











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El manejo óptimo del cultivo de la vid para fines enológicos requiere de experiencia en mejorar el rendimiento por hectárea con frutos de mejor calidad para asegurar la elaboración de vinos de calidad comercial (Adsule 2014). En la actualidad la industria del vino requiere para su elaboración frutos con mayores niveles de pigmentación, aroma, grados brix y antioxidantes (McCormick 2012). El uso de nuevas tecnologías en cultivos pretende enfrentar esta demanda mediante el uso de biorreguladores que faciliten su manejo (Ramírez et al. 2010). La prohexadiona de calcio (P-Ca) es un retardante de crecimiento que inhibe la biosíntesis de las giberelinas A 1, A 4 y A 7, por medio del bloqueo de las dioxigenasas, las cuales requieren al 2-oxoglutarato como co-substrato. Lo que ocasiona un menor crecimiento celular y reducción en el desarrollo vegetativo de las plantas tratadas (Rademacher 2000).
La P-Ca se ha utilizado en vid, con resultados en la reducción del tamaño del fruto y el número de racimos (Vaquero-Fernández et al. 2008), incrementando las propiedades organolépticas del fruto como las antocianinas y polifenoles (Lo Guidice et al. 2004). Los cuales son compuestos que inciden de forma directa en la calidad final del vino (Avizcuri-Inac et al. 2013). La P-Ca modifica la ruta metabólica de la síntesis de los flavonoides, lo que provoca un aumento de la síntesis de compuestos antioxidantes (Rademacher 2004, Puhl et al. 2008). El alto contenido de compuestos antioxidantes en el vino tinto es favorable, ya que se preserva por más tiempo; estos compuestos contribuyen al fortalecimiento inmunológico humano, protegiéndolo contra enfermedades coronarias, diabetes y varios tipos de cáncer; además de retrasar el envejecimiento (Okla et al. 2015). La etapa fisiológica de la planta y la dosis de aplicación de la P-Ca tienen un efecto determinante en el peso, diámetro, tamaño de fruto y el número y peso de racimos (Lo Guidice et al. 2004, Vaquero-Fernández et al. 2006); además, se ha demostrado que el uso de la P-Ca en vid modifica la ruta metabólica de la biosíntesis de flavonoides, lo que proporciona un mecanismo de defensa para las plantas (Puhl et al. 2008). En base a estas experiencias y considerando el mejoramiento en la calidad de frutos cosechados, se planteó la presente investigación con el objetivo de evaluar el efecto de la P-Ca sobre el rendimiento, la calidad y las propiedades organolépticas del fruto de vid cultivar Shiraz.
Materiales y Métodos
Sitio de Estudio
La investigación se realizó de 2014 a 2015 en los viñedos San Lorenzo ubicados en Parras de la Fuente, Coahuila, México; en los laboratorios de Horticultura de la Universidad Autónoma Agraria Antonio Narro (UAAAN) y el laboratorio de Investigación en Alimentos de la Universidad Autónoma de Coahuila (UAdeC), ubicados en Saltillo, Coahuila, México.
Las coordenadas geográficas del predio experimental son 30° 30’ 0.33’’ LN y 102° 11’ 31.9’’ LO con altitud de 1505 msnm, clima seco semiárido
y temperatura media anual de 14 a 18 ◦C; con lluvias de abril a octubre y precipitación promedio anual de 366 mm. Los vientos predominantes provienen del este todo el año. El tipo de suelo es de textura migajón-arcilloso con 50 % arcilla, 26 % limo, 24 % arena, 1.98 % materia orgánica y pH de 8.57. El rango de temperatura durante el experimento fue de 19 a 28 ◦C; con precipitación de 47 mm y humedad relativa promedio de 51 %.
Material Vegetal
El material vegetal consistió de plantas de vid del cultivar Shiraz, clon 174, portainjerto 420 A11 de 8 años de edad. La plantación fue establecida a 1.5 m entre plantas y 2.5 m entre hileras, con una densidad de 2 620 plantas ha-1; el manejo, se llevó de acuerdo a los estándares de la casa Madero.
Se seleccionaron cinco plantas por tratamiento, a las que se les aplicó en una o en dos ocasiones la prohexadiona de calcio (Apogee® con P-Ca al 27.5 %) en las siguientes dosis: 0 (agua-testigo), 100, 200 y 300 mg L-1; agregando 2 mL del surfactante pegodel® por litro de solución. El biorregulador fue asperjado la primera ocasion a punto de goteo el 11 de abril de 2014 cuando las plantas presentaron plena floración, con una aspersora manual de 10 L. La segunda aplicación con P-Ca en las mismas concentraciones se realizó 15 d después de la primera aspersion.
Variables evaluadas
Al momento de la cosecha se midió el diámetro polar y ecuatorial de 50 frutos por planta, para lo cual se tomaron los racimos localizados en la parte media con un vernier digital marca caliper® con escala de 0 a 10 cm. El p eso de los frutos se realizó con una báscula modelo Ohaus mo delo 3729 con capacidad máxima de 3000 g y resolución de 0.1 g. En cosecha, se determinó el número de racimos por planta, p eso por racimo y el rendimiento de fruto.
Para determinar el efecto de P-Ca sobre la calidad, se tomaron muestras de 50 frutos maduros por tratamiento y se congelaron en hielo seco (CO 2). Las muestras se llevaron al laboratorio de Horticultura de la UAAAN y se conservaron a temperatura de 7 °C durante 24 h. Las muestras fueron posteriormente trasladadas al laboratorio de alimentos de la UAdeC y se mantuvieron en congelamiento a una temperatura de -20 °C por un período de 48 h.
La determinación de procianidinas totales se realizó con la técnica de HCl-butanol (Porter et al. 1986). Para lo cual las muestras de uva se descongelaron, paraluego pesar 1 g en una balanza analítica digital modelo Velab, que se colocó en un tubo de ensayo con tapón de rosca, al que se le agregaron 5 mL de metanol, luego se dejó reposar 12 h y filtró en papel Whatman número 1. De los extractos obtenidos se tomó una alícuota de 250 µL, que se transfirieron a un tubo de ensayo, al que se le agregaron 1.5 mL de HCl-butanol al 5% (v/v) y 50 µL de reactivo férrico. La mezcla se calentó en un baño maría Yamato®, water Bath, BM 400, por una hora a temperatura de 90 °C, para luego tomar la lectura en un espectrofotómetro marca Biomate® 3, Thermo spectronic a una longitud de onda de 550 nm. Los datos obtenidos se expresaron como equivalentes del estándar de procianidina-1 (SigmaAldrich®). Para este estándar se elaboró una curva de calibración con las concentraciones de 0, 100, 200, 300, 400, 500 y 600 mg L-1.
El contenido de polifenoles totales se determinó por el método de Wong-Paz et al. (2014). Para lo cual se tomó del extracto metanólico una alícuota de 20 µL y se mezcló con 20 µL de reactivo de folin, se dejo reposar la mezcla por 5 min, para luego agregar 20 µL de carbonato de sodio y dejar reposar por 5 min, para luego diluir en 125 µL de agua destilada y tomar la lectura con un espectrofotómetro lector de microplaca Epok, Biotek instruments, controlado con el software paraanálisis de datos Gen 5. La máxima absorción de la longitud de onda se determinó para el rango UV-Vis del estándar de ácido gálico. Los datos graficaron para seleccionar la longitud de onda de 790 nm. Los niveles de polifenoles totales se expresaron en equivalentes del estándar de ácido gálico por gramo de peso seco de acuerdo a la curva de calibración construida con las concentraciones de 0, 100, 200, 300, 400, 500 y 600 mg L-1.
La determinación de antocianinas se realizó con el método de pH diferencial (Lee et al. 2005). La técnica se basa en la propiedad estructural que poseen estos compuestos para cambiar de acuerdo al pH en que se encuentran los pigmentos monoméricos de antocianinas, los cuales tienen un cambio reversible de coloración cuando se modifica su pH. La forma oxonio coloreada se expresa en pH 1.0, mientras que la forma hemiacetal incolora predomina en pH 4.5. La diferencia en la absorbancia de los pigmentos a una longitud de onda de 520 nm es proporcional a su concentración. La solución buffer de pH 1.0 se preparó pesando 1.86 g de cloruro de potasio (KCl 0.025 M) que se diluyó en 980 mL para lo cual se ajustó el pH (± 0.05) con HCl a 6.3 mL. Luego se pasó a un matraz y se aforó a 1.0 L con agua destilada. Para la preparación de la solución buffer de pH 4.5 se pesaron 54.43 g de acetato de sodio (CH 3CO 2Na 0.4 M) y se diluyeron hasta 980 mL ajustando el pH (± 0.05) con HCl, para luego transferir a un matraz y aforar a 1 L con agua destilada. La muestra metanólica se añadió en una proporción de 1 a 4, el factor de dilución se determinó por dilución hasta que la absorbancia a 520 nm estuvo dentro del intervalo lineal de la curva de calibración. Posteriormente se tomaron dos alícuotas de 50 µL del extracto de uva, que se colocaron en celdas, a las que a una se le agregó 50 µL de so lución pH 1.0 y a otra 50 µL de solución pH 4.5 , para luego tomar lecturas en el espectrofotómetro a 520 y 700 nm, para luego comparar con la lectura de la celda en blanco. La cantidad de antocianinas presentes en las muestras del extracto se expreso como equivalentes del estándar cianidina-3-glucósido (Sig ma-Aldrich®). Para este estándar, se elaboró una curva de calibración con las concentraciones de 0, 62.5, 125, 250, 500 y 1000 mg L-1.
El experimento se estableció bajo un diseño estadístico completamente al azar con cinco repeticiones por tratamiento, los datos se analizaron con el programa de análisis estadístico SAS 9.0, para determinar diferencias estadísticas entre medias se realizó la prueba de Tukey al 0.05 %
Resultados
La Tabla 1 muestra que el diámetro ecuatorial del fruto no fue modificado por la P-Ca; sin embargo, mientras el diámetro polar fue ligeramente menor en el tratamiento con 100 mg L-1 de P-Ca aplicado en una ocasion, este parámetro mostró un incremento del 3 % en el tratamiento P-Ca a 200 mg L-1. El peso y número de racimos; así como el peso individual de fruto no mostraron cambios estadísticos entre los tratamientos con P-Ca. Las plantas que recibieron en una ocasion el tratamiento con 300 mg L-1 de P-Ca mostraron un incremento en rendimiento del 53 % (Tabla 1).
Tabla 1 Efecto de la P-Ca sobre características del fruto, racimo y rendimiento en vid cultivar Shiraz
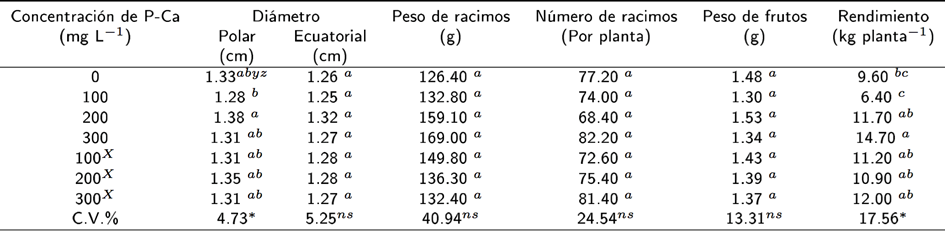
∗: Significativo y ns: no significativo a una p ≥ 0.05; C.V. : coeficiente de variación; Y: media de 50 frutos; z: valores con la misma letra dentro de cada factor en cada columna son iguales (Tukey, p ≥ 0.05); x : 2 aplicaciones.
La prohexadiona de calcio incrementó de forma significativa (p ≤ 0.05) el contenido de procianidinas totales en los frutos maduros (Figura 1). Los tratamientos con 200 y 300 mg L-1 de PCa aplicados en dos ocasiones estimularon el incremento de estos antioxidantes en un 90 y 98 %, al compararse con el testigo. La dosis de 300 mg L-1 de P-Ca aplicado una vez ocasionó un aumento del 45 % de procianidinas totales. El contenido de polifenoles totales en frutos se ilustra en la Figura 2. La prohexadiona de calcio provocó aumento significativo (p ≤ 0.05) de polifenoles totales en el fruto. El tratamiento con P-Ca a 300 mg L-1 asperjado en una ocasión tuvo un aumento del 42% de polifenoles; mientras que al aplicarse en dos ocasiones, el incremento fue del 21%.
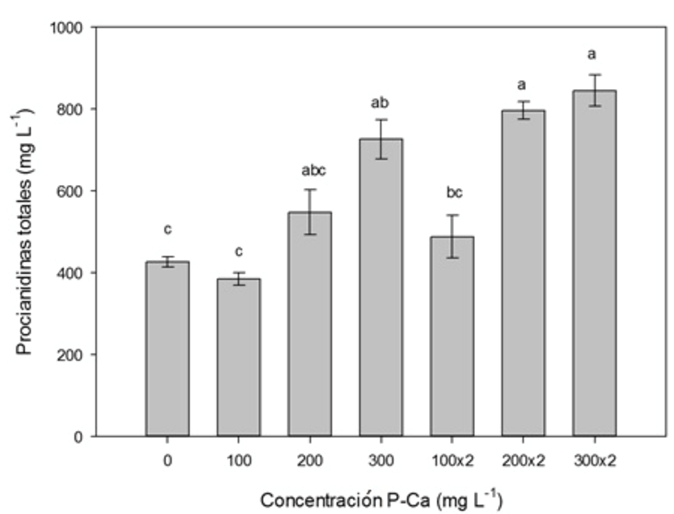
Figura 1 Efecto de P-Ca sobre el contenido de procianidinas totales (mg) en el fruto maduro de uva cultivar Shiraz. Cada barra representa la media de diez repeticiones (± desviación estándar). Letras diferentes indican diferencias estadísticas (Tukey, p ≤ 0.05).
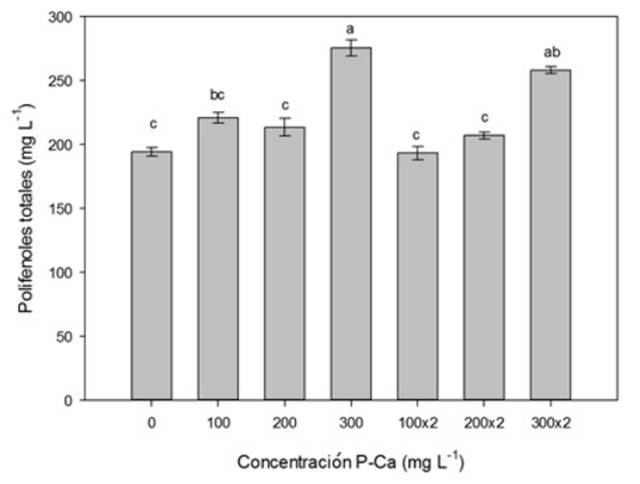
Figura 2 Efecto de P-Ca sobre el contenido de polifenoles totales (mg) en el fruto maduro de uva cultivar Shiraz. Cada barra representa la media de diez repeticiones (± desviación estándar). Letras diferentes indican diferencias estadísticas (Tukey, p ≤ 0.05).
En la Figura 3 se observa el efecto de P-Ca en el contenido de antocianinas totales en los frutos. El tratamiento con P-Ca a 200 mg L-1 en dos aplicaciones incrementó de forma significativa (p ≤ 0.05) el contenido de antocianinas, que fue un 67 % mayor al contenido de los frutos del tratamiento testigo. Los tratamientos con P-Ca a 300 mg L-1 asperjado en una ocasión y 100 mg L-1 en dos aplicaciones tuvieron un 32 % de incremento en el contenido de antioxidantes.

Discusión
La aplicación de prohexadiona de calcio en hortalizas con frecuencia reduce el crecimiento o tamaño del fruto (Rademacher 2000, Ramírez et al. 2010). Lo que se explica por la reducción de células (Ramírez et al. 2009). En el presente estudio la P-Ca no ocasionó reducción del número y peso de racimos, y del peso y tamaño de frutos de uva (Tabla 1). Estos efectos son de gran importancia hortícola ya que al no afectarse estas variables se tiene un fruto con características fenotípicas competitivas en el mercado (McCormick 2012). Estas particularidades fenotípicas han sido observadas en manzano (Costa et al. 2004) y en vid (Giacomelli et al. 2013). La etapa fenológica en que la P-Ca se aplica influye en el tamaño del fruto. Al respecto Giacomelli et al. (2013) reportan que durante la antesis ocurre una actividad elevada en la síntesis de las gibrerelinas A 1 y A 4 de la vid. Se sabe que la P-Ca inhibe la síntesis de estas hormonas (Rademacher 2004); además, cuando se aplica en vid en prefloración, el resultado es la formación de frutos pequeños (Lo Guidice et al. 2003, Lo Guidice et al. 2004) y racimos cortos (Kok et al. 2013). La P-Ca asperada cuando las plantas están en floración, bloquear la sintesís de giberelinas lo que resulta en una reducción en el tamaño y peso del fruto. El rendimiento por planta se incrementó en la mayoría de los tratamientos con P-Ca (Tabla 1). Este efecto es consistente con los reportes en vid (Lo Guidice et al. 2004), manzano (Costa et al. 2004), chile (Ramírez et al. 2010) y to mate (Ramírez et al. 2008). El rendimiento se basa en el número de ores que la planta produce (Vasconcelos et al. 2009), pero la P-Ca se caracteriza por estimular la oración (Rademacher 2004). Este incremento se relaciona con la disminución de las giberelinas biológicamente activas y al aumento de citocininas en el tejido primordio, por lo que se aumenta la formación de yemas orales (Ramirez et al. 2005). Lo cual probablemente ocurrio con la aplicación de P-Ca en el cultivar de uva Shiraz. La P-Ca una vez en el tejido, modica la dirección de translocación de asimilados (Rademacher 2000); condición que provoca que más fotosintatos y citocininas se trasladen a los frutos en desarrollo (Costa et al. 2004, Ramírez et al. 2005) lo que se evita un incremento en la caída y la reducción del desarrollo de frutos (Tabla 1).
La aplicación de P-Ca incremento los niveles de antioxidantes en los frutos maduros del cultivar Shiraz. El contenido de procianidinas (Figura 1), polifenoles totales (Figura 2) y antocianinas totales (Figura 3) aumentó de forma significativa (p ≤ 0.05) con las diferentes concentraciones de PCa. En la elaboración de vinos, los tres antioxidantes referidos contribuyen a producir un vino de mejor calidad (Lee et al. 2005). Las procianidinas proporcionan astringencia en los vinos, por lo que su contenido en los frutos influye de forma directa con la calidad final del vino (Vaquero-Fernández et al. 2006). Al respecto Kok y Bal (2014) reportan en el vid cultivar Gewürztraminer incrementos de monoterpenos en frutos cuando se aplicó P-Ca en dosis de 200 y 300 mg L-1.
Incrementos de polifenoles totales por la PCa (Tabla 1), han sido reportados por Lo Guidice et al. (2004) y Kok et al. (2013) al aplicar en la vid la concentración de 250 mg L−1 en prefloración. Mientras que Vaquero-Fernández et al. (2006) encontraron aumento de polifenoles totales en el vino con la aplicación de P-Ca en prefloración. Mientras que Avizcuri-Inac et al. (2013) reportan que la aplicación de P-Ca en las variedades Tempranillo y Grenache incrementó el contenido de polifenoles del vino. Este efecto también fue observado por Lo Guidice et al. (2004) en la variedad Cabernet Franc cuando las plantas se asperjaron con P-Ca a 250 mg L−1. En el presente trabajo los tratamientos con P-Ca a 100, 200 y 300 mg L−1 en dos aplicaciones estimularon un aumento significativo (p ≤ 0.05) en el contenido de antocianinas totales en los frutos (Figura 3). Es evidente que la presencia de antocianinas en uvas contribuye en la calidad final del vino (Kok et al. 2013, Vaquero-Fernández et al. 2006). El contenido de antocianinas en el cultivo de la vid se encuentra determinado por factores como la temperatura (Yamane et al. 2006), las prácticas culturales como el aclareo de racimos y las podas (Kok et al. 2013) y por la radiación solar que recibe el fruto (Costa et al. 2004). La P-Ca modifica la ruta metabólica de la síntesis de flavonoides mediante el bloqueo del 2-oxo-glutarato, lo que incrementó la síntesis de los compuestos antocianos (Rademacher 2000), los cuales influyen en la estabilidad de la coloración del vino (Lo Guidice et al. 2004). Al respecto Kok et al. (2013) aumentaron la cantidad de antocianinas en vid cuando aplicaron 250 mg L-1 de P-Ca en prefloración, pero los niveles de antocianinas fueron similarea a los que se obtienen cuando se realizan podas y aclareo de racimos. Mientras que Avizcuri-Inac et al. (2013) y Vaquero-Fernández et al. (2006) incrementaron el contenido de antocianinas en el vino de las variedades Tempranillo y Granache cuando aplicaron prohexadiona de calcio en prefloración. El mecanismo a través del cual la P-Ca estimula la producción de estos antioxidantes no se conoce en detalle, pero Rademacher (2000) ha propuesto que P-Ca inuye de forma directa en la síntesis de flavonoides y enzimas ligadas con su metabolismo. Lo que incrementa la producción de antioxidantes específicos como vitamina C en manzano y pera (Rademacher 2004), licopeno en tomate (Ramírez et al. 2008) y capsaicina en chile jalapeño (Ramírez et al. 2009).
Conclusiones
Con los resultados obtenidos es posible considerar el uso de P-Ca como una alternativa para incrementar en el cultivar de vid Shiraz los niveles de antioxidantes procianidinas, polifenoles y antocianinas. La prohexadiona de calcio aplicada en floración y quince días después al cultivar de vid Shiraz no altera el tamaño y peso del fruto, peso y número de racimos, pero si incrementa el rendimiento y contenido de procianidinas, polifenoles y antocianinas totales en frutos maduros.

