











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Los modelos de distribución de especies han evolucionado, el primero fue un programa sencillo como el BIOCLIM (Nix 1986), el modelo utiliza variables climáticas y detecta condiciones ambientales adecuadas para las especies. Después surgieron los modelos de regresión logística y lineal como GLM (Generalised Linear Models) (Austin y Smith 1989). Debido a los problemas derivados del cambio climático, deforestación, pérdida de biodiversidad, entre otros que se presentan en las áreas naturales del mundo, surge la necesidad de implementar nuevas herramientas y tecnologías para la modelación de hábitat de especies forestales, por lo que los modelos BIOCLIM, GLM, GARP (Genetic Algorithm for Rule-set Prediction) y GAP (Groups Algorithms Programming) se llevaron a la inteligencia artificial (Stockwell 1999). La combinación de estos modelos y el desarrollo de los sistemas de información geográfica (SIG), generaron nuevos métodos para la modelación geográfica, como MaxEnt, procedimiento de modelación de nicho ecológico que requieren de variables ambientales y registros de presencia de la especie, que se usan para producir modelos o mapas de distribución potencial de especies (Graham et al. 2004).
Hasta finales de la década pasada, se tenían aproximadamente 15 métodos para la modelación del hábitat de las especies (Martínez 2010). Siendo MaxEnt uno de los métodos de mayor precisión (Kumar y Stohlgren 2009), el cual esta basado en la máxima entropía donde se tienen datos de presencia de especies (Phillips et al. 2006). El método MaxEnt se ha aplicado en la ecología (Valdés et al. 2012), conservación de la biodiversidad (García et al. 2012, Ávila et al. 2014), especies en riesgo de extinción (Kumar y Stohlgren 2009, Aguirre y Duivenvoorden 2010), en la evaluación del impacto del cambio climático sobre la distribución de organismos (Lawler et al. 2009) y en la fauna silvestre (Ortíz et al. 2012, De la Torre y Torres-Knoop 2014). Tambíen se ha aplicado en el área forestal para la detección temprana de enfermedades (Meentemeyer et al. 2008), para la búsqueda de zonas afectadas por plagas forestales (Salinas et al. 2010) y para detectar nuevas áreas semilleras de especies amenazadas (Shaily et al. 2010).
Las especies de pinos piñoneros P. cembroides, P. culminicola y P. nelsonii, están adaptadas al clima y la topografía de la sierra madre oriental, P. cembroides presenta amplia distribución en México y es fuente importante de ingresos económicos para los pobladores locales. Mientras que P. nelsonii y P. culminicola tienen distribución restringida (Farjon y Styles 1997, Estrada et al. 2014) con estatus de protección y amenazadas de acuerdo con la NOM-059, por lo que la generación de información con modelos de distribución potencial es importante para establecer planes de manejo, para las especies con estatus de conservación (Austin y Meyers 1996). Por lo anterior el objetivo fue generar modelos de distribución potencial para P. cembroides, P. culminicola y P. nelsonii del noreste de México por medio de las variables ambientales.
MATERIALES Y MÉTODOS
Integración de registros geográficos de las especies
La fuente de registros de presencia de las especies fue de la Red Mundial de Información sobre la Biodiversidad REMIB (CONABIO 2015) de donde se descargaron los registros (coordenadas geográficas) de presencia de las especies, los cuales se validaron con la distribución conocida de la especie (Perry 1991, Farjon y Styles 1997, Estrada et al. 2014), eliminando registros con coordenadas erróneas, por lo que se obtuvieron 208 registros de P. cembroides, 89 de P. culminicola y 67 de P. nelsonii.
Obtención y edición de cubiertas SIG de variables ambientales
Del sitio web de WorldClim se obtuvieron cubiertas geográficas de 19 variables ambientales denominadas Bioclim (Tabla 1), éstas representan las históricas anuales de 1950 al 2000 de temperatura y precipitación (Hijmans et al. 2005). Son cubiertas de tipo raster en formato grid genérico compuestas por pixeles con resolución de 30 segundos de arco ( 1 km), por lo que primero se recortaron para la región noreste de México. Después con el programa ArcMap 10.2 se transformaron a formato grid ASCII. De la página web WorldClim se extrajo la cubierta raster global de altitud, que se usó para obtener la pendiente y la exposición en grados.

Modelación de la distribución potencial de las especies
La distribución potencial de las tres especies se modeló bajo los siguientes escenarios: a) con el set de 19 variables bioclimáticas y b) con 19 variables bioclimáticas más tres variables topográficas. En todos los casos se empleó el modelo logístico con un máximo de 500 iteraciones, creando curvas de respuesta y pruebas Jacknife para validar los modelos con el 50% de registros para pruebas de entrenamiento y el resto para las pruebas de modelación. El modelo resultante consiste en una cubierta geográfica formato grid con valores probabilísticos de 0 a 1 de nicho o hábitat potencial, donde 0 indica pixeles que no representan hábitat y valores cercanos a 1 representan mayor probabilidad de ocurrencia de la especie (Phillips et al. 2006).
En la modelación se consideraron pruebas de precisión a través de un análisis de omisión y comisión, una prueba de sensibilidad gráfica de área bajo la curva (AUC) y pruebas de Jacknife que analizan el comportamiento independiente de cada variable bioclimática para determinar su peso y grado de participación en la creación del modelo. Los valores de AUC generados de ambos escenarios se evaluaron entre sí para determinar el mejor modelo para la especie, donde los valores más altos de AUC indican un mejor modelo. De las variables que conformaron el modelo, se obtuvieron sus valores y se realizaron histogramas de frecuencia para conocer los rangos o límites de tolerancia de la especie (García et al. 2012).
Cálculo de superficie del hábitat potencial
De los valores de probabilidad (0 - 1) del mejor modelo de cada especie y usando el reclasificador de ArcMap 10.2 se ordenaron los valores en intervalos de 0.1 a partir de los cuales se calculó su superficie por decil para conocer el área potencial correspondiente.
RESULTADOS
Modelación de las especies con MaxEnt
En la Tabla 2 se muestran los valores de AUC de ambos escenarios planteados para la modelación MaxEnt para P. cembroides, P. culminicola y P. nelsonii en el noreste de México. El ajuste de los modelos en ambos escenarios es bueno; el valor de AUC encontrado en estas especies es de los más altos reportados en la literatura. La especie P. cembroides tiene amplia distribución con valor menor (0.998) en relación al AUC de las otras dos especies que son de distribución restringida (0.999). La Figura 1 muestra el modelo resultante de P. cembroides, donde se observa la amplia superficie de distribución potencial (0.8 - 0.85). En la Figura 2 se observa el resultado de la modelación de P. nelsonii, donde los valores más cercanos a 1 (0.7 - 0.75) se ubican en los límites de Nuevo León y sur de Tamaulipas. Para P. culminicola, el modelo generado muestra que los valores de 0.6 a 0.7 (Tabla 3) se concentran en las zonas de alta montaña entre Coahuila y Nuevo León (Figura 3).
Tabla 2: Valores AUC generados para denotar el ajuste del modelo de predicción a través de MaxEnt en P. cembroides, P. nelsonii y P. culminicola en el Noreste de México


Figura 1: Distribución potencial de P. cembroides en una porción de la SMO, México, basado en variables Bioclim y topográficas. Los últimos dos deciles se consideran como la zona con mejor probabilidad de presencia de la especie
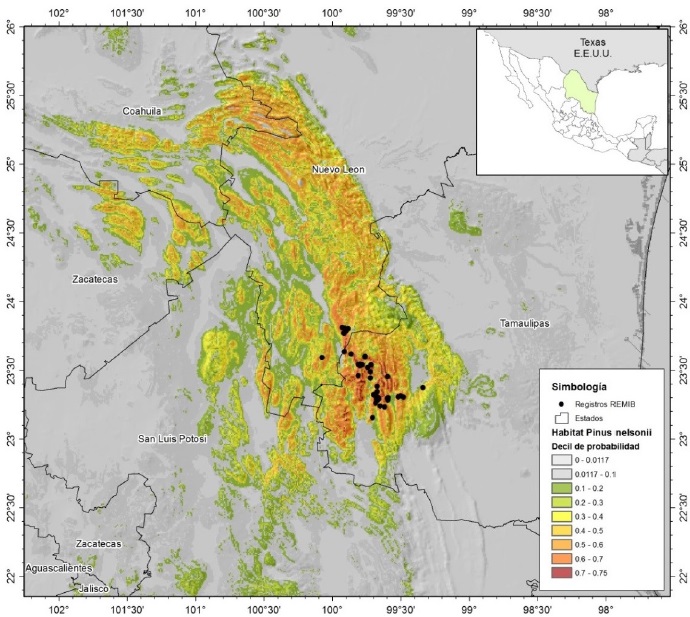
Figura 2: Distribución potencial de P. nelsonii en una porción de la SMO, México, basado en variables Bioclim y topográficas. Los últimos dos deciles se consideran como la zona con mejor probabilidad de presencia de la especie.
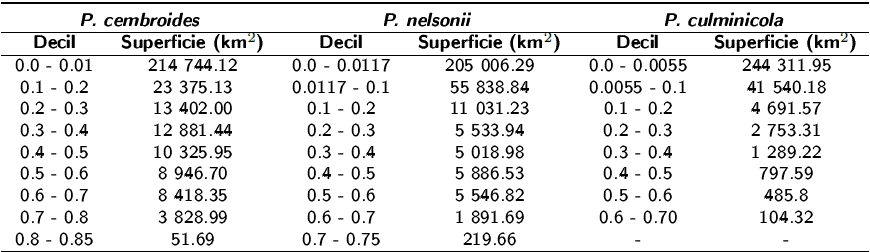
Tabla 3: Superficie por decil del modelo de hábitat potencial de P. cembroides, P. nelsonii y P. culminicola en el Noreste de México. Los decíles (0.7 - 0.85) son considerados como la zona de hábitat más probable

Contribución de las variables en la modelación y mapas de distribución
Las variables de mayor aportación al modelo generado para P. cembroides son la temperatura media del cuarto anual más seco (Bio09), temperatura máxima del mes más cálido (Bio05) y la pendiente con 26.9, 11.6 y 10.1% respectivamente (Tabla 4), con límites mínimos de 8.2, máximo de 18.7 y media de 12.4 °C (Figura 4a); mínimo de 21.1, máximo de 36.7 y media de 28.2 °C (Figura 4b) y mínimo de 0.3, máxima de 24.1 y media de 7.4 grados (Figura 4c), respectivamente. La precipitación del cuarto anual más húmedo (Bio16) tuvo 8.8% de participación, ubicando a P. cembroides como una especie con preferencia por lugares con lluvias de mediana intensidad en alguna temporada del año, el resto de otras 18 variables no pasan de un 7% de aportación o menor (Tabla 4).
Tabla 4: Porcentajes de contribución de variables ambientales en la modelación MaxEnt de distribución potencial de P. cembroides, P. nelsonii y P. culminicola en el Noreste de México


Figura 4: Histogramas de frecuencia de tres variables de mayor influencia en los modelos de distribución potencial de: Pinus cembroides (a - c), Pinus nelsonii (d - f) y Pinus culminicola (g - i) en una porción de la SMO, México.
La altitud y la pendiente fueron variables importantes en la predicción del hábitat de las especies de distribución restringida (P. nelsonii y P. culminicola), aportando al modelo 21.1 y 44.4% la altitud; 15.0 y 18.3% la pendiente, para ambas especies, respectivamente. Otra variable que aportó al modelo de predicción de ambas especies fue la temperatura mínima (Tabla 4); para P. nelsonii la temperatura mínima del cuarto anual más frío (Bio11) varió de 2.8 a 7.2 con media de 4.6 °C (Figura 4d); la altitud fue de 1 070 a 2 474 m (Figura 4e) y la pendiente osciló de 2.2 a 15.9, con media de 7.5 grados (Figura 4f). Para P. culminicola la variable temperatura media del cuarto anual más frío varió de 4.7 a 10.5 °C (Figura 4g), la altitud mínima fue de 2 232 y la máxima de 3 575 m (Figura 4h), mientras que la pendiente en la que se distribuye la especie es en promedio de 15.0 grados (Figura 4i), el resto de las variables aportan al modelo menos del 8%.
El modelo de MaxEnt predice para P. cembroides una distribución amplia en la Sierra Madre Oriental al sur de Nuevo León, Sureste de Coahuila y posible presencia en las sierras aisladas al centro del estado, y parte de Tamaulipas (Figura 1). De la misma forma, MaxEnt predice la mayor probabilidad (0.70 - 0.75) de ocurrencia de P. nelsonii en la zona suroeste de Tamaulipas entre los municipios de Miquihuana, Bustamante, Jaumave, Palmillas y Tula, además de Dr. Arroyo y Zaragoza en Nuevo León, se distribuye en las partes bajas de la sierra de San Antonio, Peña Nevada en zonas de ecotono entre los bosques templados altos y los matorrales áridos colindantes con el altiplano desértico.
La mejor condición potencial, decil de 0.70 para la distribución de P. culminicola ocurre zonas en Galeana Nuevo León, en el Cerro El Potosí, cima de un grupo de cordilleras de la sierra entre los límites de Coahuila y Nuevo León, como La Viga, La Martha, Potrero Ábrego, El Coahuilón y San Antonio (Figura 3), sitios con una elevación dentro de la distribución conocida de la especie.
Análisis de la superficie de hábitat potencial de las especies estudiadas
De acuerdo con el modelo generado para P. cembroides, la superficie estimada oscila entre 51.6 y 3 828.9 km2, lo que representa entre 0.01 y 1.31% de la superficie de los tres estados del Noreste de México. Para P. nelsonii la superficie potencial es de 219.6 km2. De las tres especies modeladas, P. culminicola es la que menos superficie potencial posee con 104.3 km2 (Tabla 3), todo esto considerando superficie estimada a partir del decil 7 (probabilidad > 0.7).
DISCUSIÓN
Considerando el criterio de la cercanía al valor de 1.0 como un mejor modelo (Phillips et al. 2006, Soberón 2011) se supone que el mejor escenario para predecir la distribución potencial de P. cembroides es aquel que incluye las 19 variables Bioclim más las variables topográficas, con valores AUC mayores en el set de datos y en el de entrenamiento, donde se establece la influencia de la topografía en la modelación de la distribución potencial de éstas especies (García et al. 2012). Para especies de distribución restringida (P. nelsonii y P. culminicola) ambos escenarios predicen de forma adecuada la distribución potencial, por lo que puede usarse cualquiera de ellos. Estudiando estas mismas especies pero con otro fin, Aguirre y Duivenvoorden (2010) obtuvieron valores AUC de 0.895, 0.933 y 0.990 para P. cembroides var. cembroides, P. nelsonii y P. culminicola respectivamente, valores altos que variaron con los del presente estudio por utilizar distintas cubiertas ambientales, de menor resolución además de haber usado una fuente distinta de registros de presencia (BRAHMS). Las principales variables predictoras para P. cembroides (Bio09 y Bio05) indican una adaptación de la especie a ambientes de temperaturas templadas con veranos cálidos, característica de bosques de piñonero de las zonas de altiplano (Perry 1991, Farjon y Styles 1997) y se asocia hasta con ocho comunidades vegetales (Estrada et al. 2014). Al respecto Passini (1982) reporta que la distribución de P. cembroides en la Sierra Madre Oriental se encuentra entre 250 y 400 mm de precipitación anual y de 17 a 20 °C de temperatura. El modelo generado en este estudio al igual que el de Aguirre y Duivenvoorden (2010)
corrobora la amplia distribución potencial de P. cembroides, y reportada por otros autores (Estrada et al. 2014).
La especie P. nelsonii se distribuye entre los 1 600 y 2 300 msnm, en zonas con precipitación anual entre 300 y 600 mm, y se ubica en pies de montaña de ambientes áridos a semiáridos (Perry 1991, Farjon y Styles 1997). Mientras que Estrada et al. (2014), reportan que distribuye en ecotonos entre matorrales, chaparrales de encino y bosques de P. cembroides. En lo referente a P. culminicola se reporta que se distribuye de los 3 000 a los 3 700 msnm, bajo condiciones de vientos fuertes, exposición preferentemente norte, creciendo a bajas temperaturas (Perry 1991, Farjon y Styles 1997), es una de las especies más vulnerables a la reducción de su hábitat por efecto del cambio climático (Gómez y Arriaga 2007). Lo resultados encontrados junto a lo reportado por Aguirre y Duivenvoorden (2010), refuerzan la observación de que P. nelsonii y P. culminicola presentan una distribución restringida, aunque existen regiones modeladas donde no se tiene registro, donde MaxEnt predice hábitat potencial que puede ser considerado para promover actividades de conservación, ya que son especies en estatus de protección y amenazadas.
Las variables que mejor predicen la distribución potencial del género Pinus, sobre todo de especies de distribución amplia se relacionan con las temperaturas altas (Cruz et al. 2012), para especies que poseen distribución restringida, la altitud predomina sobre temperatura, no obstante esto parece ser específico, ya que para especies de Abies la altitud y exposición predominan sobre las temperaturas (Martínez et al. 2016). Las variaciones en el ajuste del modelo y de la distribución potencial de las especies depende del tipo de variables bioclimáticas, escala y número de registros de especies (Aguirre y Duivenvoorden 2010, Martínez et al. 2016).
La superficie calculada de P. culminicola, se considera sobreestimada debido a que en el pasado, incendios forestales mermaron de forma significativa la superficie original. Por ejemplo, en 1998 la superficie de P. culminicola en el Cerro El Potosí se redujo un 40%, reportándose únicamente 30 ha (Estrada et al. 2014), por lo que las 10 400 ha calculadas con el modelo deben interpretarse como zonas con el bioclima y la topografía óptimas para su posible distribución o futuros estudios de repoblación.
CONCLUSIONES
Las variables Bioclim y topográficas modelan de forma adecuada la distribución potencial de P. cembroides, P. culminicola y P. nelsonii; siendo las que mejor definen el hábitat potencial la temperatura máxima y media de los meses más secos, para P. cembroides la temperatura mínima del mes más frío, la altitud en P. nelsonii, y para P. culminicola la altitud y la temperatura media del más frío. Para las tres especies, la pendiente determina su distribución potencial. El modelo predice amplia distribución potencial de P. cembroides, que puede incrementarse en el futuro debido a que el modelo integró variables dependientes de temperatura máxima y discriminó aquellas de derivadas de precipitación. Las áreas de mayor probabilidad de ocurrencia de P. nelsonii y P. culminicola dependien de la altitud, la pendiente y las temperaturas frías.

