











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La apicultura en México es una actividad pecuaria de gran importancia económica, social y ecológica, que genera importantes divisas para el país y beneficia a 40 000 apicultores (Guzmán-Novoa et al. 2011, SAGARPA 2013). Sin embargo, la presencia de abejas africanizadas Apis mellifera L. (Hymenoptera: Apidae) es uno de los principales problemas que enfrenta la industria apícola mexicana. Entre los principales efectos indeseables de este morfotipo de abeja están su comportamiento altamente defensivo y migratorio, su tendencia a abandonar o evadirse de las colmenas, lo que disminuye la producción de miel (Uribe et al. 2003).
Las abejas africanizadas se detectaron por primera vez en el sureste de México a finales de 1986 (Fierro et al. 1988), a partir de entonces se han dispersado por todo el país; se estima que se encuentran establecidas en más del 95 % de las regiones apícolas de México (Guzmán-Novoa et al. 2011). En julio de 1993 se confirmó la presencia de abejas africanizadas en la ciudad de Mexicali, Baja California (Guzmán-Novoa y Page 1994), y desde 1994 se establecieron en el sur de California (Lazaneo 2002). Se sabe que las variaciones ambientales influyen en el grado de africanización de las poblaciones de abejas, siendo las condiciones tropicales las más favorables para la introgresión de genes africanos (Medina-Flores et al. 2014). Baja California se ubica en el subtrópico mexicano con variaciones ambientales que pueden reflejarse en distintas proporciones de morfotipos africanizados y europeos en las poblaciones de Apis mellifera. En este estado, la africanización de las abejas ha sido poco estudiada, por lo que no se conoce el grado de africanización de las poblaciones de abejas, lo cual es fundamental para evaluar el impacto de este fenómeno biológico sobre la apicultura local y para el diseño de estrategias que permitan reducir los problemas asociados con las abejas africanizadas. La técnica más precisa para discriminar entre abejas europeas y africanizadas es el análisis de ADN mitocondrial. Sin embargo, los análisis morfométricos siguen siendo una forma barata y eficiente para evaluar el grado de africanización de las abejas, aun cuando estas contengan mitocondrias de origen europeo, lo que ocurre cuando zánganos africanizados se aparean con reinas europeas (Kono y Kohn 2015). Por lo anterior, el objetivo de esta investigación fue comparar la frecuencia de morfotipos africanizados y europeos de las poblaciones de abejas del Valle de Mexicali y Ensenada, Baja California mediante análisis morfométrico.
Materiales y métodos
El estudio se realizó en la región noroeste del estado de Baja California, en los municipios de Ensenada y Mexicali, al norte del paralelo 30° (Figura 1), donde las condiciones climáticas favorecen la práctica de la apicultura. Los climas predominantes son árido, muy seco, cálido y con lluvias en invierno en el Valle de Mexicali, y seco mediterráneo, templado y con lluvias invernales en Ensenada (García 1988, INEGI 2001). Los apiarios se ubican en las zonas agrícolas, se mu est rea ron 47 colonias en el Valle de Mexicali y 34 en Ensenada. Además se incluyeron 11 enjambres silvestres capturados en Mexicali y cuatro en Ensenada. Se tomaron 30 abejas de cada colonia o enjambre y se depositaron en frascos de plástico con 150 mL de alcohol al 70 %. El montaje y análisis de las muestras se realizó en el laboratorio del Departamento de Apicultura de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Autónoma de Yucatán. Se seleccionaron al azar 10 abejas obreras de cada muestra, de cada abeja se diseccionó el ala delantera y la pata posterior derecha, las cuales se colocaron en láminas para transparencias de 24 X 36 mm. Para medir los caracteres morfométricos de las abejas se utilizó un equipo de m orfo m et ría computarizada con microscopio invertido y fuente de luz incorporada (Inbumex). Las transparencias se montaron con el objetivo 10x del microscopio en una tableta digitalizadora SummaSketch II Professional Plus conectada a una computadora, y se marcaron puntos en la imagen del carácter morfométrico dentro de la tableta. Estos puntos se emplearon para calcular en milímetros (mm) la longitud del ala y del fémur de cada muestra, utilizando el programa AFUSDA7 (Pérez-Castro et al. 2002).
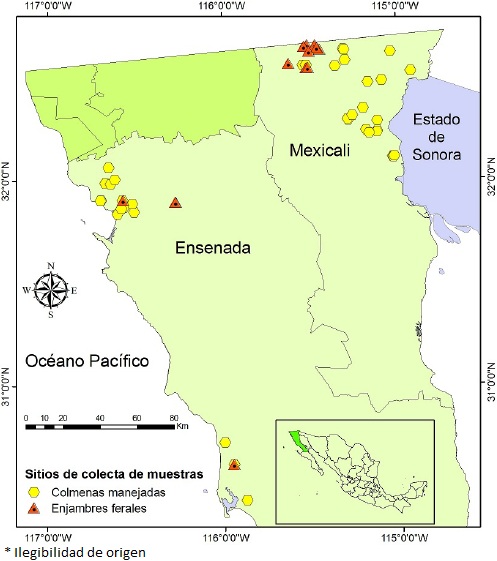
Figura 1 Ubicación de colmenas y enjambres muestreados en los municipios de Ensenada y Mexicali, Baja California.
Con los valores promedio de longitud del ala y del fémur de cada colonia se calculó la probabilidad de africanización, usando el coeficiente del procedimiento discriminatorio FABIS (Sylvester y Rinderer 1987). Las colonias con coeficiente igual o menor de -0.56 fueron consideradas africanizadas con una probabilidad de 0.99, en caso contrario se consideraron europeas. Las frecuencias de los morfotipos africanizados y europeos en las colonias manejadas y enjambres, se analizaron mediante una prueba de Chi-cuadrada con el procedimiento PROC FREQ de SAS (SAS 2001), para determinar si existen diferencias entre estas, en las localidades estudiadas. Las medias de la longitud de los caracteres morfométricos de las colonias africanizadas y europeas se compararon entre localidades mediante la prueba de t-Student con el procedimiento PROC TTEST de SAS a una significancia del 0.05 (SAS 2001).
Resultados y discusión
Los resultados de la técnica morfométrica FABIS indican que 91.49 % de las colonias de abejas de Mexicali y 67.65 % de las colonias de Ensenada tienen morfotipos africanizados (Tabla 1). En tanto que en los enjambres silvestres se encontró que 100 % en Mexicali y 50 % en Ensenada tienen morfotipos africanizados (Tabla 1). Los altos porcentajes de africanización difieren de los encontrados por Zamora et al. (2008), quienes detectaron 44 % de morfotipos y 50 % de m itoti pos de colonias africanizadas en el estado de Baja California; el mayor porcentaje de morfotipos africanizados encontrados puede deberse a que se ha incrementado el proceso de introgresión de genes africanizados en las poblaciones de abejas silvestres y manejadas en la entidad. En el caso de las muestras de Ensenada la proporción de morfotipos africanizados detectados en este estudio es similar a la reportada por Kono y Kohn (2015) quienes encontraron 61 % de morfotipos y 65 % de mitotipos africanizados.

Frecuencia de morfotipo en la misma columna seguida por la misma letra son estadísticamente iguales según la *Prueba Chi-cuadrada (X2=6.0606, gl=1, p=0.0138), ** Prueba Chi-cuadrada (X2=3.2667, gl=1, p=0.0707), *** Prueba Chi-cuadrada (X2=6.2308, gl=1, p=0.0126).
Tabla 1 Tratamientos en el primer y segundo ciclo de embriogénesis somática secundaria.
La prueba de Chi-cuadrada indicó que las frecuencias de los morfotipos africanizados en colonias manejadas no es independiente entre las localidades (X2 =6.0606, gl = 1, p = 0.0138), la aparición de estos morfotipos depende de la localidad donde se efectué el mu est reo, presentándose mayor frecuencia de morfotipos africanizados en Mexicali. Por el contrario, la frecuencia de los morfotipos europeos en colonias manejadas es independiente entre localidades (X2 = 3.2667, gl = 1, p = 0.0707), indicando que la aparición de estos morfotipos no depende de la localidad (Tabla 1). Las frecuencias de los morfotipos africanizados en enjambres silvestres no fue independiente entre las localidades (X2 = 6.2308, gl = 1, p = 0.0126), presentándose mayor frecuencia de morfotipos africanizados en Mexicali (Tabla 1). Sin embargo el tamaño de muestra en Ensenada es más pequeño en comparación con Mexicali, por lo que las diferencias podrían no ser precisas. Este resultado contrasta con lo reportado por Kono y Khon (2015) quienes registraron 70 % de morfotipos africanizados en diez muestras de colonias silvestres en San Diego, California.
La longitud promedio del fémur de las colonias africanizadas en los municipios de Ensenada y Mexicali no presentó diferencias significativas. Sin embargo, las longitudes del ala anterior de las abejas africanizadas de Ensenada fueron significativamente mayores en comparación con las de Mexicali (Tabla 2). La longitud promedio del fémur y del ala anterior de las colonias europeas en los municipios de Ensenada y Mexicali no presentó diferencias significativas. Tanto la menor proporción de morfotipos africanizados (Tabla 1), como la mayor longitud de alas de las poblaciones de abejas de Ensenada (Tabla 2), podrían indicar que en la zona costa las abejas africanizadas se han introducido de forma lenta. Al respecto Quezada-Euán (2007) menciona que el movimiento de los enjambres africanizados fue más lento en la costa del Océano Pacífico en comparación con la costa del Golfo de México, lo que pudo estar influenciado por la altitud, y los patrones de lluvia y disponibilidad de alimento, además en la migración los enjambres africanizados se vieron obstaculizados por los sistemas montañosos, lo que podría explicar los diferentes porcentajes de africanización encontrados en los municipios de Ensenada y Mexicali, ya que esta última se encuentra bordeada por sistemas montañosos de la Sierra de Juárez y San Pedro Mártir. También las condiciones ambientales pudieron influir en los diferentes porcentajes de africanización observados entre estos dos municipios. Al respecto Taylor y Spivak (1984) mencionan que las abejas africanizadas están preadaptadas para invadir y persistir en los hábitats tropicales, mientras que las temperaturas frías limitan su progreso en los climas templados. La franja de clima mediterráneo en Ensenada podría ofrecer condiciones favorables para los morfotipos europeos, por lo contrario, el clima cálido y muy seco del municipio de Mexicali puede favorecer más a los morfotipos africanizados, lo que explicaría las diferencias en las proporciones de morfotipos africanizados y europeos encontrados. Los resultados obtenidos coinciden con lo encontrado por Medina-Flores et al. (2014), quienes reportan 87 % de morfotipos africanizados en una región de clima subtropical, contra 38 % de morfotipos africanizados en una región de clima templado húmedo.

Promedios de carácter morfométrico en la misma columna seguidos por la misma letra no son diferentes estadísticamente (PROC TTEST, t-Student, P > 0.05). Los valores entre paréntesis indican los intervalos de confianza del valor de la media.
Tabla 2 Tratamientos en el primer y segundo ciclo de embriogénesis somática secundaria.
La velocidad de africanización en apiarios del municipio de Mexicali también se ha visto favorecida por la frecuente incorporación de los enjambres silvestres a los apiarios sin la eliminación de la reina, lo que puede acelerar el proceso de africanización (Smith 1991) a través del flujo genético de las poblaciones silvestres altamente africanizadas a las poblaciones domesticadas (Quezada-Euán y Medina 1998). Lo que coincide con los resultados obtenidos en este estudio, donde 100 % de los enjambres silvestres capturados en Mexicali presentaron morfotipos africanizados (Tabla 1).
Este estudio demostró que la proporción de africanización de colonias de Apis mellifera en el Valle de Mexicali, Baja California es alta, en comparación con Ensenada, lo que puede estar influenciado por las condiciones geográficas, climáticas y las prácticas de manejo. Por lo que es necesario establecer estrategias para reducir el efecto negativo de la africanización en la apicultura de Baja California, como la práctica de sustituir la abeja reina en los apiarios, por reinas seleccionadas de origen europeo.

