











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En los rumiantes, el crecimiento prenatal es influenciado por diversos factores, estacando el nivel de nutrición materna y la capacidad funcional de la placenta. Mientras que el crecimiento postnatal puede ser limitado por la nutrición materna y por factores ambientales (Bell et al. 1987). En el caso particular de los corderos, se sabe que el crecimiento predestete se afecta por la raza, tipo de nacimiento, sexo, edad al destete y el número de parto de la oveja (Hinojosa-Cuéllar et al. 2012).En los estudios en donde se evalúa la influencia del tipo de nacimiento del cordero y el tipo de parto de la oveja, sobre el crecimiento predestete de los corderos, solo se consideran los efectos principales, sin considerar la influencia de la interacción entre ambos factores (Ríos-Utrera et al. 2014).
Se han realizado pocos estudios en ovejas de pelo de clima cálido, en los que se evalúen los cambios de peso de las ovejas y sus camadas como indicadores de la productividad durante la lactancia, considerando el tipo y número de parto de la oveja (Godfrey et al. 1997, Alexandre et al. 2001). No se tiene información sobre el rendimiento y la composición de la canal de corderos de razas de pelo en edad menor de 70 d (Combellas 1997). Ya que estos estudios se han realizado en etapas posteriores al destete (Partida et al. 2009, Macías-Cruz et al. 2010). Por lo anterior, el objetivo del presente estudio fue evaluar la influencia del tipo de nacimiento de los corderos y el número de parto de la oveja, sobre el crecimiento predestete, rendimiento de la canal, composición corporal de los corderos y el comportamiento productivo de las ovejas.
Materiales y métodos
Se estudiaron 25 ovejas Blackbelly x Pelibuey (12 primíparas y 13 multíparas) y sus 39 corderos (20 provenientes de ovejas primíparas y 19 de multíparas). El estudio se llevó a cabo en la unidad ovina experimental del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, localizada en el municipio de Huimanguillo, Tabasco, México (17° 50’ LN, 93° 23’ LO), con clima cálido húmedo, lluvias todo el año (A/) y temperatura ambiente media anual de 27.8 °C (INEGI 2014).
Manejo de la oveja previo al parto y durante la lactancia
Las ovejas durante los 30 d previos a la fecha de parto se mantuvieron en pastoreo y se suplementaron con 300 g oveja-1 d-1 de un alimento comercial que contenía un 15 % de PC. El nivel de suplementación ofrecido cubrió 30 % de los requerimientos de materia seca (MS) y energía metabolizable (EM) de la etapa de gestación de ovejas Pelibuey (Chávez et al. 1995).
En la primera semana de lactación, las ovejas permanecieron estabuladas con sus camadas y se alimentaron con heno de Cynodon plectostachyus. En tanto que, en la segunda semana de lactación se pastorearon de 8 a 14 h, y el resto del día estuvieron con sus camadas recibiendo heno de C. plectostachyus. A partir de la tercera semana y hasta finalizar la etapa de lactación, las ovejas estuvieron en pastoreo de 8 a 18 h, en praderas con C. plectostachyus. El sistema de pastoreo utilizado fue de tipo controlado, la pradera se ocupó durante tres días y se le dio un descanso de 25 d. Durante toda la etapa de lactación las ovejas se suplementaron con 500 g oveja-1 d-1 de un alimento comercial con 15 % de PC. El nivel de suplementación ofrecido permitió cubrir 30 % de los requerimientos de MS y EM estimados para la etapa de lactación de ovejas Pelibuey (Chávez et al. 1995).
En las ovejas se determinó el peso y condición corporal (CC) al parto y a los 56 d posparto. La CC se evaluó utilizando una escala de uno a cinco (Thompson y Meyer 1994). En las ovejas se evaluaron las variables: peso y CC al parto y a los 56 d postparto; cambio de peso durante la lactancia (peso de la oveja al finalizar la lactancia, (kg) - peso de la oveja al parto, (kg) y de CC (CC de la oveja al finalizar la lactancia - CC de la oveja al parto) y cambio de peso diario durante la lactancia como 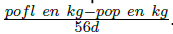 donde pofl = peso de la oveja al finalizar la lactancia y pop = peso de la oveja al parto. De manera simultánea. Se registró el peso de la camada al nacimiento y al destete. Este último valor se utilizó para calcular la eficiencia productiva de la oveja con la formula
donde pofl = peso de la oveja al finalizar la lactancia y pop = peso de la oveja al parto. De manera simultánea. Se registró el peso de la camada al nacimiento y al destete. Este último valor se utilizó para calcular la eficiencia productiva de la oveja con la formula  (Godfrey et al. 1997).
(Godfrey et al. 1997).
Manejo de los corderos durante la lactación
Los corderos estuvieron estabulados toda la etapa de lactación y se alimentaron por amamantamiento controlado, con complemento alimenticio comercial Inicia Corderos con 18 % de PC, que se ofreció a partir del quinto día de edad y heno del pasto C. plectostachyus y la leguminosa Gliricidia sepium a libre acceso a partir del día 15 de edad. El acceso al amamantamiento se controló en la segunda semana de vida de los corderos, que consistió en restringir el acceso del cordero al amaman tamiento de 8 a 14 h. De la tercera semana hasta concluir el período de la lactancia el acceso se controló de 8 a 18 h.
Para calcular la ganancia diaria de peso (GDP) predestete, los corderos se pesaron al nacimiento y al destete a los 56 d de edad. Se sacrificaron al destete con un previo ayuno de alimento no lácteo de 14 h. Posteriormente, se retiró la piel, cabeza, extremidades delanteras y traseras a la altura de la articulación carpo-metacarpiana y de la articulación tarso-metatarsiana, visceras abdominales y torácicas (Martínez et al. 1987). Se registró el peso de la canal caliente y la grasa cavitaria como la grasa interna en las cavidades abdominal y pélvica (Hernández-Espinoza et al. 2012). Posteriormente, la canal se dividió en los cortes cuello, brazo, tórax, abdomen y pierna de acuerdo con Martínez et al. (1987).
El peso vacío fue la diferencia en kg entre el peso al sacrificio y el peso del contenido digestivo, que se obtuvo por diferencia entre el peso del tracto gastrointestinal (rumen, retículo, omaso, abomaso, intestino delgado e intestino grueso) lleno y vacío. El rendimiento en canal (%) se determinó con la formula  . El rendimiento verdadero en % se calculó con la formula
. El rendimiento verdadero en % se calculó con la formula  (Martínez et al.1987). El área del músculo Longissimus dorsi se midió entre la 12a y 13a costilla por medio de una película plástica cuadriculada en cm2 (García et al. 1998). El rendimiento de los cortes primarios en la canal (%) se determinó a través de la ecuación
(Martínez et al.1987). El área del músculo Longissimus dorsi se midió entre la 12a y 13a costilla por medio de una película plástica cuadriculada en cm2 (García et al. 1998). El rendimiento de los cortes primarios en la canal (%) se determinó a través de la ecuación 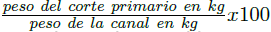 . En los corderos se evaluaron las variables: peso vivo al nacimiento (PN) y al destete (PD); GDP; peso y rendimiento de la canal caliente y verdadero; área del músculo L. dorsi; peso y rendimiento de los cortes primarios en la canal: cuello, brazos, tórax, abdomen, piernas y grasa cavitaria.
. En los corderos se evaluaron las variables: peso vivo al nacimiento (PN) y al destete (PD); GDP; peso y rendimiento de la canal caliente y verdadero; área del músculo L. dorsi; peso y rendimiento de los cortes primarios en la canal: cuello, brazos, tórax, abdomen, piernas y grasa cavitaria.
Diseño experimental y análisis estadístico
Se utilizó un diseño factorial de dos factores; que fueron el tipo de nacimiento (TN) de los corderos (sencillo y múltiple) y el número de parto (NP) de las ovejas (primípara y multípara). En el grupo con nacimiento múltiple se incluyeron los datos de una oveja que al parto y al destete tuvo tres corderos. En el estudio del crecimiento del cordero, la unidad experimental fue el cordero, mientras que en el estudio de la eficiencia productiva de la oveja, la unidad experimental fue la oveja o la camada. Todos los análisis se efectuaron con el paquete estadístico SAS 9.0 (SAS 2002) y se probó su normalidad con la prueba de Shapiro-Wilk's. Los análisis se efectuaron con el procedimiento GLM. En los modelos finales se excluyó la interacción NP x TN, cuando no fue significativa (p < 0.05), mientras que los datos de CC y de variables que no mostraron normalidad se analizaron con la prueba de la suma de rangos de Wilcoxon para datos no pareados (Milton 2001).
Resultados
Crecimiento predestete
La interacción NP x TN no afectó (p > 0.05) el PN, PD y la GDP predestete de los corderos. El NP de la oveja solo afectó (p < 0.05) el PN de los corderos. Los corderos provenientes de ovejas primíparas tuvieron un menor PN con respecto al de corderos de ovejas multíparas, con peso de 2.6 ± 0.14 y 3.1 ± 0.14 kg, respectivamente.
Con excepción del PN, el TN influyó (p <0.05) en el PD y GDP de los corderos. Las medias de cuadrados mínimos ± error estándar (EE) para PN, PD y GDP fueron de 2.9 ± 0.17 kg, 11.8 ± 0.56 kg y 159 ± 8 g en corderos de nacimiento sencillo y de 2.7 ± 0.11 kg, 10.3 ± 0.37 kg y 136 ± 5 g en corderos de nacimiento múltiple, respectivamente.
Producción de la canal
El NP influyó (p < 0.05) sobre el área del L. dorsi. Los corderos provenientes de ovejas primíparas tuvieron una menor área del L. dorsi con ± 0.50 y 8.6 ±0.45 cm2, respectivamente. En el resto de las variables no se detectó (p > 0.05) influencia del NP.
Con excepción rendimiento de la canal, el resto de las variables relacionadas con el estudio de la canal de los corderos fueron afectadas (p < 0.05) por el TN. Los corderos de nacimiento sencillo tuvieron mayor peso y rendimiento de la canal, peso de los cortes primarios, peso de la grasa cavitaria y área del músculo L. dorsi con respecto a corderos de nacimiento múltiple (Tabla 1).
Tabla 1 Influencia del tipo de nacimiento sobre la composición corporal de corderos Blackbelly x Pelibuey
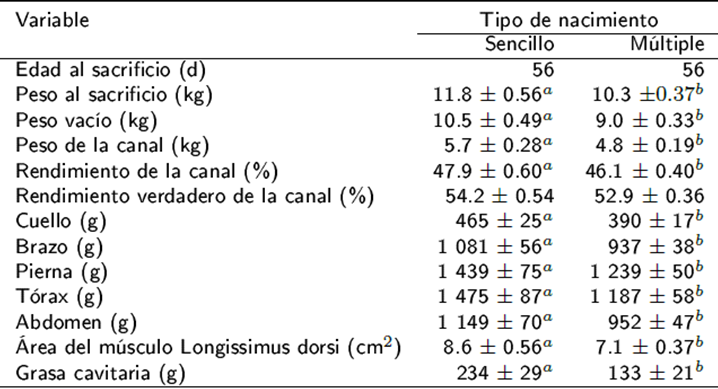
a,bMedias de cuadrados mínimos ± error estándar con diferente superíndice dentro de la misma fila indican diferencia significativa (p < 0.05).
Con excepción del rendimiento del tórax (p <0.05), el resto de las variables relacionadas con el estudio de la canal de los corderos no fueron afectadas por la interacción NP x TN (p > 0.05). Las medias de cuadrados mínimos ± EE para rendimiento del tórax en la canal en primíparas con parto sencillo, primíparas con parto múltiple, multíparas con parto sencillo y multíparas con parto múltiple fueron de 26.4 ± 0.85, 23.7 ± 0.49, 25.4 ± 0.71, y 26.0 ± 0.55 %, respectivamente. Las medias generales ± desviación estándar de los rendimientos de los cortes en la canal fueron de 8.3 ± 1.4 en cuello, 19.5 ± 1.1 en brazo, 25.1 ± 2.1 en tórax, 19.9 ± 2.3 en abdomen y 25.9 ± 2.1 % en pierna.
Productividad de las ovejas
La interacción NP x TP no afectó (p > 0.05) las variables evaluadas en las ovejas y sus camadas. El NP tuvo influencia (p < 0.05) sobre las variables: peso al parto, cambio de peso, cambio de peso diario y CC al parto. No obstante, el NP no afectó (p >0.05) al resto de las variables evaluadas en las ovejas y su camada (Tabla 2). Las variables peso vivo a los 56 d posparto, CC al parto y en el día 56 posparto y cambio en la CC no fueron afectadas (p > 0.05) por el TP. Sin embargo, el TN influyó (p < 0.05) en el resto de las variables incluidas en el estudio de las ovejas y su camada (Tabla 2).

Discusión
Crecimiento predestete
El crecimiento prenatal, expresado como el PN fue afectado por la interacción entre el feto y el ambiente materno. El PN de corderos prove nientes de madres multíparas fue mayor que el de corderos de madres primíparas, lo que coincide con Carrillo et al. (1987) y Ríos-Utrera et al. (2014), quienes reportan que en ovejas de pelo, el PN fue mayor en corderos nacidos de ovejas multíparas que en corderos de ovejas primíparas. Probablemente el útero de las ovejas multíparas tenga mayor volumen que el de las primíparas, al respecto se sabe que un mayor volumen del útero favorece el crecimiento y desarrollo fetal (Gootwine et al. 2007), lo que puede explicar la superioridad en PN de los corderos de ovejas multíparas. También, se ha documentado que factores ambientales como, la edad, talla y número de parto, influyen en el crecimiento fetal (Jainuden y Hafez 1996, Lawrence y Fowler 2002).
Un factor importante que determina el crecimiento fetal, es el número de fetos presentes en la hembra. Se ha reportado que, en ovinos, el PN del cordero disminuye con el incremento del número de corderos nacidos (Gootwine 2005). Sin embargo, en el presente estudio el TN no afectó el PN de los corderos, pesando corderos de nacimiento múltiple 94 % del PN de los de nacimiento sencillo. Un resultado similar fue reportado para ovejas Pelibuey, en el que se encontró que los corderos gemelos pesaron 96 % del PN de corderos de nacimiento sencillo (Macías-Cruz et al. 2009). Mientras que otros estudios, han encontrado que el TN afecta el PN de los corderos, pesando los corderos gemelos entre 82 y 88 % del PN de los corderos de nacimiento sencillo (Hinojosa-Cuéllar et al. 2012, Ríos-Utrera et al. 2014). En relación a la ausencia de influencia del TN sobre el PN de los corderos, al respecto Gootwine (2005) encontró que el PN se reduce con el incremento del tamaño de la camada. Mientras que Greenwood et al. (2000) reportan asociación positiva entre el peso de la oveja al apareamiento con el peso de la placenta. En el presente estudio, las ovejas con un tipo de parto múltiple, mostraron un mayor peso al parto con relación a las hembras de parto sencillo, por lo que es probable que la placenta de las ovejas con parto múltiple tenga mayor peso y eficiencia que la placenta de las ovejas con parto sencillo, lo que puede explicar la ausencia de diferencias en el PN. El PD y GDP de los corderos de nacimiento múltiple representó 91 y 87 % de el de los corderos de nacimiento sencillo. Lo que coincide con Rajab et al. (1992), Hinojosa-Cuéllar et al. (2009) y Ríos-Utrera et al. (2014) quienes reportan que, el PD y GDP predestete están influenciados por el TN, siendo superiores los valores en corderos de nacimiento sencillo que los de nacimiento múltiple. Al respecto se sabe que la frecuencia de amamantamiento de los corderos de nacimiento gemelar es mayor que la de corderos de nacimiento sencillo (Teke y Akdag 2012) y que la producción de leche en ovejas con dos corderos es mayor que la de ovejas con un cordero (Snowder y Glimp 1991). Sin embargo, una mayor frecuencia de amamantamiento y mayor producción láctea no son suficientes para que los corderos de nacimiento gemelar logren un PD y GDP predestete similar a la de corderos de nacimiento sencillo.
El crecimiento predestete es afectado por el nivel de nutrición materna y por factores que influyen en la producción de leche (Bell et al. 1987). En el presente estudio, las ovejas con parto múltiple y sencillo recibieron el mismo manejo nutricional, situación que pudo poner en desventaja a las ovejas con parto múltiple, debido a su mayor demanda de nutrimentos por su mayor peso vivo y por amamantar a dos o tres crías (NRC 2007). Lo que ocasionó menor eficiencia de crecimiento de los corderos de nacimiento múltiple, ya que la leche disponible se tiene que compartir entre el número de crías, y por consiguiente menor GDP.
Producción de la canal
El peso vivo de un animal no siempre tiene correlación con la cantidad de productos vendibles (Owens et al. 1993). De ahí la importancia de identificar los factores que afectan el rendimiento y composición de la canal. Se ha reportado que el TN afecta el peso de la canal de los ovinos de lana en finalización, teniendo los ovinos de nacimiento gemelar 90 % del peso de la canal de los corderos de nacimiento sencillo (McCoard et al. 2015). En el presente estudio, el peso de la canal de los corderos de nacimiento múltiple fue 86 % del peso de la canal de los corderos de nacimiento sencillo. Es posible, que las diferencias en el peso de la canal de corderos de nacimiento sencillo y múltiple se deba al peso de la masa muscular y el consumo individual de leche. Ya que el peso de la masa muscular, está en función del número y tamaño de las fibras musculares. En los fetos gemelos, se ha detectado que estos tienen un menor peso de su masa muscular y concentración de aminoácidos en el músculo semitendinosus, al compararlos con fetos de gestación sencilla (McCoard et al. 2000, Sales et al. 2013). La importancia de los minoácidos radica en que estos están implicados en las vías que regulan la síntesis de proteínas (Wu 2009). El menor peso de la masa muscular de los corderos gemelos al nacimiento, puede explicar el retardo en su desarrollo muscular posnatal. También, se ha reportado que el nivel de producción de leche de las ovejas que amamantan dos crías puede ser mayor (Ramsey et al. 1998, Jawasreh y Khasawneh 2007) o similar al de ovejas con una sola cría (Godfrey et al. 1997). Sin embargo, en el caso de corderos gemelos, se debe compartir la cantidad de leche que produce la oveja, lo que limita el nivel de consumo de nutrimentos. El rendimiento de la canal fue afectado por el TN, la canal de corderos de nacimiento múltiple tuvo 96 % de la canal de corderos de nacimiento sencillo. Pero al considerar el peso vacío del cordero en el rendimiento verdadero de la canal, no se detectaron diferencias. Lo que indica que a una misma edad, el rendimiento verdadero de la canal se mantiene cuando las diferencias en el peso vacío de los corderos son de baja magnitud.
El rendimiento de la canal de los corderos evaluados fue ligeramente inferior al reportado en corderos de pelo de la raza West African sacrificados a los 70 d de edad y con mayor peso (Combellas 1997) y al de corderos de lana de la raza Suffolk Down sacrificados a los 23 d de edad y con menor peso (Pérez et al. 2002). Sin embargo, el rendimiento verdadero de la canal de los corderos fue similar al encontrado en corderos West African y Suffolk Down (Combellas 1997). Las diferencias entre estos estudios se deben al rendimiento comercial de la canal, raza, edad y tipo de nacimiento, ya que en rendimiento verdadero de la canal fueron similares.
En los corderos de nacimiento múltiple el peso de los cortes primarios representó entre 80.4 y 86.7 % del peso de los de nacimiento sencillo. El mayor peso de los cortes primarios en corderos de nacimiento sencillo coincide con lo reportado en corderos de lana sacrificados a 73 y 110 d, en los que el peso de la pierna fue mayor en los de nacimiento sencillo que en los de nacimiento gemelar (McCoard et al. 2010). Específicamente, el peso de la pierna de los corderos de nacimiento múltiple representó 86 % del peso de la pierna de los de nacimiento sencillo. Al parecer, los corderos de nacimiento sencillo tuvieron una ventaja inicial sobre los de nacimiento múltiple en la composición de la masa muscular (McCoard et al. 2000, Sales et al. 2013) y en el consumo individual de leche (Godfrey et al. 1997, Ramsey et al. 1998, Jawasreh y Khasawneh 2007). Esta ventaja se mantuvo hasta el destete y se detectó en el mayor peso de la canal y en el peso de los cortes primarios objeto de la investigación.
El área del músculo L. dorsi en corderos provenientes de ovejas primíparas fue 80 % del área de corderos de multíparas. No hay estudios sobre la influencia del NP de las ovejas, sobre la variación del área del L. dorsi de los ovinos en edades menores o mayores a 90 d. Si se considera que el número de corderos al destete fue similar entre ovejas primíparas (1.5 ± 0.06) y multíparas (1.6 ± 0.06), es probable que los corderos de ovejas multíparas hayan consumido mayor cantidad de leche con respecto a los corderos de primíparas. Un mayor consumo de nutrimentos puede favorecer un crecimiento diferenciado en algunas regiones de la canal. Al respecto, existen evidencias que indican que la producción de leche en ovejas Awassi se incrementó de forma gradual después del primer parto (Jawasreh y Khasawneh 2007). Mientras que Partida y Martínez (2010) indican que el crecimiento de las regiones corporales y composición tisular en la canal ovina es afectado por la concentración energética en la dieta. Por lo que es probable que la mayor área del músculo L. dorsi en corderos provenientes de ovejas multíparas se deba a un mayor consumo de nutrimentos con respecto al de corderos de primíparas.
Los corderos de nacimiento múltiple tuvieron 82.6 % del área del músculo L. dorsi de los de nacimiento sencillo, lo que no coincide con Jucá et al. (2014), quienes encontraron que el TN no afectó el área del músculo L. dorsi en corderos de la raza Santa Ines a los 112 d. Mientras que Sales et al. (2013) señalan que en los fetos gemelos, el peso del músculo semitendinosus fue 17 % menor que el de los fetos provenientes de gestación sencilla. Una situación similar pudiera presentarse en el L. dorsi de los corderos de nacimiento múltiple y mantenerse durante la etapa posnatal, lo que explicaría las diferencias en el área del L. dorsi entre corderos de nacimiento múltiple y sencillo.
El crecimiento incluye el depósito de grasa corporal, aunque la masa muscular es la de interés primario en la producción de carne. Particularmente, el depósito de grasa perineal precede el depósito intermuscular, subcutáneo e intramuscular (Owens et al. 1993). El incremento en la tasa de depósito de grasa en los animales sin destetar, muestra una relación directa con el suministro de ácidos grasos de cadena larga preformados provenientes de la leche, que se caracteriza por poseer un alto contenido de grasa (Castellanos y Valencia 1982, Bell et al. 1987, Martínez-González et al. 2015). Los corderos de nacimiento múltiple tuvieron 61 % de la grasa cavitaria encontrada en los de nacimiento sencillo. Esta diferencia puede ser atribuida a un menor consumo de leche, y por consiguiente de grasa, en los corderos de nacimiento múltiple con respecto a los de nacimiento sencillo. En estudios futuros se deberá estudiar la influencia del tipo de nacimiento en el desarrollo metabólico y endocrino, debido a que en la etapa de crecimiento prenatal la concentración sanguínea de glucosa y aminoácidos adquieren relevancia en el metabolismo, mientras que en la etapa de crecimiento posnatal la dieta que reciben contiene un alto contenido de grasa y una menor proporción de carbohidratos (Greenwood et al. 2002, Martínez-González et al. 2015).
En corderos Suffolk Down (Pérez et al. 2002) y Segureña (Peña et al. 2005) sacrificados entre los 10 y 13 kg de peso vivo, los cortes primarios que tuvieron la mayor y menor proporción de la canal fueron la pierna y el cuello, lo que coincide con lo encontrado en el presente estudio. No obstante, el rendimiento de la pierna fue de menor magnitud. Esta diferencia puede ser atribuida a una mayor ganancia de peso predestete en los corderos de razas especializadas en la producción de carne, con respecto a los de razas de pelo. Al respecto, se ha reportado que los corderos Pelibuey tienen 66.9 % de la ganancia de peso predestete que los corderos Dorper (Hinojosa-Cuéllar et al. 2009).
En el resto de los cortes, no se realizaron comparaciones de rendimiento debido a diferencias en la definición y número de secciones corporales que comprenden los cortes primarios utilizados en los estudios que describen la composición de la canal de corderos (Peña et al. 2005, Luaces et al. 2008, Partida y Braña 2011).
En ovinos Blackbelly x Pelibuey y Pelibuey sacrificados con mayor peso al del presente estudio, se ha encontrado que el corte primario con mayor proporción en la canal es el tórax, seguido de la pierna, brazo, abdomen y cuello (Cantón et al. 1992, Partida y Martínez 2010), lo que no coincide con lo encontrado en el presente estudio. La diferencia puede atribuirse a la edad, TN, peso, raza y tipo de alimentación (Martínez et al. 1990, Luaces et al. 2007, Pérez et al. 2012). Al respecto, se sabe que el peso al sacrificio y el nivel energético de la dieta no afecta la proporción de los cortes primarios de la canal de ovinos Pelibuey finalizados (Partida y Martínez 2010). Sin embargo, es posible que en corderos con edad menor a 60 d, la proporción de los cortes primarios de la canal difiera a la de ovinos en finalización, en su orden de participación debido a que los cortes primarios de la canal tienen una velocidad de crecimiento diferente, la que está en función de la composición de hueso, músculo y grasa (Lawrence y Fowler 2002).
El rendimiento del tórax fue afectado por la interacción NP x TN. En los corderos de nacimiento múltiple provenientes de ovejas primíparas, se encontró que el tórax tuvo menor proporción en la canal con respecto a los corderos de nacimiento sencillo provenientes de primíparas y de nacimiento múltiple de ovejas multíparas. Al respecto, Luaces et al. (2007) indican que la madurez de los cortes de la canal en corderos de la raza Gallega difieren y el corte costillar tuvo madurez tardía. También, Jucá et al. (2014) encontraron que los corderos de nacimiento trillizo tuvieron un menor perímetro torácico que los de nacimiento gemelar y sencillo. Una maduración tardía de la región tórax y menor perímetro torácico pueden explicar la menor proporción del tórax en la canal en corderos de nacimiento múltiple de ovejas primíparas.
Productividad de las ovejas
Las ovejas multíparas tuvieron 118 % del peso al parto de las primíparas. Lo que coincide con lo reportado por Rajab et al. (1992), quiénes encontraron que en ovejas de las razas Morada Nova, Santa Inés y Somalí el peso al parto se incrementa con la edad. Mientras que Molina et al. (1991) reportan que las ovejas Manchega con edad mayor a tres años son más pesadas al parto que las ovejas con menor edad. La diferencia en el peso al parto entre ovejas primíparas y multíparas se relaciona con el peso a la primera concepción y el tipo de alimentación (Owens et al. 1993). Las ovejas multíparas perdieron el triple de peso durante la lactancia con respecto a las primíparas. Lo que se puede explicar, por el mayor peso vivo de las ovejas multíparas, lo que implica mayor demanda de nutrimentos para cubrir los requerimientos de mantenimiento (NRC 2007). Bajo similar manejo nutricional y capacidad productiva entre ovejas primíparas y multíparas, en las ovejas multíparas se espera un mayor uso del tejido adiposo y masa muscular para sustentar los requerimientos de nutrimentos de la producción de leche (Chávez et al. 1995).
En lo que respecta a la CC, las ovejas multíparas tuvieron un menor valor al parto con respecto a las primíparas, esta menor CC puede atribuirse a que las ovejas presentan lactancia previa, en donde es frecuente que ocurra una reducción de peso vivo y CC, cuando son alimentadas con base en el pastoreo (Godfrey et al. 1997). Es probable que durante el período de gestación de las ovejas multíparas, el consumo de nutrimentos aportados por el consumo voluntario de pasto durante el pastoreo y el complemento alimenticio asignado, un mes previo a la fecha de parto, no fue suficiente para que alcanzaran una CC similar a las primíparas.
El NP no afectó el cambio de CC, esto coincide con lo reportado en ovejas Manchegas, en donde el cambio de CC durante la lactancia fue negativo y de la misma magnitud entre ovejas con menos y más de tres años (Molina et al. 1991). La CC es una variable de fácil y de rápida medición, pero subjetiva. Sin embargo, la puntuación de CC en ovejas adultas se asocia con el contenido de lípidos en la canal, además de que es un buen indicador de las reservas de energía disponibles en las ovejas (Sanson et al. 1993). Es posible que la magnitud de los cambios detectados en la CC en 56 d, no sean suficientes para detectar diferencias significativas en su cambio entre ovejas primíparas y multíparas. En este sentido, no se encontraron estudios en ovejas en lactación que muestren la relación entre una unidad de cambio en la CC con el peso vivo. Sin embargo, en ovejas Pelibuey no lactantes y no gestantes se ha establecido que una unidad de cambio en la CC corresponde a 5.8 kg de peso vivo (Chay-Canul et al. 2011). En este estudio, las ovejas primíparas y multíparas perdieron entre 1.5 y 4.7 kg, situación que pudiera explicar que la reducción en la CC no haya sido mayor a una unidad de CC.
El peso de la camada al nacimiento y al destete no fue afectado por el NP, esto coincide parcialmente con lo indicado por Cadenas-Cruz et al. (2012) quienes encontraron que ovejas Blackbelly con uno y dos partos, tienen un peso de camada al nacimiento similar, pero, menor al de ovejas con más de dos partos. Mientras que en el caso del peso de la camada al destete, Hinojosa-Cuéllar et al. (2015) indican que las ovejas de pelo en sus primeros cuatro partos producen camadas al destete con peso similar. En contraste respecto a ovejas Blackbelly (Cadenas-Cruz et al. 2012) y Pelibuey (Magaña-Monforte et al. 2013) con más de dos partos, que producen camadas más pesadas al destete que las de primer parto.
La productividad de las ovejas fue similar entre primíparas y multíparas. Sin embargo, los valores obtenidos fueron superiores a los reportados para ovejas Blackbelly, Florida y St. Croix (Godfrey et al. 1997). Las ovejas con tipo de parto múltiple, tuvieron un mayor peso al parto que las ovejas con parto sencillo, un resultado similar, pero de menor magnitud fue reportado en ovejas Morada Nova, Santa Inés y Somalí (Rajab et al. 1992).
El TP afectó el cambio de peso y el cambio de peso diario de las ovejas. Las ovejas con parto múltiple perdieron más peso durante la lactancia que las ovejas de parto sencillo. Las diferencias en el cambio de peso entre ovejas con parto múltiple y parto sencillo pueden atribuirse a que las ovejas de parto múltiple tienen mayores requerimientos de nutrimentos, que las ovejas de parto sencillo (NRC 2007). Las ovejas de parto múltiple y sencillo estuvieron expuestas a un similar manejo nutricional, esta situación probablemente favoreció que las ovejas de parto múltiple perdieran más peso debido a un mayor uso de sus reservas corporales para apoyar la mayor producción de leche (Alexandre et al. 2001).
Las ovejas con parto sencillo y múltiple, perdieron una unidad de CC durante la lactancia, sin que se detectara influencia del TP. Este resultado difiere de lo indicado por Alexandre et al. (2001), quienes al ofrecer un alto nivel de complementación alimenticia a ovejas de pelo, permite que las ovejas con parto sencillo muestren un incremento de una unidad de CC durante la lactancia. Mientras que las ovejas con parto gemelar, muestran un incremento en su CC de 0.2 unidades. La reducción en CC detectada tanto en ovejas con parto múltiple como en las de parto sencillo, sugiere que estas no lograron cubrir sus requerimientos de nutrimentos para la etapa de lactación (NRC 2007). Las ovejas de parto múltiple tuvieron mayor productividad con respecto a las de parto sencillo, debido al mayor peso de la camada al nacimiento y al destete. La ventaja de las ovejas de parto múltiple sobre las de parto sencillo, se atribuye al mayor número de corderos en la camada. Lo que coincide con lo reportado en ovejas Morada Nova, Santa Inés y Somalí en las que la camada formada por dos corderos tienen un peso al nacimiento y al destete del 156 y 149 %, respectivamente, del peso de camadas con un solo cordero (Rajab et al. 1992). Al respecto, Hinojosa-Cuéllar et al. (2015) encontraron que el TP influye sobre el peso de la camada al destete, siendo más pesadas las camadas con dos corderos que las que tienen uno. En lo referente a la eficiencia de producción, Godfrey et al. (1997) reportan que las ovejas con parto gemelar tienen una mayor productividad que la presentada en ovejas con parto sencillo, lo que coincide con lo encontrado en el presente estudio.
Conclusiones
El tipo de parto afectó la ganancia diaria de peso predestete, peso al sacrifico y rendimiento de la canal caliente de los corderos Blackbelly x Pelibuey. Los corderos de nacimiento sencillo tuvieron un comportamiento productivo superior al de corderos de nacimiento múltiple. El número de parto de las ovejas influyó en el peso al nacimiento y área del músculo Longissimus dorsi de los corderos, los corderos provenientes de ovejas multíparas tuvieron mayor peso al nacimiento y área del músculo L. dorsi con respecto a los corderos de ovejas primíparas. El número de parto y el tipo de parto afectaron el cambio de peso de las ovejas, las ovejas multíparas y ovejas con parto múltiple tuvieron mayor pérdida de peso durante la etapa de lactancia, que las ovejas primíparas y ovejas con parto sencillo. Las ovejas con parto múltiple mostraron mayor eficiencia productivaque las de parto sencillo.

