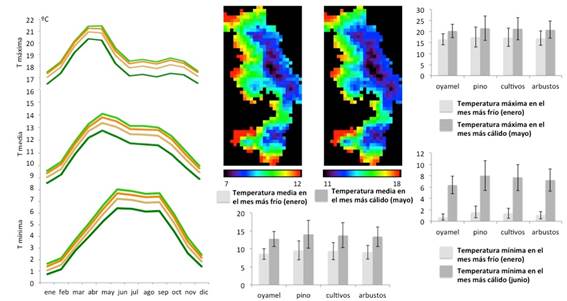
Figura 3 Temperatura mínima, media y máxima promedio de cada tipo de cobertura en la RBMM (oyamel, pino, cultivos, arbustos); mapa de distribución de la temperatura media en el mes más frío (enero) y más cálido (mayo) (tamaño de píxel 1 km2), y promedio interanual y desviación estándar de las temperaturas media, máxima y mínima en el mes más frío (enero) y más cálido (mayo), en cada cobertura (datos de WORLDCLIM).
La elevación va de los 2 200 a los 3 300 m (Figura 4). Las pendientes varían entre 2 y 12%, valores bajos debido al tamaño de pixel. Las mayores pendientes se dan en el centro, a lo largo de un eje que recorre la reserva de Norte a Sur, en la frontera entre los estados de Michoacán y México, correspondiendo a la zona núcleo, donde la cobertura predominante es oyamel; a partir de este eje las laderas caen hacia ambos lados Este y Oeste, principalmente hacia el Oeste. Hay zonas de alta pendiente en la parte centro-Oeste, donde hay pino, y en la zona Sur. Esto define la orientación de las laderas: en la mitad Norte de la reserva, el eje central de máxima altitud divide la zona en cuatro laderas: Este, Oeste, Norte y Sur, mientras que en la parte Sur de la reserva las orientaciones predominantes son Norte y principalmente Sur. En cuanto al índice de radiación, es mayor en las zonas orientadas al Sur y con fuertes pendientes. La distribución de las elevaciones muestra una clara semejanza con el mapa de precipitaciones del mes de marzo, lo que indica que la precipitación en la época más seca, y por lo tanto el mantenimiento de la humedad durante todo el año, depende principalmente de la elevación; sin embargo, el mapa de elevaciones es inverso con respecto al de temperaturas: las zonas de mayor elevación registran las menores temperaturas.
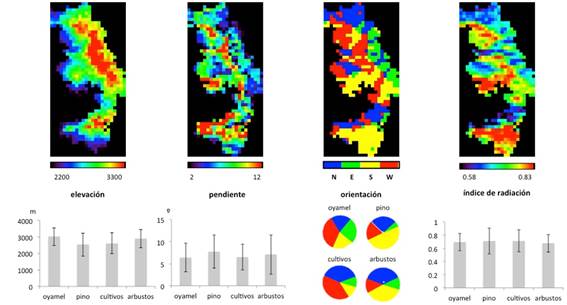
Figura 4 Mapas de distribución de la elevación, la pendiente, la orientación y el índice de radiación en la RBMM (tamaño de píxel 1 km2 ) y promedio y desviación estándar de la elevación, la pendiente y el índice de radiación y distribución de los pixeles por orientación, para cada tipo de cobertura.
La Figura 5 muestra los mapas de los promedios interanuales de las 11 variables fenológicas calculadas con TIMESAT; se aprecia que la variación de los diferentes aspectos fenológicos es congruente con los tipos de cobertura indicados en la Figura 1: la zona de oyamel, la zona de pino en el centro y el Sur y las zonas agrícolas en el Oeste. La Figura 6 muestra los promedios de las mismas variables, calculados entre todos los pixeles de una misma cobertura, y la desviación estándar espacial correspondiente; los cultivos presentan en general un comportamiento diferenciado del resto de las coberturas naturales; el pino tiene la mayor desviación que indica mayor variabilidad.
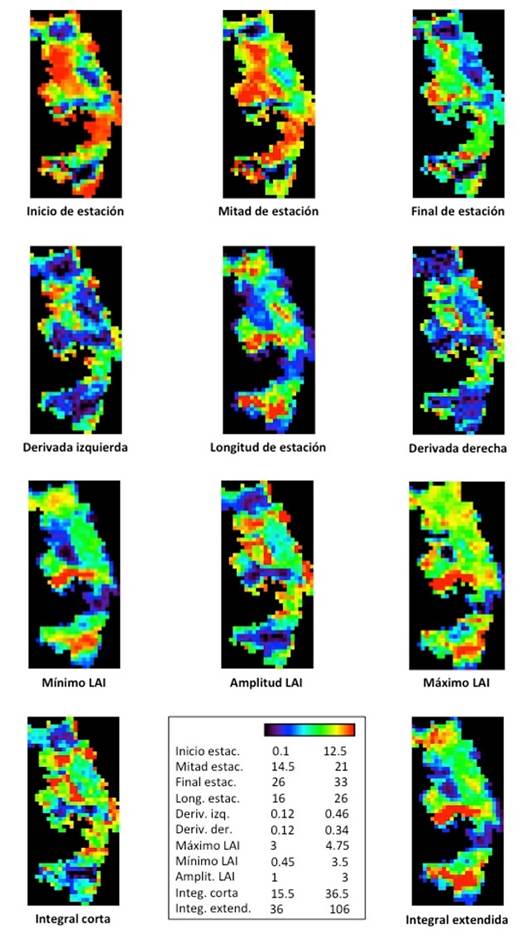
Figura 5 Mapas de los valores promedios interanuales de las variables fenológicas calculadas por TIMESAT en la RBMM (tamaño de píxel 1 km2)
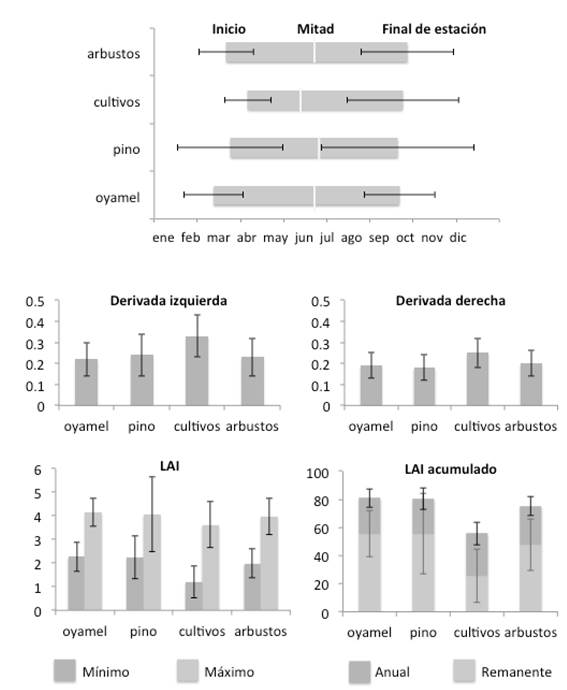
Figura 6 Promedios y desviación estándar de las variables fenológicas calculadas por TIMESAT, para cada tipo de cobertura en la RBMM (oyamel, pino, cultivos, arbustos). La desviación estándar representa la variación espacial.
Los resultados de las regresiones múltiples calculadas entre el promedio de cada una de las variables fenológicas de cada cobertura, y la elevación y el índice de radiación (que caracterizan el relieve) indican que son los cultivos (inicio de estación, mínimo de LAI, derivada derecha e integral corta) y los arbustos (inicio de estación) los que presentan una mayor dependencia del relieve, por las diferencias en elevación (Tabla 1).
Tabla 1 Regresión lineal múltiple (R2) entre el relieve (general (p), elevación (pE), índice de radiación (pRi)) y las variables fenológicas de cada tipo de cobertura en la RBMM, considerando el promedio de los promedios interanuales de cada una, en todos los pixeles de la misma cobertura.
| Variables fenológicas | oyamel | pino | cultivos | arbustos | ||||||||||||
| R2 | p | pE | pRi | R2 | p | pE | pRi | R2 | p | pE | pRi | R2 | p | pE | pRi | |
| Inicio de estación | 0.058 | ** | i** | ns | 0.113 | *** | i*** | ns | 0.216 | *** | i** | ns | 0.407 | *** | i*** | ns |
| Mitad de estación | 0.057 | ** | * | ns | 0.042 | * | ns | i** | 0.065 | * | * | ns | 0.193 | *** | i*** | ns |
| Final de estación | 0.116 | *** | i** | *** | 0.057 | ** | ns | ns | 0.055 | ns | ns | ns | 0.146 | ** | i** | ns |
| Longitud de estación | 0.017 | ns | Ns | ns | 0.144 | *** | ns | *** | 0.140 | ** | ns | ** | 0.022 | ns | ns | ns |
| Derivada izquierda | 0.031 | ns | Ns | ns | 0.147 | *** | i* | i*** | 0.062 | * | ns | ns | 0.088 | * | i* | ns |
| Derivada derecha | 0.142 | *** | i*** | 0.074 | *** | ns | i*** | 0.290 | *** | *** | i** | 0.042 | ns | ns | ns | |
| Mínimo | 0.128 | *** | i*** | *** | 0.142 | *** | ns | *** | 0.229 | *** | i*** | ** | 0.032 | ns | ns | ns |
| Máximo | 0.053 | ** | Ns | ns | 0.064 | ** | ns | ** | 0.055 | ns | ns | * | 0.053 | ns | ns | ns |
| Amplitud | 0.116 | *** | * | i*** | 0.147 | *** | ns | i*** | 0.327 | *** | *** | ns | 0.014 | ns | ns | ns |
| Integral corta | 0.132 | *** | * | i*** | 0.080 | *** | ns | i*** | 0.377 | *** | *** | ns | 0.006 | ns | ns | ns |
| Integral extendida | 0.028 | ns | Ns | * | 0.149 | *** | ns | *** | 0.113 | ** | ns | ** | 0.004 | ns | ns | ns |
***: relación altamente significativa (p≤0.001), **: muy significativa (p≤0.01), *: significativa (p≤0.05), i: inversamente proporcional. En negrita valores de R2 superiores a 0.2 (20 %); ns: no significativo
Discusión
El oyamel prospera en zonas con humedad atmosférica elevada y bajas temperaturas durante todo el año, preferentemente en laderas protegidas del viento y de la incidencia solar elevada (Castillo-Batista et al. 2018), condiciones que predominan en los bosques de Abies en México (Rzedowski 2006) y que se presentan en las zonas más elevadas de la RBMM.
Las condiciones ambientales relativamente estables a lo largo del año y la naturaleza perennifolia del estrato arbóreo, dan lugar a una baja estacionalidad de los bosques de oyamel, que se aprecia en la climatología (Figura 1). El ciclo fenológico comienza con la emergencia de las yemas vegetativas, que ocurre en marzo-abril, el desarrollo de los nuevos brotes inicia en abril-mayo, alcanzando su máximo en los meses de agosto y septiembre; a finales de junio comienza el desprendimiento de las hojas viejas, que han ido adquiriendo una coloración café ligeramente rojiza (Rzedowski 2006). El LAI mínimo es alto y puede llegar a ser cercano al máximo (desviación estándar alta), que presenta valores intermedios, con lo que la amplitud del LAI es baja. En conjunto la curva de la climatología del oyamel es poco pronunciada, provocando que una estación de crecimiento se empalme con la del siguiente año; esto puede explicar que el inicio, mitad y final de estación sean los más tempranos y ocurran con mayor lentitud que para las otras coberturas (derivadas izquierda y derecha, pequeñas); el crecimiento acumulado de la vegetación en un año es moderado, mientras que la biomasa total acumulada (integral extendida) es de las más altas (Figuras 5 y 6). Este comportamiento se aprecia en particular en una zona bien conservada de oyamel, al norte de la RBMM, de orientación Norte-Oeste (Velasco et al. 2010, Champo et al. 2012).
Espacialmente, las variables fenológicas del oyamel muestran cierta dependencia con respecto a los efectos del relieve (Tabla 1), las zonas de bosque de oyamel más bajas y con mayor radiación,que se mantienen más cálidas en otoño-invierno al inicio de la temporada seca, por lo que termina más tarde la estación de crecimiento (el final de estación presenta una variación muy significativa con el relieve (p < 0.001 para el estadístico F), el cual explica el 11.6% de la variabilidad, su vegetación decae más lentamente al final de la temporada (14.2%), su LAI presenta el mínimo más alto (12.8%) con la menor amplitud (11.6%), es decir menor diferencia estacional en la cantidad de superficie foliar, lo que es congruente con el hecho de que el LAI acumulado en un ciclo estacional es menor (integral corta, 13.2%).
La cobertura de pino en la RBMM predomina en particular en las zonas centro y sur de la reserva, en orientaciones al Sur, donde hay un alto índice de radiación, en zonas menos elevadas y con mayores pendientes que el oyamel, donde las temperaturas son menos frías, la temporada de secas es más intensa, pero hay niveles ligeramente mayores de precipitación en la temporada de lluvias, si bien hay más variabilidad espacial. El bosque de pino incluye asociaciones de Pinus (de naturaleza perennifolia) con diferentes especies de Quercus (Q. castanea, Q. crassifolia, Q. laurina, Q. obtusata y Q. rugosa) (Cornejo-Tenorio et al. 2003), principalmente caducifolias (Romero-Rangel et al. 2014), además de una amplia diversidad florística, en las superficies abiertas donde se desarrollan estratos inferiores que denotan perturbación (Cornejo-Tenorio et al. 2003, Giménez de Azcárate et al. 2003). Esta cobertura requiere de menor humedad en la época de secas que el bosque de oyamel, soportando un intervalo más amplio de humedad y temperatura. La presencia de elementos latifoliados puede explicar la gran variación espacial de todas las variables fenológicas (Figuras 5 y 6). Las hierbas, arbustos y árboles tienen diferentes comportamientos fenológicos, que reflejan diferentes respuestas a factores ambientales, en particular a la estacionalidad de las lluvias; además, las plantas perennes tienen estructuras de reserva de nutrientes que les permiten empezar su actividad reproductiva antes que las plantas anuales (Cornejo-Tenorio e Ibarra-Manríquez 2007).
Las superficies más elevadas inician la estación de crecimiento antes (el efecto del relieve explica el 11.3% de la variabilidad) (Tabla 1), pero son las superficies más iluminadas las que presentan estaciones más largas (14.4%), con arranque (14.7%) y decaimiento (7.4%) más lentos, mínimos (14.2%) y máximos (6.4%) de LAI más altos, amplitudes (14.7%) de LAI más pequeñas, menos área foliar acumulada en un ciclo (8%) y más densidad de vegetación (14.9%). La temperatura es un factor clave del reinicio de la actividad de los árboles en primavera, si bien su efecto específico varía entre los diferentes órganos; sin embargo, en el cese de la actividad de los árboles en otoño no se identifica un factor clave, si bien la temperatura, el fotoperiodo y el estrés hídrico parecen ser los factores dominantes (Delpierre et al. 2016). Al respecto Carlón-Allende et al. (2015) identifican intervalos altitudinales de distribución de los bosques de oyamel (2 320 y 3 640 m) y los bosques de pino-encino (2 020 y 3 400 m) en laderas con pendiente de terreno mayor a 20◦ y alta insolación, donde los árboles muestran mayor sensibilidad a las variaciones climáticas.
Los cultivos en la RBMM se encuentran generalmente en ambientes más cálidos y húmedos, con elevaciones menores y pendientes moderadas, con orientaciones predominantes Norte y Oeste. Su desarrollo vegetativo inicia en el comienzo de la estación de lluvias, por lo que el inicio de estación se produce más tarde que en las otras coberturas; sin embargo, el crecimiento de las plantas, es más rápido y vigoroso, por lo que llegan antes a la mitad de estación; el decaimiento de la vegetación también es rápido, si bien el final de estación se da en un intervalo de tiempo mayor a partir del cese de las lluvias. Esta marcada estacionalidad se refleja en la climatología (Figura 1), con un LAI mínimo muy bajo (puede ser casi nulo) y una gran amplitud, si bien el LAI máximo no llega a ser tan alto como el de los bosques; esto explica valores bajos de LAI acumulado, tanto en un ciclo anual como de manera permanente (Figuras 5 y 6).
El comportamiento fenológico de los cultivos está relacionado con el relieve (Tabla 1), en particular el inicio de estación se produce antes en las zonas más elevadas de esta cobertura (21.6%), que mantienen la humedad y no dependen tanto del inicio de las lluvias; la estación de crecimiento es un poco más larga en las zonas de mayor radiación solar (14%); pero el decaimiento de la vegetación es más rápido en las zonas más elevadas y frías, y con menor iluminación (29%). Las zonas más bajas y con mayor irradiación presentan un mínimo de LAI más alto (22.9%), lo que puede explicarse por la existencia de vegetación remanente, que puede deberse a la presencia tanto de cultivos perennes, como de vegetación baja (ciertos frutales mantienen las hojas, si bien muchos son caducifolios) o bien de vegetación arbórea en los márgenes de los cultivos o en superficies adyacentes entre estos. Por otra parte, en las superficies más elevadas de esta cobertura, la amplitud del LAI es mayor, probablemente debido a que los cultivos son de temporal (por ejemplo, el maíz que puede alcanzar altas densidades de hojas cuando está en pleno desarrollo, pero queda completamente seco después de la cosecha).
La zona de arbustos incluye diferentes grupos de plantas no arbóreas, como pastizales, arbustos y herbáceas. En la RBMM, esta cobertura se distribuye en un intervalo amplio de elevaciones y pendientes (Figuras 1, 2, 3 y 4), en los claros de los bosques de oyamel y pino, formando manchones de vegetación secundaria resultantes de procesos de deforestación, en los que se pueden encontrar especies asociadas a procesos de sucesión de la vegetación, como Juniperus monticola Martínez (Giménez de Azcárate et al. 2003). Por su naturaleza, la vegetación arbustiva presenta un componente herbáceo senescente en la época de secas, germinando la nueva semilla al inicio del siguiente periodo de lluvias; la presencia, en algunos casos, de un componente perennifolio da lugar a una amplia gama de comportamientos fenológicos, que se reflejan en una estacionalidad más marcada que la de los bosques de oyamel y pino. El inicio de estación es la variable fenológica que presenta la mayor dependencia del relieve, siendo inversamente proporcional a la elevación: a mayor elevación, más se adelanta la estación de crecimiento de las plantas (40.7%), así como la fecha de máxima densidad de vegetación (19.3%) y de final de estación (14.6%).
Hay incertidumbre en los algoritmos de estimación del inicio de estación a partir de series de tiempo, ya que éstas no necesariamente reflejan las observaciones de campo, por un efecto de punto/pixel: las especies monitoreadas no representan el total del pixel. Para estudiar la fenología de un tipo de vegetación se necesitarían pixeles completamente homogéneos y para los que se tuviera la certeza de que no han cambiado de cobertura en el periodo considerado; así, los datos de variabilidad temporal se explicarían por elementos ambientales. En el caso de la RBMM, se sabe que se dan fenómenos de tala, ilegal o autorizada, que cambian la cobertura de pino u oyamel a arbustos (Leverkus et al. 2017). Otro problema es el tipo de observaciones (si se reporta la aparición de la primera hoja o de la primera flor) por lo que se recomienda observar la salida de las hojas (Delpierre et al. 2016). En todo caso, TIMESAT es consistentemente mejor que otros métodos, si bien tiende a estimar inicios de estación tempranos (White et al. 2009). Por otra parte, el final de la estación de crecimiento de las plantas, que se produce con la llegada del otoño y el cese de las lluvias, es un proceso difícil de aprehender, incluso en campo, porque la ausencia es más difícil de observar que la presencia; sin embargo, su estudio es de suma importancia, ya que muchas especies migratorias que se desplazan en otoño, como la mariposa monarca, están en declive por la pérdida de hábitat de cría, hibernación o parada, o porque se altera la abundancia o calidad nutricional de sus alimentos, o por interacciones competitivas con otras especies, a consecuencia de cambios fenológicos otoñales (Gallinat et al. 2015).
El estudio de la fenología a partir de variables obtenidas de series de LAI es una forma objetiva y sistemática de abordar el conocimiento de los ecosistemas de zonas que pueden ser amplias o inaccesibles, aún en ausencia de datos de campo, durante largos periodos de tiempo. Se considera que para realizar estudios fenológicos de larga duración se debe contar con datos de al menos 12 años (Cornejo-Tenorio e Ibarra-Manríquez 2007), lo que ya es posible con las series satelitales para cualquier lugar del mundo, pues se remontan al menos 20 años. Esto permite el monitoreo de ecosistemas, el análisis comparativo entre diferentes coberturas o ubicaciones (Verger et al. 2016), así como entre el estado base de un ecosistema y su condición en un momento dado, para conocer su evolución a lo largo del tiempo como efecto del cambio climático o la actividad antropogénica.
Conclusiones
Con la serie de tiempo GEOV1 de LAI se estudió la fenología de las coberturas vegetales presentes en la RBMM y su distribución espacial, en relación con el clima y el relieve. La variabilidad de respuesta mostrada por las diferentes coberturas es el resultado de la acción conjunta entre: a) las variables ambientales de elevación, temperatura, humedad e irradiación solar, asociadas al relieve; b) la tolerancia diferencial a los factores ambientales, característica de los diferentes grupos de plantas; y c) el establecimiento de una variedad de arreglos en mosaico de la vegetación, en los que hay proporciones variables, cualitativas y cuantitativas, de los elementos vegetales principales que los componen. Se trata de un estudio de Land Surface Phenology, sin observaciones de campo, lo que puede introducir imprecisiones en las fechas de inicio y final de estación. El tamaño de pixel de 1 km obliga a generalizar, tanto en las características abióticas como en los tipos de vegetación y su presencia relativa. La información generada permite conocer el comportamiento fenológico general de los principales tipos de vegetación en la RBMM y puede constituir la línea base para estudiar su evolución reciente, en relación con factores humanos y el cambio climático.

