











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
The freshwater drum Aplodinotus grunniens belongs to a group of fish of the Sciaenidae family that includes organisms living in sea, salt and fresh water. This family comprises 70 genera with 270 species (Nelson 1994) that are distributed practically in all continents and constitute a major worldwide fishery resource. The freshwater drum is distributed throughout the Americas, in Canada from Quebec to Manitoba and Saskatchewan; in the US, east of the Rocky Mountains in San Lorenzo-Great Lakes, the Hudson Bay, the Gulf of the Mobile Bay in Alabama and Georgia, the Mississippi river, and in Central America, in the Gulf of Mexico, in the Usumacinta, Grijalva and San Pedro rivers and in southern Guatemala (Froese and Pauly 2011). The first studies of A. grunniens date back to 1950 and include information on its biology, feeding habits, spawning, and description of the egg, embryo and larva development (Edsall 1967). More recently, Rypel et al. (2006) studied its growth and age variation in rivers (lotic environment) where these fishes are significantly stronger and have longer life spans (˃30 years), versus Alabama reservoirs (lentic environment). This same author conducted a study on sexual dimorphism in four Alabama rivers and found significant differences in the size of females and males (Rypel 2007). Other studies have been carried out in other species of the family on reproductive biology and population structure (Sirangelo and Ferreira 2004, Ramos-Miranda et al. 2009, Ferreira et al. 2011, Pombo et al. 2013, Militelli et al. 2013).
In Mexico, A. grunniens fishery takes place mainly in the states of Veracruz, Tamaulipas and Tabasco, with a reported production of almost 196.49 t (CONAPESCA 2016). In the state of Tabasco fish biodiversity registry, this species is known under the common name of roncador de agua dulce (freshwater snorers or croakers) and has been catalogued as a vicarious species of the Usumacinta river basin (Espinoza-Pérez and Daza-Zepeda 2005). However, fishermen from the municipalities of Emiliano Zapata, Jonuta, Balancán and Tenosique (river zone) call it topuche or roncador. Investigations in Mexico on its biology, ecology, and fisheries are scarce; therefore, the objective of this study was to determine relevant aspects of the reproductive and population biology of Aplodinotus grunniens over an annual cycle.
MATERIALS AND METHODS
A total of 593 specimens of A. grunniens were collected between january and december 2013 from the commercial fishery of the Usumacinta river, Tenosique, Tabasco. The capture area covers the regions described by the inhabitants as the Usumacinta river Canyon Boca del Cerro (17o 25' 33" North and 91˚ 29' 29" West ); the Isla (17o 28' 25" North and 91o 31' 40" West), the Recreo (17o 29' 05" North and 91o 25' 91" West); Chaculji (17o 29' 57" North and 91o 26' 48" West), and the Copo (17o 34' 74" North and 91o 29' 89" West). The specimens were captured during the low water months using seine nets with a mesh size of 2 cm, 80 m in length and 3.0 m in height; during the periods of increased water level, fish were captured using hook and line with crayfish Procambarus llamasi as bait. Fish were killed rapidly by a blow to the head.
From each of the organisms collected, the total length (TL) was recorded using a standard ictiometer of 100 cm ± 1mm precision; the total weight (TW), liver and gonad weight were determined using a Scout ProTM digital scale of 4 000.00 g and ± 0.01 g precision. Both sex and sexual maturity were determined on the basis of gonad morphological characteristics and color in accordance with a maturation scale for partial spawning fish proposed by Nikolsky (1963) that comprises six stages: immature (I), quiescent (II), maturing (III), mature (IV), spawning (V), and spent (VI). Monthly male: female ratios were calculated for different maturity stages. The gonadosomatic index (GSI) was determined with the equation GSI=Wg/Wt*100, where Wg= gonad weight, and Wt= specimen weight (Rossendllum et al. 1987). The hepatosomatic index (HSI) was determined with the equation HSI= Wh/Wt*100, where Wh= liver weight, and Wt= specimen weight (Rodríguez-Gutiérrez 1992). The physiological state or condition factor (K) of each fish was obtained using the expression K=W/Lb*100 proposed by Ricker (1958).
The total length-weight relationship was calculated separately for both sexes by applying the mathematical relationship: TW= aTLb, proposed by Ricker (1975), where TW is the total weight, a is the intercept, TL the total length, and b the slope (growth coefficient). The average size at sexual maturity, defined as the size and age at which 50 % of fish are sexually mature (L50) and when all individuals are fit to actively participate in the reproductive process (L100), was determined on the basis of accumulated relative frequencies of male and female individuals, in maturity stages II and V (Luksenburg and Pedersen 2002), using the logistic equation method of King (2007): P=1/( 1 + exp r(L-Lm) ), where r = slope of the curve, and Lm = mean length of maximum gonad length to which 0.5 (50 %) of the ratio correspond.
Absolute fecundity was also calculated, understood as the number of mature or maturing oocytes found in the ovary. Estimates were conducted by taking three subsamples of 0.3 g (±0.001) in three sections of the gonad (anterior, middle and posterior), and the equation Fa= n*G/g was used; where G = total weight of gonad, g = weight of subsample, and n = average number of oocytes (Bagenal 1978). Relative fecundity was estimated with the formula Fr = Total oocytes/weight of fish in g. In each of these techniques weight was obtained using a Scout ProTM digital scale of 4 000.00 g and ± 0.01 g precision. Climate data such as water temperature (oC), dissolved oxygen (DO), rainfall (mm), and river water level (masl) were taken from the records of the National Water Commission.
The median weight and length values of males and females were compared using the Mann-Whitney (W) test since data did not meet the parametric criteria (Zar 1999). A multiple correlation analysis and ANCOVA were applied to the TL-TW regressions between sexes to identify possible differences between them (Sparre and Venema 1998). The sex ratio per month was compared using a Chi square test (X2) (Underwood 1997). Growth allometric equations were obtained with a logarithmic transformation (Sokal and Rohlf 1996). A Student's t-test (t) was applied to the value of slope b to determine the type of growth (Ibáñez and Fernández 2006). All the statistical analysis were performed considering an error margin α = 0:05.
RESULTS
Of the total number of specimens examined, 327 were males and 266 females. The males had a TL between 23.32 to 39.05 cm and a most frequent size of 29.00 cm. In females, TL was between 24.90 to 46.10 cm, with a most frequent size of 30.00 cm (Figure 1). Males showed significantly smaller weight and length with medians = 262.60 g and 29.71 cm, than females with medians = 362.85 g and 32.05 cm (Mann-Whitney, W = 23 700.50 and 23 430.00, p < 0.001). The analysis of the length-weight relationship between females and males indicates that there are statistically significant differences between models (ANCOVA, F1,589 = 144.46, p < 0.001). The relationship for females was TW=0.0007(TL)3.7834 and for males TW=0.0079(TL)3.0693. For both sexes the model obtained was TW=0.0013(TL) 3.5985, and the percent of data variability explained by the model (R2) was 90.78 % for females, 81.22 % for males and 89.17 % for both sexes. The observed value of b for the species (both sexes) is greater than three, an indication that the species presents an allometric-type growth (t2 = 4.48, p < 0.05) (Figure 2). Males represented 55.15 % (n=327) of analyzed organisms, while females represented 44.85 % (n = 266), leading to a male: female ratio of 1.2:1. Males showed a higher ratio in the months of june (71 %) and october (72 %), while females had maximum values slightly over 50 % in april (54 %), august (55 %), and december (52 %). The total male and female ratios were statistically different in january. The general ratio between sexes showed statistically significant differences (X211 =21.40, p = 0.02) (Figure 3). In july and september only females were captured (1 and 2, respectively) due to the high river level. For this reason, the values of GSI, HSI, and K in males were impossible to determine for these months. The maximum GSI values in females occurred during june (3.78), august (3.95), september (4.29), and october (3.48). In the remaining months this value was below 2.5. In males, the maximum value appeared in october (1.02), and it was below 1.0 in the other months (Figure 4). The hepatosomatic index (HSI) values for females showed their maximum values in august (0.91) and october (1.00), while the minimum was recorded in July (0.56). In the case of males, the maximum value was recorded in december (0.82) and the minimum in April (0.46) (Figure 4). The estimated K values had little variability throughout the year for both sexes, showing a slight increase for females in august, september, and october. The K maximum values recorded for both sexes occurred in august (females 1.65 and males 1.41). The minimum values observed were 1.35 for females (july) and 1.24 for males (february and april) (Figure 5).
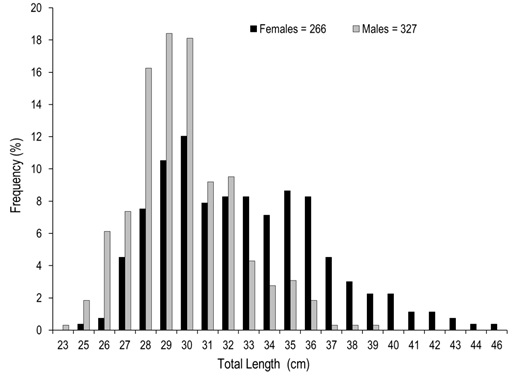
Figure 1. Total length frequency distribution of A. grunniens collected in the Usumacinta River, Tabasco, Mexico.
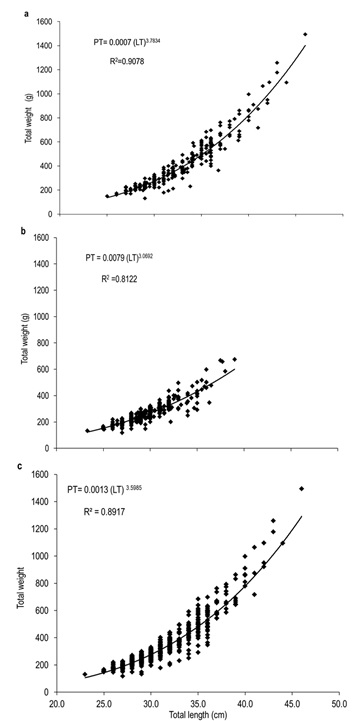
Figure 2. Length-weight relationship of A. grunniens from the Usumacinta River, Tabasco, Mexico, (a) Females, (b) Males, and (c) both sexes (N=593).
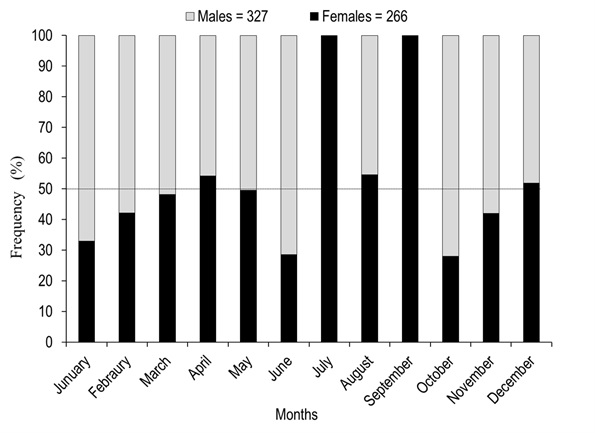
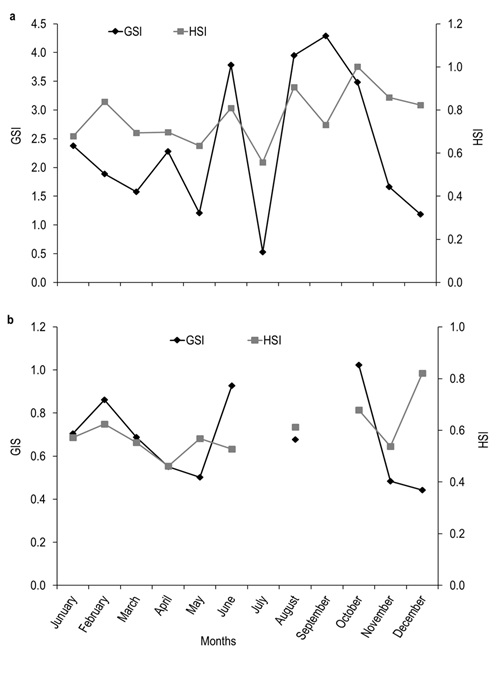
Figure 4. Gonadosomatic index (GSI) and hepatosomatic index (HSI) in (a) females and (b) males from the Usumacinta River, Tabasco, Mexico. Empty spaces in the lines indicate lack of organisms during sampling (N=593).
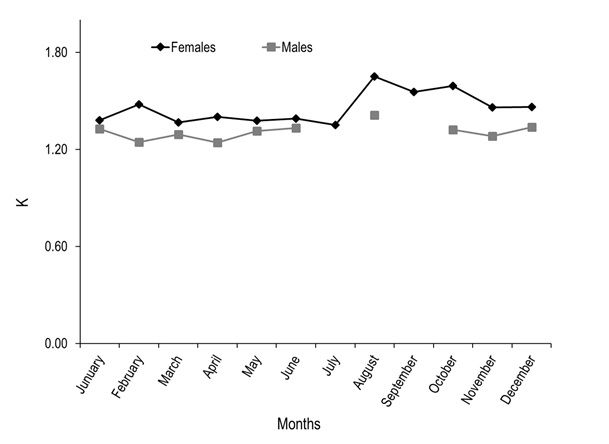
Figure 5. Condition factor (K) in females and males of A. grunniens from the Usumacinta River, Tabasco, Mexico. Empty spaces in the lines indicate lack of organisms during sampling (N=593).
The average absolute fecundity (± SD) estimated for the species (n = 50) was 68 055.17 (± 49 120 .62) oocytes, within a range of 9 632 to 223 800. The obtained relative fecundity was 138.74 (± 74.30) oocytes/g of female weight, within a range of 33.43 to 323.87 oocytes/g of weight.
In terms of maturity stages, we found that the most frequently recorded stages in females were quiescent (II) and spawning (V). Females in maturing stage (III) showed the highest value in september, october, and november. Females in mature stage (IV) were found in february, april, june, october, and november. With the exception of september, there were females in spawning stage (V) throughout the reproductive cycle, with march, april, june, and august standing out for their higher frequency. Spent females (VI) were more frequently recorded in april, june, july, and october. Two females analyzed in July showed the higher frequency (Table 1, Figure 6). In regards to males, the quiescent (II) and maturing (III) stages were the most frequent during the annual cycle. The mature stage (IV) displayed the maximum values in february and october. The male specimens in spawning stage (V) were recorded throughout the annual cycle, but with a higher frequency in march and june. Spent males (VI) showed their maximum frequency in june and august, respectively (Table 1, Figure 6).
Table 1. Macroscopic description of gonad maturity stages of A. grunniens from the Usumacinta River, Tabasco, Mexico (based on Nikolsky, 1963).

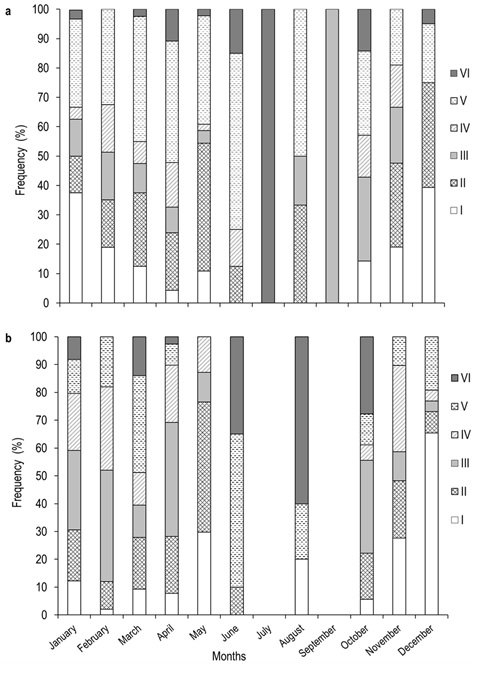
Figure 6. Maturity stage distribution frequency in (a) females and (b) males of A. grunniens from the Usumacinta River, Tabasco, Mexico. Empty spaces indicate lack of organisms during sampling.
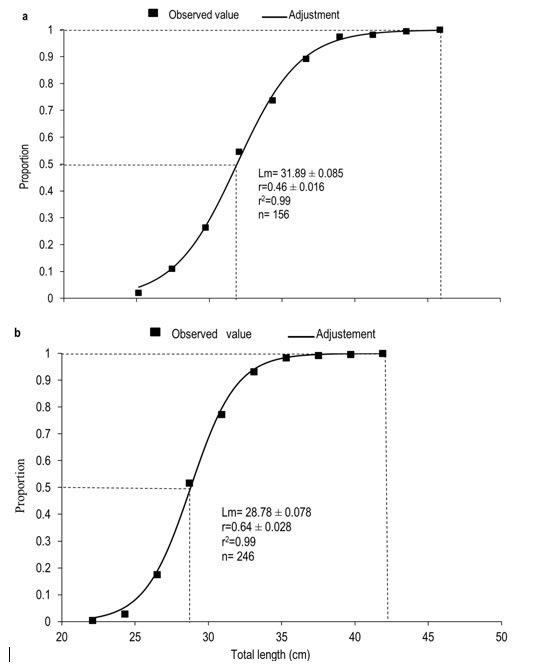
The average length at first maturity in females was 31.89 cm, the length at which they are fit for spawning (L100) was 45.85 cm of TL. In males, the average length at first maturity was 28.78 cm and all males are fit for spawning from 41.90 cm of TL (Figure 7). Throughout the annual cycle studied, a mean water temperature of 26 (± 1.44) oC was recorded in the Usumacinta river, with a minimum of 24 oC in december and a maximum of 29 oC in May. The DO recorded an average of 8.25 (± 0.62) mg L-1, with a minimum of 7 mg L-1 during june and a maximum of 9 mg L-1 during april, may, july, and august. pH averaged 7.83 (± 0.38), with a minimum of 7.00 in april and september and a maximum of 8.00 for the remaining months of the annual cycle. Rainfall averaged 8.70 (± 5.33) mm, with a minimum of 4 mm in april and a maximum of 19 mm in November. The river level was 15.18 (± 2.78) masl, with a minimum of 11.70 in april and may and a maximum of 18.90 in september and november, respectively.
DISCUSSION
Despite the small mesh size used (2 cm) for seining, during the period of study of A. grunniens, a size distribution between 20.00 and 46.00 cm (TL) was recorded. These values are similar to the range recorded by Chávez et al. (1989) at the San Pedro river in Tabasco, Mexico, which was from 19.00 to 47.10 cm. Although there are no studies reporting freshwater drum juveniles or larvae in the area, we speculate that early stages of the life cycle may occur in the lower portion of the river, since floating eggs and larvae are transported downstream by currents. Furthermore, Rypel et al. (2006) reported that size distributions of freshwater drums captured in Alabama habitats using electrofishing and gill nets, were skewed toward juveniles in lentic reservoirs and adults in rivers; proposing that freshwater drum make movements of some scale across aquatic landscapes during their lives. Marcano et al. (2002) suggest that size structure depends on the environment they live in, mainly in terms of food availability, population density and ecological factors. Therefore, the recording of large sizes in our sampling grounds may be conditioned by a migratory behavior of the species, as is the case with other fish species. Reports of the early life history of this species indicate that movements between habitats may also be associated with changes in diet (Swedberg and Walburg 1970). This behavior stands out as an adaptation to exploit the seasonal productivity in different areas, at different times of the year, and among specific feeding, habitat and spawning areas (Saborido 2005). Our data suggest that freshwater drum use this portion of the Usumacinta river as spawning grounds, using other areas of the system as nursery grounds. Unfortunately, there have been no attempts at documenting this behavior.
In this study, A. grunniens females were significantly bigger than males, a condition similar to that reported by Rypel (2007) in the lakes and rivers of Alabama, US. However, it differs from the data recorded by Edsall (1967) in Lake Erie, where the specimens showed similar sizes for both sexes. The difference observed in the size of females may be attributed to intrinsic (genetic) factors of the species, type of habitat, and time of year (Gómez and Guzmán 2005). Nevertheless, Saborido (2005) considers that the main factor in controlling growth, survival, and reproduction is the energy acquired by fish through feeding.
The observed length-weight relationship showed an allometric-type growth similar to that reported by Chávez et al. (1989), for organisms captured in the San Pedro river, Balancán, Tabasco, Mexico. In relation to the allometry observed in A. grunniens, Jacquemin and Pyron (2013) pointed out that body size contributes more importantly to morphological variation than sex variations or river location. At the same time, they established that the interaction between body size, maturity, and river location indicates that an individual's morphology is not necessarily a direct relation between the fish and their environment, but rather it is multi-faceted and results from the combination of allometry, sex and river location (local habitat). They also mention that the most noticeable change in size occurs during ontogeny, the time when embryos, larvae and/or juveniles grow, and that many allometry examples are ontogenetic in type. This allometric condition is similar to that of other Sciaenidae species, such as the Micropogonias furnieri in the Gulf of Paria, State of Sucre, Venezuela, and the Cienega Grande de Santa Marta in the Caribbean region of Colombia (Gómez and Guzmán 2005, Cancio et al. 2006). The data of this study differed from those recorded for other marine origin species of the same family with isometric growth, such as Stellifer lanceolatus, from the southern region of the Gulf of Mexico, and Pogonias cromis, in northeast Florida (Ramos-Miranda et al. 2009).
The sex ratio obtained in this study is close to an equilibrium between males and females in the population (55 % males: 45 % females), so apparently this species conforms to fisher´s theory on equal sex ratios. In other studies with A. grunniens and despite using different sampling methods, several authors have reported proportions close to the one found in this study, indicating slight biases toward one or the other gender. Jacquemin and Pyron (2013) reported a ratio of 1.01:1.00 in specimens captured in the Wabash river, being similar to the ratio recorded by Edsall (1967) for the same species in Lake Erie. In turn, this ratio also resembles the one reported by Grua et al. (2009) for marine origin species of the same group (Sciaena umbra and Cynoscion albus) in the Gulf of Montijo, Panama, and in the Balearic Islands, Spain, respectively, while Potts et al. (2010) reported 1.00:1.40 for Argyrosomus coronus in the mouth of the Kunene Estuary and Baia Farte, Angola. On the other hand, Chávez et al. (1989) in the San Pedro river and Rypel (2007) in different habitats in Alabama found sex ratios favoring females (0.65:1.00 and 0.71:1.00, respectively).
The GSI is used in gonadal development studies as an indicator that helps describe the reproductive cycle. In this work, four reproductive peaks were noted in june, august, september, and october for females, while there were only three peaks for males in february, june, and october. The GSI consistency for both sexes in some months establishes a synchronization of reproduction. These data indicate that A. grunniens is a sequential spawning species that reproduces most of the year. Similar results were reported for southern US, in the Ohio river, where the spawning of this species spans over five months, from april to september. However, the spawning season in the western region of Lake Erie was reported for june, july, and november, and for may and june in Texas, with temperatures ranging between 18 to 26 oC (Wallus and Simon 2006, Chad et al. 2007). In the case of Mexico, gonads in an advanced stage of maturity were observed most of the year in the San Pedro river, Tabasco (Chávez et al. 1989). Moreover, in the current study A. grunniens showed a reproductive behavior that is similar to that of other Sciaenidae species, such as Larimichthys polyactis (yellow sea bass), Sciaena umbra (banded sea bream), and Plagioscion squamosisimus (sea bass), in Korea, the Balearic Coast, Spain, and the Ucayali Coast, Peru, respectively (Grau et al. 2009, Lim et al. 2010).
The HSI in females is considered as an indicator directly linked to the reproductive cycle and decreases right before spawning (Rodríguez-Gutiérrez 1992). This relationship is based on the fact that the liver produces vitellogenin during vitellogenesis, a protein that is taken by oocytes in development as reservoirs of nutrients and energy for embryos and larvae. Taking into account this physiological activity, the female HSI in this study shows a decrease in march, april, may, and july, while in males it is reduced in march, april, june, and november. The trend is not clear since this index decreases like the GSI and later diverges in the other sampling months. Nevertheless, to ensure that reproduction is established in this way, it is necessary to have a greater number of specimens during july and september. Moreover, Grau et al. (2009) pointed out that there is a similar behavior in Sciaena umbra, where both the HSI and GSI decrease from may to august, and then diverge from september to october.
In this study, the K factor indicates that, during the reproductive cycle, A. grunniens showed a good condition, was feeding and using energy constantly for the maturation of the ova mainly from february to june. However, the values of K increased from august to december, the period with the highest rainfall in the study area. Hence, this increase may be related to the accumulation of energy in the form of fat, an adaptive physiological strategy used for generating an energy reservoir as observed for the Menticirrhus littoralis species (Sirangelo and Ferreira 2004). Rypel et al. (2006), noted that the lipids and physical condition of A. grunniens are significantly related with the type of habitat and vary with the age of the fish. However, this increase in the value of K may be conditioned by food availability during the period of increased river level which, in turn, reduces the possibility of feeding for species like this that have benthonic habits and mainly consume clams, bottom crustaceans and worms, and occasionally fish (Jacquemin and Pyron 2013). Nevertheless, the observed variation ok K may be interpreted as the period of sexual conditioning of the population prior to reproduction and spawning. This factor is associated with the biological and ecological phenomena of recruitment, growth, maturity and spawning like in the case of Cynoscion arenarius, in the southern Gulf of Mexico, and Euthynnus lineatus, in the coastline of Oaxaca, Mexico (Ramos-Cruz 2009).
In regards to fecundity, Rypel (2007) mentions that A. grunniens is a highly fecund species (> 1 million eggs). However, in this study, female fecundity (n=50) was significantly lower than this value, as well as for other reported fecundity data for the species, like the findings of Chávez et al. (1989) in the San Pedro river, Balancan, Mexico, and Daiber (1953), in Lake Erie, as well as the case reported by Chad et al. (2007) for a 3.5 kg female in Texas with 600 000.00 eggs. For the Lewis and Clark Lake, US, 6 to 9-year-old females (n=17), with a TL between 30.70 and 38.60 cm and a weight of 295.00 to 670.00 g, were reported to have a range of 34 000.00 to 65 500.00 eggs. However, the average fecundity of 68 055.00 eggs recorded in the current study is within the ranges mentioned above. This variation may be due to differences in maximum sizes of females present in the Usumacinta system. In this sense, Wallus and Simon (2006) established that the reproductive potential variation of the species may be modified by feeding, reproductive status, age, length and weight, as well as temperature.
The highest proportion of sexual maturity (stage V) found in this study for the period from january to june and from october to december, with an average temperature of 27oC, is in line with the study carried out in the San Pedro river, Balancán, Tabasco, Mexico, as well as for the data recorded from the beginning of May to july in Texoma Lake, from may to july in the Mississippi river, and in june for Lake Winnebago and Lake Erie, and Canada, respectively. It differs from those of mature specimens captured in the Tennessee river, in september (Chávez et al. 1989, Wallus and Simon 2006).
The mean lengths at first maturity (L50) of A. grunniens in this study were higher than those determined for females (28.6 cm) and males (25.1 cm) of the San Pedro river, Balancán, Tabasco, Mexico, as well as those of another species of the family in the Balearic Islands, Spain (Chávez et al. 1989, Gil et al. 2013). However, they are similar to the values obtained in marine Sciaenids, such as the cat-fish (Plagioscion squamosissimus) in Ucayali, Peru, Micropogonias furnieri in the Gulf of Paria, Sucre, Venezuela, Macrodon ancylodon in the Estuary of Caete, northern Brazil; and from those reported for Plagioscion magdalenae in the Bay of Marajo, estuary of the Amazon river (Gómez and Guzmán 2005, Barbosa et al. 2010). This shows that L50 may be the consequence of genetic variability and size among the species of Sciaenidae. The data generated by L50 are relevant since they are used as a management strategy for fishing management plans, for the species to be captured at sizes greater than those of first maturity (Perera-García et al.2008).
CONCLUSIONS
Aplodinotus grunniens can reproduce practically during the entire year. The species presents a marked sexual dimorphism, with females being larger than males, and differences in the mean maturity length and growth indicators. This research provides baselines to propose a strategy for fisheries management in the region, with the intention of incorporating strategies for the conservation of the species.

