











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En México la comercialización de cortes primarios y/o canales provenientes de corderos jóvenes, menores a tres meses, de razas de pelo no es usual. Sin embargo, este tipo de productos cárnicos representan una opción para diversificar los productos ovinos factibles de comercializar (Sañudo et al. 1998). Como ocurre en la Unión Europea, donde existe demanda de cordero lechal debido a que su carne posee características organolépticas que permiten apreciarla desde el punto de vista gastronómico (Miguélez et al. 2007).
En los países de la región mediterránea de la Unión Europea se producen y sacrifican corderos menores a 90 d, lo que permiten obtener una canal clasificada como ligera, con categorías según el peso de A: menor a 7kg; B: de 7.1 a 10kg y C:de10.1 a 13 kg, este tipo de canal representa un producto importante en el mercado de la carne ovina (Sañudo et al. 1998, Miguélez et al. 2007). Sin embargo, los estudios efectuados alrededor de la composición de la canal en corderos menores de 45 d y ovinos destetados antes de los 90 d, indican que este tipo de canales muestran características distintivas con respecto a las de corderos de mayor edad (Miguélez et al. 2007). De ahí la importancia de realizar estudios sobre la composición de la canal de corderos, los cuales representan un importante recurso genético para producir carne en el trópico (Quiroz y Oliva 2013).
El estudio de los factores que afectan el peso y rendimiento de la composición tisular de la canal, es importante por la posible relación del peso vivo (PV) con la cantidad de productos vendibles (Borton et al. 2005). Otros estudios con corderos lechales de razas de lana, reportan la influencia de la raza, peso y sexo en la composición tisular de la canal y de diversos cortes primarios (Miguélez et al. 2007, Luaces et al. 2008, Pérez et al. 2012). Sin embargo, no se ha evaluado la influencia del tipo de nacimiento, ya que se han utilizado corderos de nacimientos sencillo. En el caso de corderos jóvenes de razas de pelo, existe poca información sobre la composición tisular de la canal (Combellas 1977), donde no se considera la participación del tipo de nacimiento del cordero, factor que puede ser importante, debido a que las ovejas de pelo muestran una alta prolificidad (Macedo y Arredondo 2008). Con base en estos antecedentes, el objetivo del presente estudio fue evaluar la influencia del tipo de nacimiento de los corderos y el número de parto de la oveja sobre la composición tisular de cortes primarios y de la canal. También, se determinó la relación de la composición tisular de los cortes primarios con el peso y rendimiento de tejido blando y hueso en la canal.
Materiales y métodos
El estudio se realizó en la unidad ovina experimental del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, localizada en el municipio de Huimanguillo, Tabasco, México (17° 51' LN, 93° 23' LO), el clima es caliente y húmedo con lluvias todo el año Af (m) (García 2004). La media anual de temperatura y humedad relativa es de 25.7 °C y 85.3 %, respectivamente (CONAGUA 2013). Se utilizaron 28 corderos Blackbelly x Pelibuey, 13 hembras y 15 machos, con edad de 56 d y peso promedio de 10.6 ± 0.4 kg. Se usó un diseño experimental completamente al azar con dos factores, los cuales fueron el tipo de nacimiento (TN) de los corderos (sencillo y gemelar), el número de parto (NP) de las ovejas (primípara y multípara) y la interacción TN x NP.
Manejo de la oveja durante la lactancia
En la primera semana de lactación, las ovejas estuvieron estabuladas con sus camadas y se alimentaron con heno de C. plectostachyus. En la segunda semana de lactación, las ovejas salieron a pastoreo de 8 a 14 h y el resto del día estuvieron con sus camadas, recibiendo heno de C. plectostachyus. A partir de la tercera semana y hasta finalizar la etapa de lactación se incrementó el número de horas de pastoreo en las ovejas de 8 a 18 h en praderas con C. plectostachyus. En promedio, el período de ocupación fue de 3 d en cada pradera y 25 d de descanso. Durante toda la etapa de lactación las ovejas se suplementaron con 500 g oveja−1 d−1 de un alimento con 15 % PC.
Manejo de los corderos durante la etapa de lactación
Con el fin de evitar que los corderos lactantes se infectaran con nematodos gastrointestinales durante el pastoreo, estos permanecieron estabulados durante toda la etapa de lactación y se alimentaron con amamantamiento controlado, a partir del quinto día de edad se utilizó un complemento alimenticio de tipo comercial con 18 % de PC, además de C. plectostachyus y Gliricidia sepium a libre acceso; restringuiendo el amamantamiento de 8 a 14 h en la segunda semana de vida de los corderos. A partir de la tercera semana y hasta concluir la lactancia, se encerraron corderos y madres después de las 18 h.
Estudio de la canal
Los corderos se sacrificaron a los 56 d de edad, con un ayuno de alimento no lácteo de 14 h. Posteriormente, se procedió a retirar la piel, cabeza, las extremidades delanteras y las traseras, vísceras abdominales y torácicas. Para registrar el peso de la canal caliente (Hernández-Espinoza et al. 2012).
La canal se dividió en cuello, brazo, tórax, abdomen y pierna; de acuerdo a la metodología de Martínez et al. (1987). Cada uno de los cortes primarios se disecó, separando y pesando el tejido blando (TB) y hueso (Cantón et al. 1992), el TB incluyó músculo, ligamentos y grasa de cobertura. El peso del TB y hueso se determinó en cada uno de los cinco cortes primarios, el peso del brazo y pierna incluyó las dos extremidades. Una vez efectuado este procedimiento se pesó el total de TB y hueso, el rendimiento de TB (%), consistió en la relación: peso del TB en el corte primario o canal completa entre el peso del corte primario o canal completa por 100. El rendimiento de huesos (%) se calculó con la relación: peso de los huesos en el corte primario o canal completa entre el peso del corte primario o canal completa por 100 (Cantón et al. 1992).
Análisis estadístico
Todos los análisis se efectuaron utilizando el paquete estadístico SAS (SAS 2002). A los datos se les aplicó la prueba de Shapiro-Wilks para probar que tuvieran una distribución normal. Para el análisis de los datos de los corderos se utilizó un modelo de efectos fijos, en donde los factores fueron TN y NP. Las variables peso al sacrificio, peso de los huesos en canal, peso del hueso en cuello, brazo, tórax y pierna, peso del tejido blando en tórax, rendimiento del hueso en cuello, rendimiento del tejido blando en abdomen y pierna, y las relaciones tejido blando: hueso en brazo, abdomen y pierna no tuvieron una distribución normal por lo que se analizaron con el test de la suma de rangos de Wilcoxon para datos no pareados (Milton 2007).
Para establecer la relación entre el peso del TB y hueso en cada uno de los cortes primarios de la canal con el peso y rendimiento de TB, y el hueso de la canal completa, se utilizó el procedimiento de correlación de Spearman. Debido a que el TN afecto (p < 0.05) el peso del TB y el hueso de la canal, los análisis de correlación se realizaron considerando la influencia del TN. Se utilizaron los valores críticos del coeficiente de Spearman para establecer si los diferentes pares de variables muestran asociación significativa (Weathington et al. 2012).
Para explicar la variación en el peso y rendimiento del TB, y hueso en la canal completa de los corderos con base en el peso del TB, y hueso en cada corte primario, se utilizó el procedimiento de regresión múltiple con la opción stepwise, a una probabilidad de p < 0.05. Los análisis de regresión se realizaron considerando la influencia del TN. La variable dependiente fue el peso y rendimiento del TB, y hueso en la canal caliente y las variables independientes el peso del TB y hueso en los cinco cortes primarios. A partir de la matriz de correlaciones se seleccionaron como variables independientes aquellas que mostraron el coeficiente de correlación (r) significativamente distinto de cero, a un nivel de significancia de p < 0.05, y que no mostraran relación significativa entre ellas, para evitar la colinealidad entre las variables independientes (López 1998).
La ecuación de predicción propuesta fue Y= β0 + β1X1+ βkXk. En donde: Y= es el valor esperado de la variable dependiente (peso y rendimiento del TB en la canal completa), β0 = el intercepto con Y, β1, ... βk = es la pendiente de la regresión de Y respecto de X1 para valores fijos de los otros X, y X1, ... Xk = es el valor de la variable independiente.
Resultados
El NP y la interacción NP x TN no afectaron las variables estudiadas (p > 0.05). El TN influyó en el peso de la canal, el peso del TB y huesos en la canal de los corderos (p < 0.05). Los corderos de nacimiento sencillo mostraron un mayor peso de la canal, así como del TB y huesos en la canal con relación a los de nacimiento gemelar (Tabla 1). El TN no influyó en el peso al sacrificio, rendimiento de la canal, relación TB:hueso, rendimiento del TB y huesos en la canal (Tabla 1), el peso, rendimiento de TB, hueso en cuello, brazo y abdomen (Tabla 2). El peso del hueso en tórax y del TB en pierna fueron afectados por el TN (p < 0.05). El peso del hueso en tórax y TB en pierna fue menor en los corderos de nacimiento gemelar con relación a los corderos de nacimiento sencillo. El peso y rendimiento del TB en tórax y el peso, y rendimiento del hueso en pierna no fueron influidos por el TN (p > 0.05). La relación TB:hueso en los cinco cortes primarios estudiados no fue afectado por el TN (Tabla 2) (p > 0.05).
Tabla 1. Influencia del tipo de nacimiento sobre la composición tisular de la canal de corderos lactantes Blackbelly x Pelibuey.
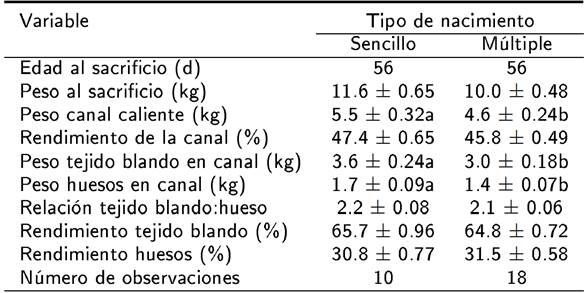
a, b medias de cuadrados mínimos ± error estándar con diferente superíndice dentro de la misma fila indican diferencia significativa (p < 0.05).
Tabla 2. Influencia del tipo de nacimiento sobre la composición tisular en cortes primarios de corderos lactantes Blackbelly x Pelibuey.
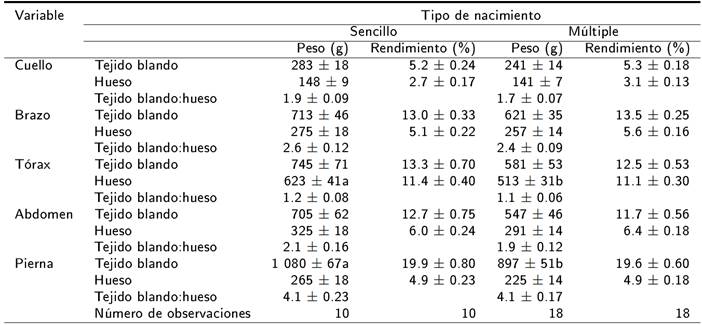
a, b medias de cuadrados mínimos ± error estándar con diferente superíndice dentro de la misma fila indican diferencia significativa (p < 0.05).
En corderos de nacimiento sencillo, los pesos del TB en brazo, tórax, abdomen y pierna mostraron relación positiva con el peso del TB en canal (p < 0.01). Mientras que el peso del TB y hueso en brazo, tórax, abdomen y hueso en pierna tuvieron una asociación positiva con el peso del hueso en canal (p < 0.01). A diferencia del peso del TB y hueso en canal, el rendimiento del TB y hueso en canal no mostraron relación con el peso del TB y hueso en los cortes primarios motivo de la investigación (p > 0.01) (Tabla 3).
Tabla 3. Matriz de correlaciones entre tejido blando y hueso en los cortes primarios y la composición tisular de la canal de corderos lactantes Blackbelly x Pelibuey provenientes de nacimiento sencillo.
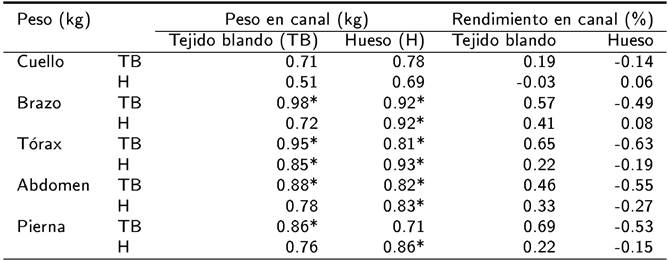
* p < 0.01.
Con excepción del hueso en el cuello, el resto de las variables mostraron relación positiva con el peso del TB en la canal de corderos gemelos (p < 0.01). El peso del hueso en la canal de corderos gemelos tuvo asociación positiva con el peso del TB y hueso en los cortes primarios (p < 0.01), con excepción del TB en tórax (Tabla 4). El rendimiento del TB en canal se correlacionó con todas las partes de la canal (p < 0.01), excepto con el brazo que no se correlacionó (Tabla 4). El rendimiento del hueso en la canal, no se correlacionó (p > 0.01) con ninguna de las variables estudiadas en los cortes primarios.
Tabla 4. Matriz de correlaciones entre tejido blando y hueso en los cortes primarios y la composición tisular de la canal de corderos lactantes Blackbelly x Pelibuey provenientes de nacimiento gemelar.
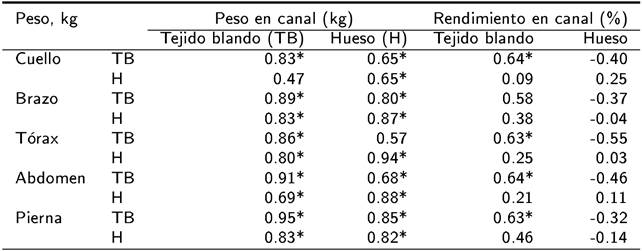
* p < 0.01.
En la tabla 5 se muestran las ecuaciones para predecir el peso y rendimiento del TB y hueso en la canal de corderos de nacimiento sencillo y gemelar. En corderos de nacimiento sencillo, la ecuación para predecir el peso del TB en canal consideró el TB en brazo (R2 = 0.96, p < 0.01), mientras que el peso del hueso en canal se estimó con el peso del hueso en brazo (R2 = 0.84, p < 0.01). En corderos de nacimiento gemelar, el peso de TB en canal se estimó con el peso del TB en pierna (R2 = 0.90, p < 0.05) y el peso del hueso en canal con el peso del TB en tórax (R2 = 0.89, p < 0.01). En corderos de nacimiento sencillo, el rendimiento del TB en canal se estableció con el peso del TB en pierna (R2 = 0.47, p < 0.05), mientras que el rendimiento del hueso en canal se estimó con el peso del TB en tórax (R2 = 0.40, p < 0.05). En corderos de nacimiento gemelar, las ecuaciones para predecir el rendimiento de TB y hueso en canal consideran el peso del TB en cuello (R2 =0.41, p < 0.01) y el peso del TB en tórax (R2 = 0.30, p < 0.05) respectivamente.
Tabla 5. Ecuaciones de predicción de la composición tisular de la canal completa a partir del tejido blando (TB) y hueso (H) en cortes primarios en corderos lactantes Blackbelly x Pelibuey de nacimiento sencillo y gemelar.
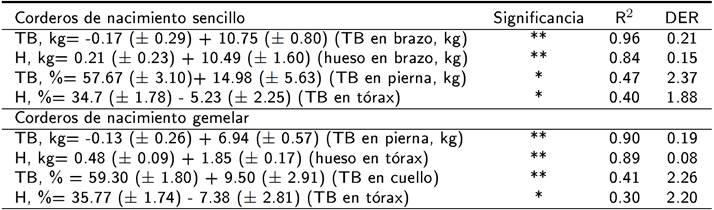
R2= coe_ciente de determinación; DER= desviación estándar residual; * p < 0.05; ** p <0.01.
Discusión
Composición tisular de la canal
El peso individual de la canal de los corderos de nacimiento gemelar representó el 83.7 % del peso de los corderos provenientes de nacimiento sencillo, resultado que es menor a menor a lo reportado en ovinos de lana (89.8 %) sacrificados en edad promedio de 142 d con peso promedio de la canal de 16 kg (McCoard et al. 2010). Los corderos de nacimiento gemelar tuvieron un menor peso de la canal, del TB y del hueso en canal; con respecto a los de nacimiento sencillo. Al respecto, en los fetos gemelos de ovinos se ha registrado un menor peso de su masa muscular con relación a los fetos de gestación sencilla (Sales et al. 2013), posiblemente una masa muscular con menor peso, explique el menor desarrollo muscular posnatal en corderos gemelos con respecto a corderos de parto sencillo. Estas diferencias pueden atribuirse al menor tamaño corporal de los corderos de nacimiento gemelar. Al respecto, Hurtado et al. (2003) indican que el perímetro torácico, altura a la cruz y la longitud del cuerpo de los corderos de nacimiento gemelar de la razas West African y Barbados Barriga Negra representa el 88.8, 93.4 y 92.8 %, respectivamente, del registrado en corderos de nacimiento sencillo.
El músculo, hueso, grasa total y otros tejidos representaron el 53.2, 24.2, 16.8 y 6.3 % de la composición tisular de la canal en corderos Manchegos lactantes (Díaz et al. 2003). Mientras que en corderos West-African de 71 d de edad, el músculo, hueso y grasa en la canal representaron el 59.3, 21.3 y 19.4 % de la canal, respectivamente (Combellas 1997). En el presente estudio, el TB incluyó tejido muscular, ligamentos, y grasa de cobertura, situación que implica considerar ciertos límites al contrastar los valores de rendimiento del TB del presente estudio, con los de rendimiento de músculo obtenido en estudios efectuados en la canal de corderos lactantes de razas de lana.
El rendimiento de TB en la canal fue superior al reportado para músculo en corderos West-African (Combellas 1997), tipo Terrincho-PDO (Santos et al. 2008) y Manchegos (Díaz et al. 2003). Sin embargo, al adicionar al valor de rendimiento en músculo, los valores de rendimiento en grasa y otros tejidos reportados en corderos West-African, tipo Terrincho-PDO y Manchegos, respectivamente, se generó una estimación del rendimiento en TB en la canal, las cuales fueron de 78.7, 78.1 y 76.3 %, respectivamente. Estas estimaciones de rendimiento de TB son superiores a las obtenidas en el presente estudio.
Con respecto al hueso en canal y la relación TB:hueso en canal, los corderos presentaron un mayor rendimiento en hueso y una menor relación TB:hueso en canal, con relación a los corderos West-African, tipo Terrincho-PDO y Manchegos. El rendimiento hueso en canal y la estimación de la relación TB:hueso en canal en corderos West-African, tipo Terrincho-PDO y Manchegos, fue de 21.3 y 3.7, 21.0 y 3.6, y 24.2 y 3.1 %, respectivamente.
Los corderos Blackbelly x Pelibuey tuvieron durante la lactancia una menor capacidad para depositar masa muscular, pero mayor para el hueso en canal, con relación a lo reportado por Combellas (1997), Santos et al. (2008) y Díaz et al. (2003). Es posible, que estas diferencias son atribuidas a la mayor ganancia de peso pre-destete de los corderos West-African (Combellas 1997) y Manchegos (Vergara y Gallego 1999) con respecto a la reportada en los corderos Pelibuey x Blackbelly (Ríos-Utrera et al. 2014). El rendimiento en TB y huesos en cuello, brazo y abdomen fue similar entre corderos de nacimiento sencillo y gemelar. Al respecto los estudios disponibles sobre corderos de razas de lana provenientes de parto sencillo (Carrasco et al. 2009) y en ovinos de pelo en finalización (Martínez et al. 1990, Cantón et al. 1992).
Los corderos de nacimiento sencillo mostraron un mayor peso del hueso en tórax y del TB en pierna con respecto a los de nacimiento gemelar. En el caso del peso del hueso en tórax, es importante considerar que en este corte primario se tuvo la menor relación TB:hueso y que el perímetro del tórax es una medida corporal que muestra una relación positiva con el peso vivo del animal, al respecto se sabe que un incremento en el perímetro del tórax implica una cavidad torácica de mayor tamaño (López-Carlos et al. 2010). En este sentido, los corderos de nacimiento presentaron una tendencia (p = 0.06) a un mayor peso al sacrificio, lo que pudiera explicar que en este tipo de corderos se haya detectado un mayor peso del hueso en tórax con relación a corderos de nacimiento gemelar. Adicionalmente, se ha indicado que el perímetro torácico de corderos de nacimiento gemelar es de menor magnitud con respecto al de corderos de nacimiento sencillo (Hurtado et al. 2003). Circunstancia que contribuye a explicar la mayor proporción de hueso en tórax en corderos de nacimiento sencillo.
Con respecto al TB en pierna, los corderos de nacimiento sencillo mostraron un mayor peso del TB con relación a los de nacimiento gemelar. Al respecto se sabe que los fetos de gestación sencilla tienen una mayor masa muscular con relación a los fetos de gestación gemelar (Sales et al. 2013). Por lo que durante el desarrollo muscular posnatal la superioridad de los corderos de nacimiento sencillo se manifieste en mayor desarrollo muscular de las piernas, debido a que estas representan el corte primario con mayor TB de la canal, así como la mayor relación TB:hueso con relación a los otros cortes primarios. Al respecto Ruíz de Huidobro y Cañeque (1994) reportan que el músculo de la pierna muestra mayor peso y contribución porcentual en la canal en corderos; mientras que el músculo en el cuello tuvo el menor peso y participación porcentual en la canal, estos resultados concuerdan con lo obtenido en la presente investigación.
La magnitud de la relación TB:hueso en cada uno de los cortes primarios, está determinada en parte por el número, tamaño y peso de los huesos, así como por la función y peso del TB. El tórax presentó la menor relación TB:hueso de los cinco cortes estudiados, lo que puede atribuirse, a que tiene el mayor número y peso de huesos (Akers y Denbow 2013). El cuello, es el corte más liviano y con el menor peso de TB, situación que explica su baja relación TB:hueso. El abdomen, tuvo una magnitud intermedia en su relación TB:hueso, debido a que en esta parte se encuentran los huesos que conforman la región dorsal, sacra y coccígea de la columna vertebral, así como los huesos de la pelvis. La proporción de TB está dada, por los músculos del lomo y abdomen (Akers y Denbow 2013). Los cortes primarios brazo y pierna fueron los que presentaron la mayor relación TB:hueso, debido a que en estos miembros se encuentran los músculos con mayor peso y volumen (Akers y Denbow 2013).
El crecimiento de las regiones corporales y la composición tisular en la canal tiene diferencias en su ritmo de crecimiento, el cual depende de la dieta. En este sentido, en ovinos Pelibuey en finalización, la pierna y espaldilla presentan un crecimiento precoz con respecto al abdomen, tórax y cuello, siendo el tórax la región que crece más tarde. En lo referente a los componentes tisulares de la canal, el hueso es el que crece más rápido, el músculo crece de forma intermedia y la grasa de forma tardía (Partida y Martínez 2010). Las diferencias en la relación TB:hueso en los cortes de corderos de nacimiento sencillo y gemelar pueden deberse a una superioridad que muestran los corderos de nacimiento sencillo en ganancia de peso, crecimiento de regiones corporales y velocidad de crecimiento tisular.
Predicción de la composición tisular de la canal
A diferencia de lo detectado en corderos de nacimiento gemelar, en los de nacimiento sencillo un menor número de variables de la composición tisular de los cortes primarios tuvo relación con el peso del TB y hueso de la canal. Lo que puede deberse a diferencias en la velocidad de crecimiento de los tejidos y de las regiones anatómicas en la etapa posnatal (Lawrence y Fowler 2002, Partida y Martínez 2010). Adicionalmente, estas diferencias pueden ser mayores entre corderos de nacimiento sencillo y gemelar. Es importante considerar que la comparación de resultados entre estudios sobre composición tisular de los cortes primarios de la canal se dificulta, debido a la gran divergencia en la manera de despiezar las canales ovinas. Además de los diferentes nombres que reciben cada una de las partes, ya que se destinan a usos múltiples (Partida y Braña 2011). Por lo que la asociación entre composición tisular de los cortes primarios de la canal con la composición tisular de la canal puede diferir entre estudios, debido a que esta puede ser afectada por la metodología para determinar los componentes tisulares en un corte primario, el número y tipo de cortes (Miguélez et al. 2007) y la raza (Luaces et al. 2008). Aun con estas diferencias los resultados permiten identificar factores que muestran estrecha relación con la composición tisular de la canal (Miguélez et al. 2007, Partida y Braña 2011).
Al respecto, Luaces et al. (2008) en un estudio con corderos lechales de la raza Gallega de nacimiento sencillo, encontraron relación entre el músculo de seis cortes primarios con el músculo de la canal. Estos resultados son similares a los del presente estudio, en el sentido de que el TB en cuatro de los cinco cortes primarios tuvo relación con el TB en canal.
En estudios de calidad de carne y composición tisular de la canal de corderos de nacimiento sencillo, con frecuencia se utilizan los cortes de brazo y pierna (Luaces et al. 2008, Pérez et al. 2012). Mientras que en ovinos adultos de pelo se utiliza la región del brazo o espaldilla para estudiar la calidad de la carne (Partida et al. 2009, Vázquez et al. 2011). En el presente estudio se encontró que la inclusión del peso del TB en el brazo en la ecuación de predicción permitió explicar el 96 % de la variación en TB en la canal. Sin embargo, en los corderos de nacimiento gemelar el peso del TB en la canal se estimó con precisión con una ecuación que consideró el peso del TB en pierna. Al parecer, las diferencias que muestran corderos de nacimiento sencillo y gemelar en la composición tisular de sus cortes primarios permiten que algunas regiones corporales tengan mayor magnitud de predicción del TB en canal. Los resultados obtenidos en el presente estudio, no concuerdan con lo indicado por Cantón et al. (1992) en ovinos Blackbelly y Blackbelly x Pelibuey en finalización, en donde el TB en tórax permitió predecir (R2= 0.86) el TB en la canal. Por otra parte, Luaces et al. (2008) establecieron que la variación en el músculo en canal en corderos Gallegos tipo lechal requiere incorporar en la ecuación (R2= 0.82) las variables: músculo (R2 parcial = 0.76), hueso (R2 parcial = 0.04) y grasa subcutánea de la espalda (R2 parcial = 0.02); en corderos ternascos la predicción del músculo en canal (R2= 0.71) se estableció con una ecuación que incluyó dos variables: músculo (R2 parcial = 0.67) y hueso de la espalda (R2 parcial = 0.04). En el presente estudio las ecuaciones generadas para predecir el peso del TB y hueso en canal solo consideraron una sola variable independiente, debido a que se detectó correlación (p < 0.05) entre las variables que describen la composición tisular de los cortes primarios incluidos en el estudio. Adicionalmente, es factible que las diferencias entre estudios en cuanto a la magnitud para predecir la variación del TB en canal por medio de la composición tisular en los cortes primarios se atribuya a edad de los ovinos, raza, TN, número de cortes primarios y metodologías para establecer la composición tisular en los cortes primarios (Miguélez et al. 2007).
En corderos de nacimiento sencillo, la variación en el peso del hueso en la canal se explicó con una ecuación que consideró el peso del hueso en brazo. Lo que concuerda con Luaces et al. (2008) quienes encontraron que las ecuaciones para determinar el hueso en canal consideraron el hueso en la espalda (brazo), con una R2 de menor magnitud a la detectada en el presente estudio. Al parecer, en corderos de nacimiento sencillo Blackbelly x Pelibuey el brazo es un corte primario con gran valor para predecir la composición tisular de la canal, ya que su composición permite explicar de forma adecuada la variación en el peso del TB y hueso en canal. En corderos de nacimiento gemelar, el peso del hueso en la canal se predijo con una ecuación que incluyó el peso del hueso en tórax. Lo que coincide con lo reportado por Cantón et al. (1992) quienes indican que la ecuación para determinar el hueso de la canal consideró el hueso en tórax.
Las ecuaciones para predecir el rendimiento de TB y hueso resultaron de baja magnitud (R2 < 0.48). Resultados similares fueron obtenidos por Carrasco et al. (2009) en corderos de nacimiento sencillo con R2 de 0.87 para predecir el peso y de 0.58 % para rendimiento de músculo. Cuando se obtiene una ecuación con una R2 de bajo valor, se sugiere considerar otro tipo de variables con mayor valor predictor (Herrera y García 2011). Lo cual difiere con Carrasco et al. (2009) quienes encontraron R2 de 0.87 para predecir el peso y de 0.58 % para rendimiento de músculo en corderos de nacimiento sencillo. Por lo que Luaces et al. (2008) proponen incorporar variables no depreciadoras de la canal al modelo, como peso y anchura de la cabeza, además del peso de las extremidades para predecir hueso en canal, en tanto que Carrasco et al. (2009) sugieren incluir en el modelo el ancho de la canal y el peso de la grasa que cubre a los riñones.
Conclusiones
El número de parto de las ovejas no afectó la composición tisular de los cortes primarios y de la canal en corderos lactantes Blackbelly x Pelibuey. Sin embargo, el peso de la canal caliente de corderos de nacimiento sencillo fue mayor que el de nacimiento gemelar, lo cual se atribuyó al mayor peso del tejido blando y del hueso en los cortes primarios del tórax y pierna. En nacimiento sencillo y gemelar, el peso del tejido blando y hueso en los cortes primarios presentaron asociación con el peso del tejido blando y hueso de la canal. En corderos de nacimiento sencillo, el peso del tejido blando y hueso en brazo tuvieron un alto valor para predecir la composición tisular de la canal. Mientras que en corderos de nacimiento gemelar, el peso del tejido blando en pierna y hueso en tórax mostraron alto valor para predecir el peso del tejido blando en canal y hueso en canal, respectivamente.

