











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
A nivel mundial se tiene un registro de 9 271 especies de aves (Navarro-Sigüenza et al. 2014). Pero a pesar de esta gran diversidad de especies las tazas de extinción han incrementado de forma drástica debido a la destrucción y alteración de su hábitat, que altera los procesos ecológicos positivos y negativos, como la segregación de nichos y la com petencia por nuevos espacios (Birdlife International 2008). Durante la obtención del alimento, las aves emplean diferentes técnicas para la obtención del recurso, basado en gremios tróficos y competencia por alimento (Torrens et al. 2017). La teoría del forrajeo óptimo menciona que las aves utilizan técnicas de cacería que requieren menor gasto energético para obtener su alimento, con mayor aporte nutricional (Ginnobili y Roffe 2016, Cruz-Miranda et al. 2017). De igual forma el comportamiento de los organismos se define por la selección natural, misma que se integra por eventos y coacciones que determinan la supervivencia de las especies, y determinan el éxito reproductivo (Frere et al. 2005). La hipótesis del gasto de crianza indica que las aves residentes, en época reproductiva están limitadas por la presencia de pocos artrópodos de talla grande, mismas que capturan para optimizar su adecuación reproductiva, mientras que en periodo no reproduc tivo se tiene abundancia de presas de menor tamaño que ayuda a los organismos adultos a sobrevivir (Johnson et al. 2005, Fandiño et al. 2016). A partir de diversas estrategias las aves demuestran una utilización del hábitat que les permite establecer interacciones positivas (Velasco etal. 2017, Seeholzer et al. 2017). Un aspecto importante que determina la coexistencia de aves insectívoras es la distribución de sus presas, lo cual puede determinar la ecología de las aves insectívoras (De la Ossa et al. 2017).
En México se registran 1 107 especies de aves, de las cuales 102 son endémicas (Berlanga et al. 2015), que se distribuyen desde zonas tropicales hasta semiáridas, encontrándose algunas en alguna categoría de riesgo (SEMARNAT 2010). Debido a una amplia gama de factores de perturbación y disturbio (Navarro-Sigüenza et al. 2014), lo que ha provocado diversos procesos de segregación por es pacio, alimento y dominio de nuevos nichos ecológicos (Pineda-Pérez et al. 2014). Por ello las aves insectívoras tienen fluctuaciones, debido a la estacionalidad de insectos, viéndose obligadas a recurrir a las siguientes acciones: cambiar su dieta, consumir insectos inactivos o abandonar el área y buscar un nuevo nicho alimenticio (Ortiz-Pulido et al. 2016).
Una zona con un rodal forestal relevante para aves insectívoras en bosque templado de pino-encino, se localiza en la Cuenca San Bernardino, en la parte central del Eje Neo volcánico transversal; sujeta a perturbación como la tala inmoderada, urbanización, cacería furtiva, tráfico ilegal y el cambio de uso de suelo (Moreno 2007). En general, las aves juegan un papel importante en el control biológico y equilibrio del ambiente, por lo que se consideran bioindicadoras de perturbación (Navarro-Sigüenza et al. 2014); forman parte de cadenas tróficas, y son dispersoras de semillas y polinizadoras (Aguilar y Tinoco 2017). A pesar de su importancia ecológica, en México existen pocos estudios sobre ecología trófica de aves (Ugalde-Lezama et al. 2011 y Pineda-Pérez et al. 2014), que reportan la determinación de dietas de aves insectívoras de selvas y bosques templados. Por lo anterior, el objetivo del presente trabajo fue determinar de forma comparativa las dietas y coexistencia trófica en la estructura de aves insectívoras de un bosque de pino-encino en tres elevaciones de la Cuenca de San Bernardino, Texcoco, Estado de México.
Materiales y métodos
Área de estudio
La Subcuenca del río San Bernardino se ubica en el municipio de Texcoco de Mora, Estado de México. En la parte este del monte Tláloc, a una altitud de entre 2 600 y 3 000 msnm, en las coordenadas 98°47’ 44.19 y 98° 45’ 20.85 LN y 19° 28’ 0.91 y 19°26’ 29.68 LO. Se evaluaron las elevaciones altitudinales (EA) de : 2 600 msnm (EA2600), 2 800 msnm (EA2800) y 3 000 msnm (EA3000) considerando una superficie de 21 ha para cada elevación. En cada EA se usó muestreo sistemático (MS) con distancias predeterminadas de 200 m en arreglos a manera de cuadriculas, con tamaño de muestra de ocho unidades de elección (UEL) con una superficie de 50 m2 cada uno y 200 m de distancia entre cada punto (Gallina y López-González 2011). Cada 16 d se realizó el seguimiento de las aves de octubre de 2016 a enero de 2017, aplicando un total de 5 muéstreos. El monitoreo se realizó en función del muestreo de recuento en puntos con radio fijo y búsqueda intensiva. Este esquema consistió en que el observador al llegar a cada punto de muestreo, se mantenía quieto por un minuto y luego recorría un radio de 25 m en búsqueda de ejemplares, utilizando binoculares de 8 x 42 m de resolución marca Bushnell. Únicamente se registraron individuos que se encontraban alimentando o aplicando alguna técnica de cacería, de las 07:00 a 14:00 h, periodo en que las aves presentan mayor actividad forrajera.
El monitoreo de insectos se realizó con el método de corte y sacudida de ramas, que consiste en ubicar a las aves alimentándose. Para luego colocar una bolsa de plástico en la rama en la que se encontraba el ave y sacudir con el fin de obtener los insectos de los que potencialmente se pudiera estar alimentando. La identificación de los insectos se realizó con claves taxonómicas propuestas por Borror et al (1989), Equihua (1989) y Stehr (1991). Los insectos se fotografiaron con una cámara digital marca Pax Cam 3, con óptica Kern Macro-Switar, modelo profesional para equipos de microscopía electrónica de barrido marca JEOL 35- C y se medieron con el analizador de imágenes Image Tool para Windows versión 3.0. Para la obtención de excretas se llevó a cabo la contención física de aves mediante el método de captura con redes de niebla mediante esfuerzo constante (Villaseñor y Santana 2002). Las redes utilizadas tuvieron un tamaño de 12 m de largo por 2.5 m de ancho y luz de malla de 36 mm de diámetro, se colacaron de las 07:00 a 16:00 h. Las aves capturadas se colocaron en cajas de cartón por una hora con la finalidad de obtener sus heces, las cuales se colocaron en viales con alcohol al 70% y se trasladaron al laboratorio de entomología de la Universidad Autónoma Chapingo. Las muestras se analizaron con la técnica propuesta por Whitaker (1988) con un microscopio estereoscopio con resolución de 10 a 80 x. Los fragmentos encontrados se identificaron con las claves taxonómicas antes señaladas; mientras que la identificación de las aves se realizó con las guías de identificación estándar (Peterson y Chalif 1989, National Geographic 2002).
Para determinar la posible asociación entre las aves e insectos derivados de la técnica de corte y sacudida de ramas, las técnicas de cacería y los insectos consumidos; se aplicaron análisis de regresión de poisson (ARP), con el procedimiento de selección de variables polinómica por pasos hacia atrás (Stepwise), el ajuste de los modelos se realizó con el criterio del mínimo Akaike en R.13.0 (Akaike 1969). Para establecer posibles diferencias estadísticas en los datos registrados en las diferentes elevaciones, e inferir si los datos registrados son los potencialmente existentes, se aplicaron análisis de x2 y Krustal Wallis, en virtud de no cumplirse los supuestos de la estadística paramétrica, los análisis se realizaron con el programa JMP IN versión 8.0. Para visualizar de forma gráfica la posible relación entre las aves y las variables evaluadas, se realizaron análisis de correspondencia simple (ACS), con el programa Statistica™ versión 13.0.
Resultados
Los resultados de la regresión de Poisson para los datos de corte y sacudida de ramas sugieren que siete familias de insectos presentan efecto sobre las abundancias de aves en las elevaciones consideradas, presentando un AIC de 65.691, 57.811, 106.08 y 87.63 (Tabla 1). Mientras que las técnicas de cacería sugieren que una técnica presenta efecto sobre la abundancia de aves en las elevaciones, con AIC de 28.735, 25.21, 32.823 y 67.65 (Tabla 2).
Tabla 1 Resultados de regresión Poisson para las abundancias de las familias de insectos registradas en cada elevación.
| Elevación altitudinal 2600 | |||||
| Coeficientes : | Estimados | Error Estándar | Valor de Z | Pr(>|Z|) | Significancia |
| (Intercepto) | -1.029 62 | 0.447 21 | -2.302 | 0.0213 16 | * |
| Anthocoridae | 0.105 62 | 0.014 23 | 7.421 | -11.831 | *** |
| Bostrichidae | 0.287 13 | 0.139 44 | 2.059 | 0.0394 84 | * |
| Cicadellidae | 0.377 01 | 0.090 35 | 4.173 | -1.891 | *** |
| Coccinellidae | 1.319 53 | 0.316 22 | 4.173 | -1.891 | *** |
| Geometridae | 0.483 18 | 0.134 16 | 3.601 | 0.000 316 | *** |
| Membracidae | 0.574 26 | 0.278 89 | 2.059 | 0.039 484 | * |
| Miridae | 0.603 98 | 0.167 7 | 3.601 | -0.831 | *** |
| No identificado | 0.201 68 | 0.034 86 | 5.785 | -1.751 | *** |
| Elevación altitudinal 2800 | |||||
| (Intercepto) | -2.397 9 | 1 | -2.398 | 0.016 49 | * |
| Anthocoridae | 2.397 9 | 1.414 21 | 1.696 | 7.009 | . |
| Cicadellidae | 0.400 73 | 0.109 54 | 3.658 | 0.000 254 | *** |
| Coccinellidae | 1.198 95 | 0.707 11 | 1.696 | 7.009 | . |
| Eulophidae | 2.397 9 | 1.414 21 | 1.696 | 7.009 | . |
| Gelechiidae | 0.799 3 | 0.471 4 | 1.696 | 0.089 968 | . |
| Largidae | 2.397 9 | 1.414 21 | 1.696 | 0.089 968 | . |
| Lumbricidae | 1.198 95 | 0.707 11 | 1.696 | 7.009 | . |
| Miridae | 1.335 78 | 0.365 15 | 3.658 | -1.451 | *** |
| No identificado | 0.353 47 | 0.081 08 | 4.359 | -3.691 | *** |
| Phoridae | 2.397 9 | 1.414 21 | 1.696 | 7.009 | . |
| Tephritidae | 2.397 9 | 1.414 21 | 1.696 | 7.009 | . |
| Elevación altitudinal 3000 | |||||
| (Intercept) | 0.699 | 1.369 | 0 | 1 | |
| Cicadellidae | 7.059 | 2.539 | 3.184 | -1.541 | ** |
| Geometridae | 2.309 | 1.049 | 10.832 | < 2E-16 | *** |
| Lauxaniidae | 2.30 | 2.949 | 5.838 | -3.721 | *** |
| No identificado | 0.819 | 0.339 | 7.746 | -5.531 | *** |
| Todas las elevaciones | |||||
| (Intercept) | 0.251 314 | 0.235 702 | 1.066 | 0.286 32 | |
| Anthocoridae | 0.065 686 | 0.008 022 | 8.188 | -13.331 | *** |
| Cicadellidae | 0.129 302 | 0.018 4 | 7.027 | -9.881 | *** |
| Coccinellidae | 0.385 111 | 0.117 851 | 3.268 | -1.911 | ** |
| Geometridae | 0.212 178 | 0.019 756 | 10.74 | < 2E-16 | *** |
| Lauxaniidae | 2.051 271 | 0.394 405 | 5.201 | -5.011 | *** |
| Miridae | 0.277 987 | 0.058 321 | 4.766 | -4.121 | *** |
| No identificado | 0.073 754 | 0.006 659 | 11.075 | < 2E-16 | *** |
| Tephritidae | 0.477 549 | 0.071 27 | 6.701 | -8.911 | *** |
Códigos de significancia 0 '***', 0.001 '**', 0.01'*', 0.05 '.', 0.1 ' '
Tabla 2: Resultados de regresión Poisson para las abundancias de aves y las técnicas de cacería exhibidas en cada elevación.
| Elevación altitudinal 2600 | |||||
| Coeficientes : | Estimados | Error Estándar | Valor de Z | Pr(>|Z|) | Significancia |
| (Intercepto) | 0.099 54 | 0.347 19 | 0.287 | 0.774 | |
| Colectar | 0.227 77 | 0.050 32 | 4.527 | 0.009 | *** |
| Elevación altitudinal 2800 | |||||
| (Intercepto) | -0.572 | 0.996 3 | -0.574 | 0.566 | |
| Remover | 0.490 8 | 0.287 6 | 1.706 | 6.809 | . |
| Elevación altitudinal 3000 | |||||
| (Intercepto) | 0.706 2 | 0.306 | 2.308 | 0.021 | |
| Colectar | 0.154 6 | 0.037 2 | 4.156 | -1.761 | *** |
| Elevaciones 2600-3000 | |||||
| (Intercepto) | 0.415 3 | 0.234 13 | 1.774 | 0.0761 | . |
| Colectar | 0.1857 6 | 0.029 41 | 6.315 | -7.291 | *** |
| Remover | 0.235 48 | 0.108 01 | 2.18 | 0.929 | * |
Códigos de significancia 0 '***', 0.001 '**', 0.01'*', 0.05 '.', 0.1 ' '
Para el consumo de insectos se observa que un orden de insectos tiene efecto sobre las abundancias de las aves con AIC de 25.159, 19.909, 18.755 y 33.503 (Tabla 3). Los datos de corte y sacudida de ramas derivados del ARP indican que no existen diferencias significativas entre elevaciones (Tabla 4). Para las técnicas de cacería colectar y remover, no se observaron diferencias significativas entre ellas (Tabla 5). Los insectos consumidos (excretas) Coleóptera e Hymenoptera, no presentan diferencias significativas entre elevaciones, por loque son estadísticamente similares. Los resultados de X2 para los datos de corte y sacudida de ramas derivados del ARP, sugieren que las familias de insectos registradas son las que potencialmente podrían presentarse en las elevaciones estudiadas, a excepción de Miridae (Tabla 6). Las técnicas de cacería colectar y remover son las que pueden ser exhibidas por las aves (Tabla 7), mientras que los insectos de los órdenes Coleóptera e Hymenoptera son los más consumidos por la avifauna.
Tabla 3: Resultados de Regresión Poisson para la abundancia de aves y los órdenes de insectos registrados en las excretas de cada elevación
| Elevación altitudinal 2600 | |||||
| Coeficientes : | Estimados | Error Estándar | Valor de Z | Pr(>|Z|) | Significancia |
| (Intercepto) | 0.470 004 | 0.353 553 | 1.329 | 0.184 | |
| Coleóptera | 0.041 604 | 0.006 553 | 6.348 | -7.821 | *** |
| Elevación altitudinal 2800 | |||||
| (Intercepto) | -0.223 144 | 0.499 999 | -0.446 | 0.655 | |
| Coleóptera | 0.048 649 | 0.008 637 | 5.633 | -6.211 | *** |
| Elevación altitudinal 3000 | |||||
| (Intercepto) | -1.386 294 | 0.999 932 | -1.386 | 0.1656 | |
| Coleóptera | 0.038 083 | 0.007 865 | 4.842 | -4.711 | *** |
| Himenóptera | 0.231 049 | 0.136 077 | 1.698 | 6.959 | . |
| Elevaciones 2600-3000 | |||||
| (Intercepto) | 1.098 612 | 0.258 199 | 4.255 | -2.901 | *** |
| Coleóptera | 0.012 521 | 0.001 106 | 11.32 | < 2e-16 | *** |
Códigos de significancia 0 '***', 0.001 '**', 0.01'*', 0.05 '.', 0.1 ' '
Tabla 4: Resultados de Kruskal-Wallis para las familias de insectos, derivadas del ARP, en las elevaciones 2600, 2800, 3000 en la cuenca de San Bernardino, Texcoco, Estado de México.
| Familias de insectos datos de presencias y ausencias | |||
| Familia | Ji cuadrado | Grados de libertad | Prob> Ji cuadrado |
| Anthocoridae | 3.5 | 1 | 0.061 4 |
| Bostrichidae | 3.230 8 | 1 | 0.072 3 |
| Cicadellidae | 0 | 1 | 1 |
| Coccinellidae | 0.807 7 | 1 | 0.368 8 |
| Geometridae | 0 | 1 | 1 |
| Membracidae | 3.230 8 | 1 | 0.072 3 |
| Miridae | 6.222 2 | 1 | 0.0126* |
| No identificado | 0.583 3 | 1 | 0.445 |
| Eulophidae | 0 | 1 | 1 |
| Gelechiidae | 0 | 1 | 1 |
| Largidae | 0 | 1 | 1 |
| Lumbricidae | 0 | 1 | 1 |
| Phoridae | 0 | 1 | 1 |
| Tephritidae | 0.477 3 | 1 | 0.489 7 |
| Lauxaniidae | 1.5 | 1 | 0.220 7 |
| Familias de insectos datos de frecuencias | |||
| Anthocoridae | 7.466 7 | 4 | 0.113 2 |
| Bostrichidae | 3.230 8 | 1 | 0.072 3 |
| Cicadellidae | 3.966 7 | 4 | 0.410 5 |
| Coccinélida | 1.507 7 | 2 | 0.470 6 |
| Geometridae | 4.666 7 | 4 | 0.323 2 |
| Membracidae | 3.230 8 | 1 | 0.072 3 |
| Miridae | 6.968 9 | 2 | 0.030 7* |
| No identificado | 4.9 | 5 | 0.428 2 |
| Eulophidae | 0 | 1 | 1 |
| Gelechiidae | 0 | 1 | 1 |
| Largidae | 0 | 1 | 1 |
| Lumbricidae | 0 | 1 | 1 |
| Phoridae | 0 | 1 | 1 |
| Tephritidae | 3.627 3 | 3 | 0.304 6 |
| Lauxaniidae | 1.5 | 1 | 0.220 7 |
Tabla 5:Resultados de Kruskal-Wallis para colectar y remover, Coleóptera e Hymenoptera derivadas del ARP, en las elevaciones 2600, 2800, 3000 de la cuenca de San Bernardino Texcoco Estado de México.
| Técnicas de cacería datos de presencia ausencia | |||
| Técnica | Ji cuadrada | Grados de libertad | Prob> Ji cuadrado |
| Colectar | 0.063 5 | 1 | 0.801 1 |
| Remover | 1.257 4 | 1 | 0.262 2 |
| Técnicas de cacería datos frecuencias | |||
| Colectar | 3.125 2 | 4 | 0.537 1 |
| Remover | 1.257 4 | 1 | 0.262 2 |
| Ordenes de insectos | |||
| Coleóptera | 8.871 6 | 6 | 0.180 9 |
| Hymenoptera | 0.530 3 | 1 | 0.466 5 |
Tabla 6: Análisis de X2 para las Familias de insectos probablemente colectadas por las aves durante su alimentación en las elevaciones bajo estudio.
| Familias de insectos datos de presencia ausencia | ||||
| Familia | N | Grados de libertad | Ji cuadrado | Prob> Ji Cuadrado |
| Anthocoridae | 15 | 2 | 5 | 0.082 1 |
| Bostrichidae | 15 | 2 | 4.615 | 0.099 5 |
| Cicadellidae | 15 | 2 | 0.536 | 0.765 |
| coccinellidae | 15 | 2 | 1.154 | 0.561 6 |
| Geometridae | 15 | 2 | 5 | 0.082 1 |
| Membracidae | 15 | 2 | 4.615 | 0.099 5 |
| Miridae | 15 | 2 | 6.667 | 0.357* |
| No identificado | 15 | 2 | 2.5 | 0.286 5 |
| Eulophidae | 15 | 2 | 2.143 | 0.342 5 |
| Gelechiidae | 15 | 2 | 2.143 | 0.342 5 |
| Largidae | 15 | 2 | 2.143 | 0.342 5 |
| Lumbricidae | 15 | 2 | 2.143 | 0.342 5 |
| Phoridae | 15 | 2 | 2.143 | 0.342 5 |
| Tephritidae | 15 | 2 | 0.682 | 0.711 1 |
| Lauxaniidae | 15 | 2 | 2.143 | 0.342 5 |
| Familias de insectos datos de frecuencias | ||||
| Anthocoridae | 15 | 8 | 10 | 0.265 |
| Bostrichidae | 15 | 2 | 4.615 | 0.099 5 |
| Cicadellidae | 15 | 8 | 7.25 | 0.509 9 |
| Coccinellidae | 15 | 4 | 4.154 | 0.385 6 |
| Geometridae | 15 | 8 | 10 | 0.265 |
| Membracidae | 15 | 2 | 4.615 | 0.099 5 |
| Miridae | 15 | 4 | 9.867 | 0.042 7* |
| No identificado | 15 | 10 | 12 | 0.285 1 |
| Eulophidae | 15 | 2 | 2.143 | 0.342 5 |
| Gelechiidae | 15 | 2 | 2.143 | 0.342 5 |
| Largidae | 15 | 2 | 2.143 | 0.342 5 |
| Lumbricidae | 15 | 2 | 2.143 | 0.342 5 |
| Phoridae | 15 | 2 | 2.143 | 0.342 5 |
| Tephritidae | 15 | 6 | 5.182 | 0.520 7 |
| Lauxaniidae | 15 | 2 | 2.143 | 0.342 5 |
Tabla 7 Resultados de los análisis de X2 para las técnicas de cacería y ordenes de insectos consumidos por las aves durante su alimentación.
| Datos de presencia ausencia | ||||
| Técnica | N | Grados de libertad | Ji Cuadrada | Prob> Ji Cuadrada |
| Colectar | 22 | 2 | 2.245 | 0.325 5 |
| Remover | 22 | 2 | 2.794 | 0.247 4 |
| Datos de frecuencia | ||||
| Colectar | 22 | 8 | 6.675 | 0.572 1 |
| Remover | 22 | 2 | 2.794 | 0.247 4 |
| Ordenes de insectos consumidos | ||||
| Coleóptera | 34 | 12 | 14.081 | 0.295 6 |
| Hymenoptera | 34 | 2 | 1.351 | 0.508 9 |
Los resultados del ACS para corte y sacudida de ramas derivados del ARP muestran una asociación en la inercia total (28.84%), reconociendo la conformación de tres grupos de aves que hacen uso de las familias de insectos registrados (Figura 1). Las técnicas de colectar y remover tuvieron sociación, mostrando la conformación de tres conjuntos avifaunísticos que hacen uso de estas técnicas (Figura 2). El consumo de insectos señala asociación (47.87%), lo que indica la conformación de cuatro bloques de aves que consumen estos insectos (Figura 3). Lo que sugiere que, durante su alimentación, las aves establecen estrategias tróficas que les permite tener dietas diferenciales a partir de la utilización de los insectos disponibles.
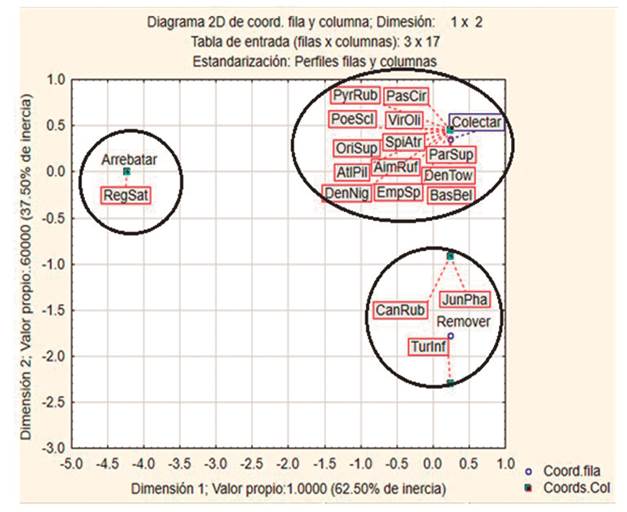
Figura 1: Representación gráfica de los resultados del ACS para las familias de insectos colectados por las aves durante su alimentación en las elevaciones 2600, 2800, y 3000.
Figura 2: Representación gráfica de los resultados del ACS para las técnicas de cacería utilizadas por las aves durante su alimentación en las elevaciones 2600, 2800, y 3000.

Discusión
La tendencia de las técnicas de cacería empleadas por las aves en este estudio, coincide con lo reportado por Pineda-Pérez et al. (2014) en un estudio sobre ecología trófica de aves insectívoras, quienes mencionan que varias especies avifaunísticas presentan pico grueso, lo que les permite capturar coleópteros. Esto ha sido confirmado por Colorado (2004) quien comprobó que la morfología del pico y de las patas explican los insectos que capturan las aves. Los patrones de cacería exhibidos concuerdan con lo reportado por Adamik y Korñan (2004) quienes mencionan que las aves se alimentan de presas disponibles en la corteza, empleando la técnica de colectar. Sobre lo mismo Cueto y López de Casanave (2002) reportan que las aves Passerinas utilizan las técnicas de colectar y revoloteo. Mientras que Caicedo-Argüelles y Cruz-Bernate (2014), señalan que la alimentación es la actividad en que las aves invierten más tiempo y las técnicas de alimentación que utilizan son recoger y capturar. En contraste, Camphuysen y Webb (1999) y Ostrand (1999) mencionan que las aves integran grupos de alimentación, como resultado de la conglomeración de insectos, lo que ceba a predadores que interactúan entre sí; pero de acuerdo con Mills (1998) éste comportamiento se explica a través del reclutamiento visual en el que las aves que se alimentan atraen a otras, por lo que funcionan como indicadoras catalíticas.
Los órdenes de insectos consumidos concuerdan con lo reportado por Perovic et al. (2018) en un estudio de la dieta de aves insectívoras, como controladoras de plagas, que consumieron fueron Coleóptera e Hymenoptera (Haro 1998). Lo que concuerda con Rosas-Espinoza et al. (2008) quienes estudiaron la dieta de Melanerpes formicivorus, encontrando que además de bellotas de Quercus castanea, Q. viminea, Q. resinosa y Pinus magnoliifolia, también consumen insectos de los órdenes Coleóptera y Lepidóptera, además de savia de árboles. Por su lado, Recher y Majer (2006) evaluaron el efecto de la exclusión de aves mediante la depredación de Artrópodos de dosel en un bosque de eucalipto, encontrando que los Coleópteros y Lepidópteros larvales son los más consumidos debido a su alta disponibilidad, por lo que dichas aves evaluaron el efecto de la exclusión de aves mediante la depredación de Artrópodos de dosel en un bosque de eucalipto, encontrando que los Coleópteros y Lepidópteros larvales son los más consumidos debido a su alta disponibilidad, por lo que dichas aves controlan algunos Coleópteros considerados plaga.Sobre lo mismo Figueroa y Corales (2003) reportan que las presas más ofrecidas a los críos de Picoides lignarius fueron los Coleópteros. En contraste Gámez-Virués et al. (2010) encontraron que la mayor disponibilidad de insectos presentes en estratos arbóreos son Araneae y Formicidae. Al respecto Gámez-Virués et al. (2007) señalan que el estudio del espectro trópico en la determinación de las dietas en aves insectívoras podría coadyuvar en el conocimiento de su historia natural con fines de manejo del hábitat.
Las tendencias evaluadas en los comportamientos alimenticios, coexistencia y segregación trófica exhibidos por las aves del presente estudio, coincide con lo reportado por Chávez-Villavicencio et al. (2012) quienes estudiaron la segregación de aves insectívoras con base a la morfometría del pico y su longitud total, hallando que la competencia interespecífica hace que las especies puedan coexistir en simpatría, considerando la distancia ecológica de segregación que disminuye la competencia, lo que hace que las aves coexistan. Sobre lo mismo Pacheco y Castilla (2000) demostraron que dos especies de aves ostreras coexisten debido a que una consume presas de textura blanda y la otra dura, por lo que no hay competencia. En contraste, Velasco (2016) señala que los estratos suelen ser una variable que podría o no incidir sobre la distribución de insectos y por lo tanto, sobre su distribución y consumo, impactando de diversas formas la coexistencia y segregación trófica en este grupo taxonómico. Al respecto Peña et al. (2010) mencionan que la especialización del hábitat y procesos como la dispersión, podrían ser un factor determinante en la coexistencia de especies. Al respecto, Chablé-Santos et al. (2012) evaluaron el efecto trófico de Streptopelia decaocto sobre especies nativas de Yucatán, encontrando segregación por competencia de alimento, espacios de anidación y por portador virus que afecta a las aves nativas.
Conclusiones
Se determinó la composición de las dietas y coexistencia alimenticia en aves insectívoras de las elevaciones evaluadas. La comunidad avifaunística exhibe coexistencia y segregación trófica en la partición de los recursos alimenticios que les permite su ensamblaje en las elevaciones analizadas. Este es el primer estudio sobre la ecología básica alimenticia de este grupo en esta región estudiada. La altitud en bosques templados no es un factor determinante en la coexistencia.

