











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En la región tropical de México con frecuencia la finalización de corderos se efectúa en condiciones de estabulación con dietas integrales que permitan una ganancia diaria de peso (GDP) mayor a 180 g y conversión alimenticia menor a siete (Muñoz-Osorio et al. 2015). En este sistema de producción se evita la exposición de los animales a los efectos negativos de los factores climáticos del sistema bajo pastoreo (Zamora-Zepeda et al. 2015) y se reduce la posibilidad de infestación con nematodos gastrointestinales (González-Garduño et al. 2011, López et al. 2013). Sin embargo, el alto costo económico de las dietas integrales de tipo convencional y el manejo en estabulación pueden disminuir las utilidades del proceso de finalización (González-Garduño et al. 2013). Por lo que es necesario considerar el uso de recursos alimenticios disponibles en el trópico, que puedan integrarse a la dieta o como complemento alimenticio que aporten proteína y almidón, sin elevar el costo de la ración y con resultados similares a las dietas con ingredientes convencionales (Luna-Palomera et al. 2010).
Entre los recursos disponibles se encuentran las semillas o granos de Canavalia ensiformis. La harina de sus semillas contiene 3.35 Mcal EM kg-1 de materia seca (MS) (Martín y Palma 1999), en tre 18.8 y 29.8 % de proteína cruda (PC) (Ajayi et al. 2010, Doss et al. 2011) y entre 24.7 y 56.7 % de almidón; lo que la hace atractiva desde el punto de vista nutricional (Zamora 2003, Sivoli et al. 2007). Sin embargo, el uso de harina de semillas de canavalia se encuentra limitado por factores antinutricionales (FAN) (Akanji et al. 2015). Además de la baja o nula disponibilidad comercial de semillas (Cáceres et al. 1995) y de las pocas publicaciones que se tienen sobre su empleo en ovinos (Domínguez-Bello y Stewart 1990, Hernández- Montiel et al. 2016). Por lo anterior, el objetivo de la presente revisión es presentar los resultados que se han obtenido en la productividad y estado de salud de los ovinos que consumen C. ensiformis como parte de su alimento.
Potencial productivo de la canavalia
En México, la leguminosa C. ensiformis no se utiliza como ingrediente alternativo en la alimentación de los ovinos a nivel comercial. Además de la poca información sobre la influencia que tiene su consumo en los ovinos (Cáceres et al.1995, Hernández-Montiel et al. 2016). La semilla de C. ensiformis tiene amplias posibilidades de uso en la alimentación de rumiantes debido a que proporciona forraje y semillas (Villanueva-Avalos et al. 2010). Es una planta anual que se desarrolla en la regiones tropicales y subtropicales, debido a que puede crecer en temperaturas de 14 a 27 °C, con precipitación de 711 a 4 191 mm. Se adapta a un amplio rango de condiciones edáficas, prosperando en suelos ácidos con pH de 4.0 a 6.5, aunque su desarrollo es mayor en pH de 6.5 (Sheahan 2012).
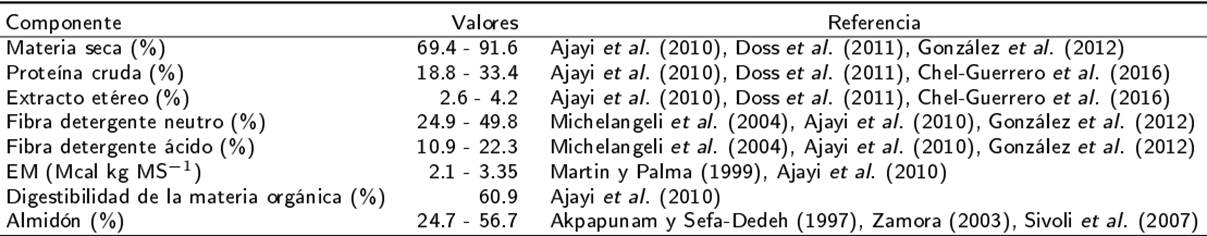
Las semillas maduras se cosechan de los 180 a los 300 d, con producción máxima de 4.6 t de semilla ha-1, aunque es más frecuente cosechar 1 t ha-1 (Sheahan 2012). En el estado de Yucatán, México, se reportan rendimientos de entre 1 y 3.8 t de semilla ha-1, pero se requieren realizar estu dios de tipo agronómico (Hernández-Montiel et al. 2016, Chel-Guerrero et al. 2016). La importan cia de la canavalia como alimento para rumiantes radica en su aporte de proteína cruda, almidón y fibra (Sivoli et al. 2007, Ajayi et al. 2010). De acuerdo con Estupiñán et al. (2007) el forraje de canavalia tiene 52.3 % de fibra detergente neutro (FDN), 37.6 % de fibra detergente ácido (FDA), 56 % de digestibilidad de la FDN y 49.5 % de di- gestibilidad de la FDA (Tabla 1).
Concanavalina a y canavanina en las semillas de canavalia
Los FAN y sustancias tóxicas presentes en C. ensiformis protegen a la planta contra el ataque de bacterias, virus, insectos y herbívoros (Rosenthal 1991); pero, este tipo de sustancias son tóxicas para los animales (Ologhobo et al. 2003). En la Tabla 2 se indican los principales FAN detectados en las semillas de C. ensiformis, destacando concanavalina fracción A (Con A) y canavanina debido a que son los principales FAN presentes en C. ensiformis, cuya presencia en el organismo animal se asocia con efectos negativos en el consumo de alimento e integridad y funcionalidad del epitelio intestinal (Rueda et al. 2003). En las semillas de C. ensiformis se encuentran las proteínas globulares Con A y concanavalina fracción B (Con B). La primera es la más abundante, pudiendo representar el 23 % de la proteína total de la semilla, mientras que Con B representa el 0.9 % de la proteína total (Schlesier et al. 1996). La Con A es considerada una lectina con propiedades importantes debido a su capacidad para propiciar interacciones entre las células, ya que contiene sitios de unión específicos para residuos α-D-glucosilo y α-D-manosilo presentes en la superficie de células animales y microorganismos (Murray et al. 2013).

Con A tiene relevancia desde el punto de vista nutricional, debido a que reduce la eficiencia produc tiva de los animales, afectando de forma negativa la digestión de los alimentos, al reducir la contribución de los microbios en la digestión y alterar la integridad del epitelio intestinal (Udedibie y Carlini 1998, Ologhobo et al. 2003). Los daños en la integridad del epitelio intestinal asociados al consumo de Con A afectan de forma indirecta el rendimiento productivo del animal, debido a que inhibe la actividad enzimática, reduce el nivel de consumo de alimento y compromete la salud (Larue-Achagiotis et al. 1992, Méndez et al. 1998). Las lesiones en la mucosa intestinal afectan los mecanismos de defensa natural del animal, lo que reduce su capacidad para contener las bacterias inocuas, que al pasar de la luz intestinal hacia la linfa, sangre e hígado, generan la muerte (Ologhobo et al. 2003). Con A puede unirse a polisacáridos como el almidón y formar precipitados, lo que reduce la disponibilidad de los carbohidratos para su digestión y absorción (Udedibie y Carlini 1998). Resultados de estudios in vitro indican que Con A reduce la actividad de la pepsina y amilasa afectando la tasa de digestibilidad de la proteína y del almidón (Thompson y Gabon 1987). La interacción entre Con A con los microbios que habitan el tracto intestinal pueden generar la aglutinación de las bacterias al unirse a Con A con los carbohidratos de superficie de los microorganismos, lo que reduce la contribución de los microor ganismos en la digestión de los alimentos, además de interferir con la capacidad de adhesión de las enterobacterias con la mucosa intestinal (Jaffe 1980). La formación de uniones de Con A con las glucoproteínas y glucolípidos del epitelio intestinal, afectan la integridad y funcionamiento del epitelio al inhibir la actividad enzimática de las microvellosidades de los enterocitos y la eficiencia de absorción de nutrientes (Sridhar y Seena 2006).
La L-canavanina es un aminoácido no proteínico que se encuentra en las semillas, hojas, tallos y raíces de la C. ensiformis y puede representar entre el 4 y 5 % del peso seco de las semillas (Zamora 2003). En la función digestiva de los animales adquiere importancia debido a su analogía estructural con la L-arginina y su capacidad para inhibir el transporte de aminoácidos (Rueda et al. 2003). En diversos estudios efectuados en microbios, insectos y células humanas se ha demostrado que la L-canavanina afecta la formación y función de las proteínas que tienen dentro de su composición L-arginina y que el efecto es más marcado cuando el medio contiene una baja concentración de arginina (Swaffar et al. 1994). En las aves, la L-canavanina inhibe el transporte de L-lisina, L-metionina y L-arginina dependiente del Na+ en la membrana apical del enterocito (Rueda et al. 2003). En las dietas balanceadas para animales de granja es frecuente que estas contengan alta concentración de arginina (Udedibie y Carlini 1998).
Procesamiento de las semillas de canavalia para reducir su toxicidad
La eliminación o reducción de los FAN de las semillas de C. ensiformis son esenciales para aprovechar el valor nutricional en la alimentación de los animales (Chel-Guerrero et al. 2016, Hernández- Montiel et al. 2016). En términos generales, los efectos negativos de los FAN en las semillas de canavalia pueden ser eliminados, reducidos y/o inhibidos, si las semillas reciben algún tratamiento físico, químico ó biológico antes de ser consumidas (Elias et al. 2009, Vadivel y Biesalski 2012, Owen et al. 2015). Con frecuencia, la aplicación de sólo un tratamiento a las semillas no logra eliminar los FAN de forma efectiva, por lo que se requiere utilizar dos o más tratamientos antes de su incorporación como parte de los alimentos (Akande y Fabiyi 2010, Doss et al. 2011, Alagbaoso et al. 2016).
Se tiene varios procesos tecnológicos que se pueden aplicar a las semillas de C. ensiformis para eliminar, reducir o inactivar los FAN y para alterar la estructura del almidón (Tabla 3), con la finalidad de mejorar el acceso de los gránulos al ataque enzimático y la utilización de los nutrientes (Sivoli et al. 2007, Emenike et al. 2016). En términos generales, los principios utilizados para el procesamiento de las semillas de canavalia son el fraccionamiento de la semilla, temperaturas elevadas con o sin humedad, alta presión, remojo en agua, tratamientos químicos, fermentación y germinación (Akande y Fabiyi 2010). También se ha encontrado que al someter dietas que incluyen harina de canavalia a la fermentación, se reduce la presencia de FAN y se aumenta la calidad nutritiva (Elias et al. 2009).
Tabla 3: Influencia de algunos métodos de procesamiento sobre los factores antinutricionales en semillas crudas de C. ensiformis.

La mayor parte de los estudios para evaluar la influencia de la incorporación de semillas de canavalia en la dieta, se han realizado con harina de semilla cruda (Dixon et al. 1983, Domínguez- Bello y Stewart 1990). Asumiendo que el rumiante dispone de una población de microbios en el rumen capaz de eliminar las toxinas, así como tolerar o adaptarse a la presencia de los FAN presentes en las semillas de canavalia (Domínguez-Bello 1996). Algunas semillas de leguminosas como Clitoria ternatea tienen propiedades defaunantes al reducir la actividad de los protozoarios, lo que puede aumentar el flujo de proteína de origen microbiano al intestino delgado (Ajayi et al. 2010). De ahí la importancia de establecer la capacidad de los ovinos para tolerar diferentes niveles de inclusión de harina de semillas de canavalia, sin que se afecte el rendimiento productivo y la salud (Dixon et al. 1983, Domínguez- Bello y Stewart 1990).
Al incluir harina de semillas de canavalia en el alimento para ovinos se debe considerar que el almidón de semillas de leguminosas presenta digestibilidad de entre 7 y 26.1 %, la cual es baja con respecto a la de almidones de cereales y tubérculos (Sivoli et al. 2007). La presencia de diversos FAN (Con A, canavanina y taninos) con efectos negativos, fundamentalmente en los monogástricos, se debe a que generan reducción del consumo de alimento (Ologhobo et al. 2003). Aunque existen diversos procesos para reducir, eliminar y/o inhibirá los FAN de la canavalia (Akande y Fabiyi 2010), no hay estudios en donde se haya evaluado la respuesta productiva de los ovinos alimentados con una dieta en la que se haya aplicado un proceso a las semillas de C. ensiformis o al alimento antes de incorpo rarse a la dieta (Valiño et al. 2015). Como la extrusión para favorecer el uso de semillas de canavalia (Mustafa et al. 2003).
Cambios en la población microbiana del rumen
En ovinos West African el consumo de dietas con 0, 20 y 40 % de semilla de canavalia no generó cambios en el número de bacterias viables totales, bacterias celulolíticas y hongos. Al parecer, la población total de bacterias ruminales no se afecta por los FAN de las semillas de canavalia, lo que explica la tolerancia de los rumiantes. Sin embargo, en muéstreos de líquido ruminal se detectó un in cremento en la población de bacterias Gram negativas conforme se aumentó el nivel de inclusión de canavalia. Esto se atribuye a un compuesto antibacterial no identificado que actúa de forma selectiva sobre estas bacterias (Domínguez-Bello y Stewart 1990). Por otra parte, se requieren realizar estudios que demuestren la proporción del nitrógeno de la semilla de canavalia que se incorpora como proteína microbial, sin que se presenten problemas de toxicidad y viabilidad para las bacterias del rumen (Castillo-González et al. 2014, Calabró 2015). Se tiene evidencias derivadas de estudios efectuados en bacterias, insectos y células de humanos que indican que la incorporación de L-canavanina en sustitución de L-arginina afecta la formación y función de proteínas vitales para el crecimiento y sobrevivencia de las células (Rosenthal 1991, Swaffar et al 1994). Una línea de investigación no explorada, se relaciona con la posible capacidad de interacción entre la Con A con los sitios de unión específicos (α-D-glucosilo y α-D-manosilo) presentes en la superficie de las células epiteliales del rumen (Styriak et al. 1994). Este tipo de interacción puede afectar la adhesión de las bacterias a la superficie del epitelio del rumen, membranas de las células vegetales y gránulos de almidón presentes en el rumen y/o favorecer su aglutinación (Choudhury 2015). No se encontraron estudios que aborden el posible daño que produce la Con A al epitelio del rumen, el cual puede afectar el transporte de nutrimentos.
Los protozoarios del rumen adquieren importancia debido a que pueden contribuir con el 60 % de los productos de la fermentación microbiana en el rumen y con el equilibrio del pH ruminal. La mayoría de los protozoarios presentes en el rumen- retículo son ciliados, aunque existen algunas especies flageladas (Kamra 2005, Choudhury 2015). En bovinos alimentados con heno de pasto Taiwan (Pennisetum purpureum) complementado con harina de canavalia, se encontró que los animales que recibieron el complemento con canavalia se incrementó el número total de protozoarios, aumentando la población de protozoarios flagelados y se redujo la concentración de protozoarios holotricos y entodinomorfos. En contraste, los animales con alimento sin complementado de canavalia se incre mentaron los protozoarios holotricos y entodinomorfos (Sandoval-Castro y Herrera 2001). En ovinos alimentados con una dieta con 40 % de follaje de Leucaena leucocephala y 60 % de Pennisetum purpureum, disminuyó la concentración de protozoarios entodinomorfos con respecto al tratamiento 100 % de P. purpureum, la reducción en la población de protozoarios se atribuyó a un alto consumo de FAN presentes en la L. leucocephala (Barros-Rodríguez et al. 2015). En rumiantes, el consumo de follaje y semillas de las leguminosas tropicales C. ensiformis, Acacia angustissima, A. farnesiana, Calliandra calothyrsus Enterolobium cyclocarpum and Lysiloma latisiliquum se asocia con defaunación (Sandoval-Castro y Herrera 2001, Monforte-Briseño et al. 2005).
Los protozoarios ruminales se encuentran en el material fibroso con lenta tasa de pasaje en el rumen y pueden adherirse a partículas de alimento y al epitelio del retículo. Mientras que las bacterias ruminales pueden adherirse a los protozoarios, es posible que este tipo de unión sea por la interacción lectina-carbohidrato (Choudhury 2015). Para rumiantes alimentados con canavalia, la Con A puede interferir en la capacidad de adhesión de los protozoarios al epitelio ruminal y al alimento (Murray et al. 2013). La demostración y significancia de la interferencia de la Con A con la capacidad de fijación de los protozoarios y bacte rias ruminales es un tema de investigación que se debe abordar en estudios futuros (Choudhury 2015). Los protozoarios tienen la capacidad de engullir y lisar bacterias, de modo que en un escenario en donde las bacterias hayan incorporado la canavanina como parte de sus proteínas, se debe establecer si la síntesis de proteínas de la degradación de proteínas con canavanina ocasionan daños o si los sistemas enzimáticos de los protozoarios permiten la degradación de L-canavanina en canalina (Rosenthal et al. 1989).
Los hongos ruminales representan aproximadamente el 8 % de la biomasa en el ecosistema ruminal, los cuales son de tipo anerobio y se encuentran adheridos al interior de fragmentos vegetales, lo que contribuye en la digestión de la fibra (Castillo-González et al. 2014). Es probable que la integridad y funcionalidad de los hongos ruminales se afectada por los FAN de canavalia, debido a que C. ensiformis produce isoformas de ureasa con acción insecticida y fungicida (Becker-Ritt et al. 2007). La ureasa más abundante es JBURE-I, seguida por la canatoxina y JBURE-IIB, la toxicidad de la ureasa de canavalia ocurre en dosis micromolares y varía con el género y especie de hongo, causando inhibición de la proliferación, alteraciones morfológicas en la pared y membrana, cambios en el transporte de H+ y en la permeabilidad de la membrana, lo que induce a la muerte celular (Postal et al. 2012).
Los bacteriófagos son los virus de bacterias y se encuentran en el rumen en gran número (Kamra, 2005). Este tipo de microrganismos afectan a las bacterias y pueden ocasionar su lisis, participando de esta forma en el recambio de la masa bacterial en el rumen (Choudhury 2015). La influencia de los FAN de la canavalia sobre los bacteriófagos no ha sido estudiada. Sin embargo, estudios in vitro indican que L-canavanina afecta el proceso de replicación de algunos virus (Robertson et al. 1984). La relación que existe entre los microbios presentes en el ecosistema del rumen es compleja, estable y al mismo tiempo dinámica (Kamra, 2005, Choudhury 2015). Los protozoarios compiten de forma eficaz con las bacterias por los sustratos y pueden influir sobre las bacterias mediante predación, de modo que diferencias en el número de protozoarios del rumen influye sobre el número de bacterias (Castillo-González et al. 2014). Estudios efectuados en rumiantes con harina de canavalia indican que los FAN presentes en la canavalia influyen más en las bacterias Gram positivas que en la concentración total de bacterias (Domínguez-Bello y Stewart 1990) y en el caso de los protozoarios del rumen, la incorporación de canavalia reduce la concentración de protozoarios holotricos y entodinomorfos (Sandoval-Castro y Herrera 2001). No hay estudios en los que se evalué los cambios cuantitativos en la población de bacterias, protozoarios y hongos en rumiantes con dietas de canavalia. Este tipo de estudios es importante, debido a que parte de la variación ob servada entre un animal y otro en respuesta a un tratamiento dietético se deben a la tendencia individual a tener distintos cocientes entre bacterias y protozoarios en el rumen (Teather et al. 1984).
Aunque el rumen es considerado un compartimento con una población de microbios capaces de eliminar, procesar e inhibir toxinas presentes en los alimentos (Domínguez-Bello 1996). No se tienen evidencias que apoyen que los microrganismos del rumen tienen la capacidad de procesar y evitar los efectos negativos de Con A y canavanina, por lo que es conveniente evitar la inclusión de harina de semillas crudas en la dieta de rumiantes, y en especial en los animales lactantes que aún no tienen desarrollado el rumen (Jami et al. 2013).
Fermentación ruminal
En ovinos, se han evaluado los efectos de diferentes niveles de inclusión de harina de semillas de canavalia cruda en la dieta sobre indicadores de fermentación ruminal (Dixon et al. 1983). Al respecto, Domínguez-Bello y Stewart (1990) al estudiar el efecto de tres niveles de inclusión de harina de semillas de canavalia en la dieta, encontraron que la inclusión de canavalia, reduce la concentración de N-amoniacal y el valerato con respecto al tratamiento sin canavalia. La reducción en la concentración de N-amoniacal fue acompañada de un incremento en la concentración de aminoácidos y péptidos libres. Por lo que proponen que el decremento en la concentración de N-amoniacal está relacionado con la inhibición de hidrólisis de péptidos. Al respecto Dixon et al (1983) reportan que el pH, N-amoniacal, proporciones y concentración molares de los ácidos grasos volátiles (AGVs) no fue afectada por la inclusión de semillas de canavalia en el suplemento.
Al estudiar la degradación ruminal de la harina de semillas de canavalia en ovinos, se encontró que la tasa de recambio de la fase liquida es mayor en el suplemento sin canavalia con respecto al suplemento con canavalia (Dixon et al. 1983). Mientras que González (2004) al determinar la degradación ruminal de la materia seca y proteína cruda de la harina de C. ensiformis; encontró que la materia seca y la proteína cruda son degradadas en el rumen, y que la degradación efectiva de la proteína cruda no fue afectada con altas tasas de recambio ruminal. Con respecto a la degradación de la proteína cruda, Ekanayake et al. (1999) indican que los aminoácidos no proteínicos pueden constituir más del 33.4 % del nitrógeno total, lo que resalta la importancia de los microbios ruminales en el proceso de degradación de los compuestos nitrogenados no proteínicos. Mientras que Sridhar y Seena (2006) reportan que en semillas de C. ensiformis el 50 % de las proteínas pertenecen a la fracción globulina. Las cuales presentan una alta solubilidad y degradabilidad en el rumen (Wadhwa et al. 1993). La desaparición de las lectinas del rumen indica su grado de solubilidad, aunque no necesariamente su degradabilidad, existiendo la posibilidad de que una parte de las lectinas salga del rumen conservando su capacidad de unión a residuos de carbohidratos y de esta forma tenga influencia antinutricional en el intestino delgado (Dixon y Hosking 1992).
Consumo de alimento y ganancia diaria de peso
Estudios en ovinos para establecer la influencia del nivel de inclusión de harina de semillas de C. ensiformis sobre la GDP y la conversión alimenticia, no han generado resultados consis tentes con respecto a la identificación del nivel de inclusión que permite maximizar la GDP y el consumo de alimento (Domínguez-Bello y Stewart 1990, Hernández-Montiel et al. 2016). Al respecto Dixon et al. (1983) al evaluar tres niveles de in clusión de harina de semillas de canavalia en la ración de ovinos, con una mezcla de harinas de grano de maíz y pasta de soya; encontró que el consumo de alimento fue similar en los tres niveles de canavalia evaluados. Sin embargo, la conversión alimenticia fue afectada por el nivel de inclusión de canavalia, resultando similar entre los niveles 0 y 22 % (7.9 y 8.9, respectivamente), pero, diferente en tre éstas y el nivel de 32 % (10.8). En contraste se reporta que la inclusión de semillas de canavalia en 0, 20 y 40 % de la dieta de ovinos no generó cam bios importantes en el consumo de alimento y en el peso vivo (Domínguez-Bello y Stewart 1990). Sin embargo, Hernández-Montiel et al. (2016) indican que la inclusión de 25 % de harina de semillas de canavalia en un alimento fermentado y sin fermentar afecta el consumo de alimento de los ovinos con respeto a un tratamiento sin canavalia. El menor consumo de alimento en dietas con canavalia se asocia con una menor palatibilidad (Hughes-Jones et al. 1981), digestibilidad de la fracción fibrosa de la materia seca y presencia de FAN en las semillas de canavalia (Chel-Guerrero et al. 2016).
En las hembras, la GDP no fue afectada por el nivel de inclusión de semillas de canavalia (Dixon et al. 1983). Sin embargo, en los machos la GDP si fue afectada por el nivel de inclusión de canavalia, sin que se detectaran diferencias en la GDP en los tratamientos con 0 y 22 % de canavalia (Tabla 4). En contraste, ovinos machos alimentados con un alimento, sin fermentar o fermentado con canavalia, tuvieron una menor GDP con respecto a un alimento sin canavalia; el menor crecimiento fue por un menor consumo de MS, que redujo el con sumo de EM y PC (Hernández-Montiel et al. 2016). Los resultados de Dixon et al. (1983) indican que el sexo del cordero debe considerarse en el diseño de raciones que incluyan semillas de canavalia. Al parecer, las hembras toleran un mayor nivel de in clusión de canavalia sin que se afecte su GDP y su consumo de alimento. Mientras que, en el caso de los corderos, la harina de semillas de C. ensiformis debe incluirse hasta en un 22 % de la ración con el fin de no afectar la GDP y la conversión alimenticia.
Tabla 4: influencia de la inclusión de semillas crudas de Canavalia ensiformis sobre la productividad y estado de salud de los ovinos.

Es probable que la inclusión de entre 25 y 32 % de canavalia en la dieta, permita que se alcance una concentración de Con A y L-canavania que afecta de manera significativa la funcionalidad y viabilidad de bacterias y protozoarios ruminales, y una fracción de estos FAN logren llegar sin cambios al intestino delgado, produciendo daños al epitelio y reduciendo la eficiencia productiva de los ovinos. En corderos en crecimiento y con pesos superiores a 30 kg, no se ha encontrado que el consumo de dietas integrales con harina de semillas de canavalia tenga un efecto negativo sobre el estado de salud (Domínguez-Bello y Stewart 1990, Hernández-Montiel et al. 2016).
Uso potencial de la canavalia en ovinos
Por la resistencia que muestran los nematos gastrointestinales (NGI) a los diversos fármacos disponibles para su control (López-Arellano y Mendoza de Gives 2011). Se han realizado propuestas para el control de los NGI que no impliquen la aplicación de fármacos, destacando aquellos que se basan en el empleo de plantas y semillas con actividad antihelmíntica (Galina y Cuéllar 2009). Entre los diversos principios activos que pueden tener las plantas y semillas con actividad antihelmíntica se encuentran los taninos (Hoste et al. 2006), los cuales forman complejos con proteínas y en menor grado con iones metálicos, aminoácidos y polisacáridos (Makkar et al. 2007). Aunque las semillas de C. ensiformis contienen taninos (Doss et al. 2011, Vadivel y Biesalski 2012), en los niveles en los que se ha estudiado su consumo no se han reportado animales enfermos (Dixon et al. 1983, Domínguez- Bello y Stewart 1990, Hernández-Montiel et al. 2016). La Con A puede ser otro metabolito de canavalia que afecta la viabilidad de las larvas de los NGI, debido a que Con A tiene la capacidad de unirse con residuos de carbohidratos (Murray et al. 2013); que pueden estar presentes en la superficie de las larvas lo que afecta sus funciones vitales (Githiori et al. 2006).
En términos generales, la eficiencia productiva que se obtiene en ovinos con diversos niveles de inclusión de semillas de canavalia cruda en el alimento (Dixon et al. 1983) no es mayor a la obtenida con ingredientes convencionales (Partida et al. 2009). Por lo que es necesario evaluar si la aplicación de diversos procesos físicos, químicos y biológicos a la harina de canavalia permiten su uso en la alimentación de ovinos (Valiño et al. 2015). De manera adicional, se debe considerar que la aplicación de estos procesos implica un costo extra que repercute en el sistema de producción (Hernández- Montiel et al. 2016). Por otra parte, es necesario considerar que los procesos aplicados a las semillas de canavalia no remueven todos los FAN, además de deficiencias en los métodos analíticos para determinar la existencia de varios de ellos (Akande y Fabiyi 2010).
Conclusiones
Aunque hay información sobre el manejo agronómico de la C. ensiformis, el cultivo comercial de esta leguminosa no se realiza en las regiones tropicales de México. La MS y PC de la harina de semillas de C. ensiformis tiene alta degradabilidad ruminal en ovinos, sin que se hayan detectado cambios negativos en la función ruminal asociada al consumo de esta semilla. Los resultados de es tudios sobre los microbios ruminales no establecen que estos sean afectados de forma negativa. Los resultados de los estudios en donde se evalúa la in fluencia de diferentes niveles de inclusión de harina de canavalia en dietas integrales, indican que esta puede incluirse hasta en un 22 % sin que se afecte el estado de salud y ganancia diaria de peso de los corderos.

