











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
En las últimas décadas, el sureste de México ha sufrido severos procesos de degradación, particularmente provocado por el cambio de uso de suelo y deforestación (Ramos et al. 2004). La perturbación de hábitats naturales por acciones antropogénicas es un fenómeno global (Achard et al. 2002), causado por la agricultura (Harvey et al. 2006) y la urbanización (Husté y Boulinier 2007). Al respecto se sabe que las zonas periurbanas son un espacio limítrofe que constituyen un territorio de interfase, entre dos tipos de espacios geográficamente oposicionales y diferenciados entre el campo y la ciudad (Fernández y De la Vega 2017). En estas zonas se realizan actividades agrícolas, cría de ganado y se encuentran pequeños parches de vegetación, expuestos a la presión urbana (Ávila 2009).
Los recursos forestales de las áreas periurbanas son una opción que permite la generación de alimentos, bioenergía y la reducción de los gases efecto invernadero (Civeira 2015). Siendo los ecosistemas de bosques los que destacan por su capacidad de fijar carbono (C) en sus estructuras, de manera particular en la parte leñosa (Sullivan et al. 2017). Según Avendaño et al. (2009) el fuste de los árboles almacena aproximadamente el 84% de biomasa, de la que el 46% es C. Debido a lo anterior, el objetivo del presente trabajo fue evaluar la composición florística, y la captura de C en los parches de vegetación arbórea del área periurbana de Ocuiltzapotlan-Macultepec.
Materiales y métodos
Área de estudio
El trabajo se realizó en el área periurbana Ocuiltzapotlan-Macultepec Centro, Tabasco, ubicada entre las coordenadas 18o 07’ 7.75” y 18o 09’ 39” LN y 92o 52’ 24” y 92o 50’ 55” LO. El clima es cálido húmedo con abundantes lluvias en verano, temperatura media anual de 27 oC, y precipitación de 1 926 mm (INEGI 2017). La diversidad de suelos del área incluye Antrosoles, Fluvisoles, Gleysoles, Luvisoles, Lixisoles, Cambisoles, y en particular Acrisoles; todos ellos sobre un paisaje geomorfológico de terrazas costeras, formadas sobre rocas detríticas, con extensas áreas de lomeríos, pendiente suave, y buen drenaje superficial, con zonas de valles acumulativos y depresiones receptoras de sedimentos fluviodeluviales en la época de lluvias (Zavala-Cruz et al. 2016).
Muestreo de la vegetación
Mediante recorridos de campo se identificaron cuatro grupos de vegetación en el área de estudio: parches de selva media subperennifolia (SM), Plantación forestal (PF), Frutales (FR) y árboles en potrero (AP). En cada uno de los parches se establecieron parcelas de muestreo de 1000 m2 (20 x 50 m) en las que se realizó un inventario, para determinar las especies arbóreas; diámetro normal a 1.3 m del suelo con una cinta diamétrica, y la altura con una regla de Christensen (Christensen et al. 1996). Con la ayuda de los propietarios de los parches se registraron los nombres comunes de las especies arbóreas, y con literatura especializada y el apoyo de expertos, se identificaron los nombres científicos y se realizó la clasificación taxonómica a nivel de familia, género y especie.
Análisis de diversidad, uniformidad y similitud
La diversidad de especies arbóreas en las diferentes formaciones vegetales, se calculó con el índice de Shannon-Wiener (Magurran 2004).
H'= -∑si=1pilnpiDónde: pi es la proporción (o abundancia relativa) de cada especie en la población.
También, se evaluó la uniformidad o equidad (E) de las especies, mediante la ecuación E = H’/ln (S). Dónde: H’ es el índice de Shannon-Wienner, y S el número total de especies. El valor de E se sitúa entre 0 y 1 en los que valores cercanos a 1 representan condiciones de igualdad en la abundancia entre las especies de una población, mientras que valores cercanos a 0 indican la predominancia de una sola especie. La semejanza florística se calculó con el Coeficiente de Sorensen: Si = (2C / (A + B)), que se basa en la relación presencia-ausencia de especies entre dos sistemas. Dónde: A es el número de especies encontradas en la comunidad A, B el número de especies encontradas en la comunidad B, y C el número de especies comunes en ambos sistemas, es decir que compara el porcentaje de especies que se comparten entre dos poblaciones determinadas.
Análisis estructural
Para conocer la estructura, se calculó la densidad, dominancia y frecuencia por especie, el Índice de Valor de Importancia (IVI), que se calculó con la Ecuación propuesta por Stiling (1999).
IVI= DR+DOR+FRDónde: DR=Densidad relativa; DOR = dominancia relativa y FR = Frecuencia relativa.
La estructura vertical se determinó con los siguientes estratos de formación vegetal: bajo (0-10 m), medio (11-20 m) y alto (> 21), mientras que la estructura horizontal contempló seis clases diamétricas con ancho de 10 cm, como lo establecen López- Pérez et al. (2014).
Carbono en la biomasa aérea
Para cuantificar el carbono (C) fijado en la biomasa arbórea de cada formación vegetal se utilizó la ecuación propuesta por Chave et al. (2014).
AGBest=0.0673 X (pD2H)0.976Dónde: D es el diámetro normal en cm, H es la altura total en m, y p es la densidad de la madera en g cm-3.
Los valores de densidad de la madera se obtuvieron de la Global Wood Density Database (Zanne et al. 2009). Para el cálculo del contenido de C en la biomasa arbórea, se empleó la fórmula propuesta por el IPCC (2003).
C=B*FCDónde: C es el carbono en la biomasa arbórea, B: la biomasa arbórea en kg, y FC: la fracción de C (0.5).
Análisis de los datos
El contenido de carbono se analizó mediante estadística no paramétrica, con la prueba de Kruskal Wallis a un nivel de significancia de p < 0.05 (Siegel y Castellan 1995), mientras que los análisis se realizaron con el programa estadístico Statgraphics® centurión XVI.
Resultados y discusión
Los 402 individuos que se encuentran en los relictos de vegetación del área periurbana Ocuilzapotlán-Macultepec se agruparon en 21 familias, 35 géneros y 38 especies, el 62% de las familias estuvieron representadas por un género, 24% por dos géneros, y el 14% restante por más de 2 géneros. Las familias más representadas fueron las Fabáceas (cinco géneros y cinco especies), Bignoniáceas (cuatro géneros y cuatro especies) y Malváceas (tres géneros y tres especies). Las Meliácea, Palmae, Sapindácea y Sapotáceo tu vieron dos géneros y dos especies cada una, en tanto que la familia Rutácea tuvo dos géneros y cinco especies. Del total de individuos registrados, el 55% se agruparon en cuatro familias: Fabácea (18%), Apocynácea (15%), Malvácea (12%) y Anacardiácea (10%). Las especies más frecuentes fueron: Haema- toxylum campechianum (63 individuos), Tabernae- montana alba (60 individuos), Mangifera indica (41 individuos), Guazuma ulmilifolia (32 individuos), Coccoloba barbadensi y Tabebuia rosea (22 individuos cada una), y Rinorea guatemalensis (20 individuos). Los estudios sobre espacio periurbano en el estado de Tabasco son escasos, lo que hace difícil contrastar los hallazgos bajo el enfoque de la riqueza florística del espacio periurbano. Al respecto, Zavala-Cruz et al. (2016) señalan que la vegetación natural arbórea periurbana de la ciudad de Villahermosa, Tabasco, se distribuye en relictos de selva mediana subperennifolia de Brosimum alicastrum en los lomeríos, Haematoxylum campechianum en las llanuras de inundación y valles, así como vegetación secundaria de Cecropia obtisifolia, Heliocarpus donnell-Smithii y Guazuma ulmifolia en la planicie fluviodeltaica, con excepción de Brosimum alicastrum, todas estas especies se encontraron en el presente estudio. Mientras que Bodin et al. (2006), señalan que la importancia de conservar las áreas periurbanas, radica en su riqueza florística, más allá de su extensión, ya que actualmente el establecimiento de zonas prioritarias para la conservación a escalas estatales o nacionales, se hace solo considerando áreas extensas e ignorando pequeñas áreas producidas por fragmentación del hábitat, aunque estas pueden contener diversidad.
El índice de diversidad de Shannon-Wienner (H’) indica que los FR registraron la mayor diversidad de especies (H’ = 2.8), seguido de los parches de SM (H’ = 1.9), y AP (H’ = 1.0). Con base en el valor de equidad de especies (E), el número de especies es más equitativo en los FR con E de 0.84, seguido de la SM y AP con valores de E de 0.70 y 0.53, respectivamente. Las pruebas pareadas para determinar el coeficiente de similitud florística de Sorenson (Si), indican que la SM y AP, son similares en un 17% con respecto a las especies presentes en las dos poblaciones (St = 0.17), entre SM y FR del 36%, mientras que entre AP y FR fue del 34%. Los valores obtenidos son menores que los reportados para áreas no perturbadas, como lo demuestran Carreon-Santos y Valdez-Hernández (2014) quienes reportaron valores de H’ = 3.3 en selva mediana sub- perenifolia, H’ = 3.3 a 3.9 para selva mediana peren- nifolia (Vázquez-Negrín et al. 2011); H = 3.4 en selva alta perennifolia y en selva mediana subcaducifolia de H’ = 3.6 a 4.8 (Maldonado-Sánchez y Maldonado- Mares 2010). Por lo que los índices de diversidad muestran que la SM ha sido severamente intervenida, debido a que los pobladores extraen la madera para construcción de casas, leña, estacas para alambrado y otros usos domésticos. Para las formaciones vegetales a base de frutales (FR) se observó una mayor diversidad de especies como mango, cítricos, zapotes y chicozapotes, además de árboles maderables como cedro y caoba, en estas formaciones vegetales el manejo es fundamental para conservar dichas especies para comercializar los frutos o autoconsumo y por otro lado la madera se usa para muebles o venta. La riqueza de especies presente en los parches de vegetación en los potreros es más limitada, ya que solo dejan las plantas de interés, razón por la cual resalta la especie Haematoxylum campechianum, debido a su uso para reparar las cerca de alambrado que dividen los predios. En la Tabla 1, se muestran las especies que tuvieron los mayores IVI por formación vegetal, el valor más alto fue de 56.03 para la Ceiba pentandra localizada en la SM, debido a su mayor área basal, le siguen en orden de importancia, Tabernaemontana alba (48.58) y Guazuma ulmifolia (32.69). En árboles de potreros (AP), los IVI más altos lo tuvieron las especies Haematoxylum campechianum, Coccoloba barbadensi y Tabebuia rosea con valores de 127.09, 77.39 y 43.02, respectivamente; debido a su mayor abundancia y área basal; las tres especies son comunes en praderas de inundación. Mientras que los parches de vegetación correspondiente a frutales (FR) registraron IVI de 45.39, 24.12 y 19.82 para las especies Mangifera indica, Manilkara Zapota y Spathodea campanulata, respectivamente, debido a su frecuencia y área basal.
Tabla 1: Especies con mayor índice de valor de importancia por cada formación vegetal.
| Familia | Especie | Densidad relativa | Frecuencia relativa | Dominancia Relativa | IVI |
| SM | |||||
| Euphorbiaceae | Apium pedicellatum | 5.36 | 6.67 | 12.19 | 24.21 |
| Malvaceae | Ceiba pentandra | 3.57 | 6.67 | 45.80 | 56.03 |
| Malvaceae | Guazuma ulmifolia | 11.90 | 10.00 | 10.79 | 32.69 |
| Anacardiaceae | Mangifera indica | 1.79 | 3.33 | 15.30 | 20.42 |
| Violaceae | Rinorea guatemalensis | 11.90 | 10.00 | 3.86 | 25.76 |
| Apocynaceae | Tabernaemontana alba | 35.71 | 10.00 | 2.86 | 48.58 |
| 10 especies más | 29.76 | 53.33 | 9.20 | 92.30 | |
| Total | 100 | 100 | 100 | 300 | |
| AP | |||||
| Polygonaceae | Coccoloba barbadensi | 19.81 | 23.08 | 34.50 | 77.39 |
| Fabaceae | Haematoxylum campechianum | 59.43 | 23.08 | 44.58 | 127.09 |
| Bignoniaceae | Tabebuia rosea | 6.60 | 23.08 | 13.34 | 43.02 |
| 4 especies más | 14.15 | 30.77 | 7.58 | 52.50 | |
| Total | 100 | 100 | 100 | 300 | |
| FR | |||||
| Urticaceae | Cecropia obtusifolia | 11.15 | 2.33 | 2.12 | 15.60 |
| Anacardiaceae | Mangifera indica | 11.15 | 9.30 | 24.94 | 45.39 |
| Sapotaceae | Manilkara Zapota | 6.69 | 2.33 | 15.11 | 24.12 |
| Bignoniaceae | Spathodea campanulata | 5.58 | 2.33 | 11.92 | 19.82 |
| Bignoniaceae | Tabebuia rosea | 3.72 | 6.98 | 7.20 | 17.89 |
| Lamiaceae | Vitex gaumeri | 10.04 | 4.65 | 1.81 | 16.50 |
| 22 especies más | 52.68 | 72.10 | 36.89 | 162.68 | |
| Total | 100 | 100 | 100 | 300 |
SM = selva mediana, AP = árboles en potrero, FR = frutales
La altura promedio de los árboles fue de 8.5 m para SM con rango de 2 a 30 m, para los FR fue de 12.9 m, con rango de 3 a 36 m y la formación vegetal AP fue de 7.4 m con rango de 3 a 34 m. La mayoría de las especies arbóreas se encuentran entre 0 y 10 m de altura, por lo que en FR se encontró el 53%, para el SM 74% y 91% para AP. En el estrato medio de 11 a 20 m de altura, los porcentajes encontrados fueron de 30, 22 y 9% para FR, SM y AP, respectivamente; mientras que, en el estrato alto mayor de 20 m de altura, los FR y SM representaron solo el 17 y 4%, respectivamente (Figura 1). Estos resultados indican diferentes estados de sucesión de los parches de vegetación, debido a la extracción de madera en SM, a la preservación selectiva de las especies arbóreas de interés en FR y AP. Las tendencias observadas son similares a las reportadas por otros autores en selvas tropicales como López-Pérez et al. (2014) quienes reportan que en el estrado bajo se acumula el 80% de las especies, en el medio el 13% y en el estrato alto entre 4 y 9%. Mientras que Zamora-Crescencio et al. (2017) indican que el mayor número de individuos se concentran en el primer estrato, disminuyendo según incrementa la altura.
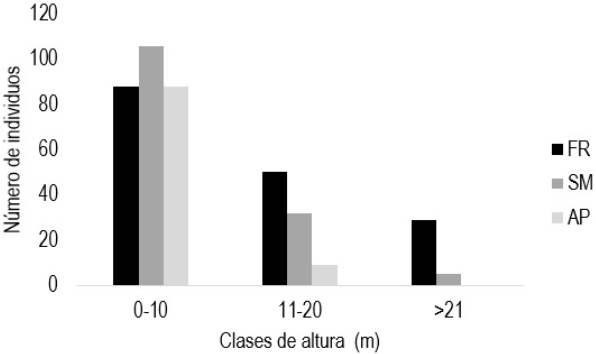
Figura 1: Distribución vertical del componente arbóreo en las tres formaciones vegetales. SM = selva mediana, AP = árboles en potrero, FR = frutales.
El diámetro promedio de los árboles en SM fue de 17.3 con rango de 5.1 a 160.7 cm, en FR de 21 con rango de 5.1 a 65.3 cm y en AP de 20 cm con rango de 5.5 a 66.8 cm. Para la distribución de las clases diamétricas, la mayor proporción de individuos se concentraron en la clase de 0 a 10 cm para FR y SM con 33 y 56%, respectivamente. Para AP la mayor proporción (36%) lo presentó la clase de 11 a 20 cm, en la clase de diámetros mayores de 50 cm se encontraron 4, 3 y 4% para FR, SM y AP, respectivamente (Figura 2). Se encontró que la mayor cantidad de individuos son menores a los 10 cm de diámetros, resultados similares son reportados por Maldonado-Sánchez y Maldonado-Mares (2010), quienes indican que ente el 64 y 76%, corresponden a individuos cuyas clases diamétricas son las más bajas (< 10 cm), comportamiento que se encontró en el presente estudio.
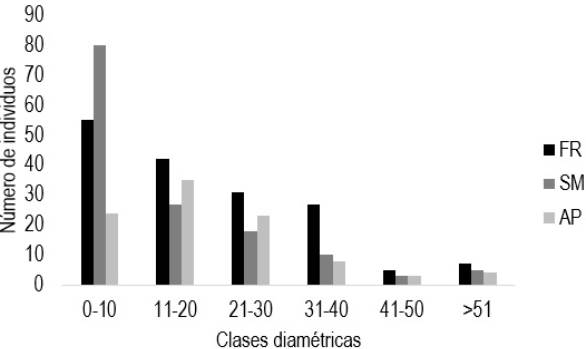
Figura 2: Distribución diamétrica del componente arbóreo en las tres formaciones vegetales. SM = selva mediana, AP = árboles en potrero, FR = frutales.
La densidad de árboles por tipo de vegetación fue de 477 árboles ha-1 para SM, 288 árboles ha-1 para FR y de 323 árboles ha-1 para AP. Los mayores valores de área basal (AB) se encontraron en SM (28.14 m2 ha-1) debido a los arboles de ceiba que tuvieron diámetros mayores a 51 cm, en FR fue de 20.75 m2 ha-1, mientras que en AP fue de 14.85 m2 ha-1. Al respecto Honorio-Coronado et al. (2015) en bosques aluviales, reportan valores de AB similares a los encontrados en el presente estudio, pero la acumulación de C en la biomasa superior a la estimada en la presente investigación, llegando a acumular hasta un 60% más C, con respecto a SM, FR y AP del área periurbana Ocuiltzapotlán-Macultepec. La mayor acumulación de C, se atribuye a variables como la altura y la densidad de la madera, lo que concuerda con los resultados encontrados, ya que aunque el área basal es similar entre ambos trabajos, las alturas del arbolado periurbano en la SM, FR y AP no rebasan los 10 m. En el polígono del área periurbana Ocuiltzapotlán-Macultepec son pocas las especies con diámetros grandes debido al manejo de las plantaciones y saqueo de los relictos de selvas que aún pueden encontrarse. El C estimado en la biomasa arbórea mostró diferencias significativas entre las formaciones vegetales, para SM registró 84.82 Mg C ha-1, seguido de FR con 80.94, PF con 48.42 Mg C ha-1, y AP con 38.99 Mg C ha-1, esta capacidad de captura de carbono aérea está influenciada por factores como la composición florística, la riqueza de especies, el área basal y el manejo proporcionado a las formaciones vegetales. Al respecto Aguirre- Calderón y Jiménez-Pérez (2011) reportan que el carbono secuestro en los ecosistemas forestales en forma de biomasa aérea, varía en función de la composición florística, la edad y la densidad de población de cada estrato. Mientras que Honorio-Coronado et al. (2015) reportan que los bosques aluviales capturan en promedio 193.9 Mg C ha-1, pero se ha documentado que estos bosques pueden disminuir su capacidad de almacenamiento hasta alcanzar tasas de almacenamiento de tan solo 72.9 Mg C ha-1 debido al uso irracional de los recursos naturales, y su transformación en áreas agropecuarias.
El área periurbana Ocuiltzapotlán-Macultepec tiene menor diversidad de especies que áreas no perturbadas, pero a pesar de la reducción provocada por la intervención antropogénica, tiene una importante diversidad de especies, particularmente de frutales, selva mediana subperennifolia y árboles en potreros. Con base en el valor de equidad de especies, el número de especies es más equitativo en los FR, seguido de SM y AP. La altura y el diámetro de las especies le confieren una estructura de la vegetación arbórea, suficiente para albergar una biomasa que llega a acumular 84.82 Mg C ha-1 en SM, seguido de FR, PF y AP con valores de 80.94, 48.42 y 38.99 Mg C ha-1, respectivamente. La captura de carbono es una de las principales funciones ambientales del área periurbana evaluada.

