










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkEcosistemas y recursos agropecuarios
versão On-line ISSN 2007-901Xversão impressa ISSN 2007-9028
Ecosistemas y recur. agropecuarios vol.3 no.7 Villahermosa Jan./Abr. 2016
Artículo científico
Caracterización isoenzimática de cultivares de nopal (Opuntia spp.)
Isoenzyme characterization of nopal cultivars (Opuntia spp.)
Arnoldo Flores-Hernandez1, Cecilia B. Peña-Valdivia2, Luis Hernández-Montiel3, Rogelio Ramírez-Serrano3, Ricardo Trejo-Calzada1, Cesar Alberto Meza-Herrera1, Pablo Preciado-Rangel4, Bernardo Murillo-Amador3*
1 Unidad Regional Universitaria de Zonas Áridas-Universidad Autónoma Chapingo. Bermejillo, Durango, México.
2 Botánica, Colegio de Postgraduados, Campus Montecillo, México.
3 Centro de Investigaciones Biológicas del Noroeste, S.C., La Paz, Baja California Sur, México. *Autor de correspondencia: bmurillo04@cibnor.mx
4 Instituto Tecnológico de Torreón, Torreón, Coahuila, México.
Artículo recibido el 27 de enero de 2015
Aceptado el 4 de agosto de 2015
RESUMEN
En nopal (Opuntia spp.) como en muchos géneros, los problemas de clasificación taxonómica son frecuentes, esto complica la identificación correcta de cultivares. Este estudio se realizó con la finalidad de detectar polimorfismo en nueve cultivares de nopal de las especies O. amyclaea Ten. (O. albicarpa), O. megacantha, O. crassa y O. ficus-indica (L.) Mili., ubicados en la reserva de germoplasma de la Unidad Regional Universitaria de Zonas Áridas de la Universidad Autónoma Chapingo. La proteína soluble de las raíces se separó por electroforesis para evaluar la presencia de nueve sistemas enzimáticos. De estos, fosfoglucomutasa (PGM), 6-fosfogluconato deshidrogenasa (6-PGD), glutamato oxaloacetato transaminasa (GOT) y malato deshidrogenasa (MDH) expresaron reacción suficiente para una identificación de bandas de actividad enzimática. Las enzimas málicas (ME) y aconitasa (ACO) no presentaron polimorfismo, por tanto no se recomiendan para la clasificación de cultivares de nopal. En contraste, PGM, 6-PGD, GOT y MDH presentaron diferencias o polimorfismo indeterminado en su patrón de bandeo, por lo que se consideran relevantes para la identificación taxonómica y evaluación de la variabilidad genética de nopal.
Palabras clave: Enzimas, marcadores moleculares, taxonomía, variabilidad.
ABSTRACT
Taxonomic classification problems are frequent in the case of nopal (Opuntia spp.), as well as for many other genera; this complicates the accurate identification of cultivars. This study was done with the objective of detecting polymorphism in nine nopal cultivars of the 0. amyclaea Ten. (0. albicarpa), 0. megacantha, 0. crassa and 0. ficus-indica (L.) Mill, species located in the germplasm reserve of the Unidad Regional Universitaria de Zonas Aridas of the Universidad Autonoma Chapingo. The soluble protein of the roots was separated through electrophoresis to evaluate the presence of nine enzyme systems. Among these, phosphoglucomutase (PGM), 6- phosphogluconate dehydrogenase (6-PGD), glutamate oxaloacetate transaminase (GOT) and malate dehydrogenase (MDH) showed a sufficient reaction for the identification of bands of enzymatic activity. The malic enzymes (ME) and aconitase (ACO) did not present polymorphism; they are therefore not recommended for the classification of nopal cultivars. In contrast, PGM, 6-PGD, GOT and MDH presented differences or undetermined polymorphism in their band pattern; they are therefore considered relevant for the taxonomic identification and evaluation of the genetic variability of nopal.
Key words: Enzymes, molecular markers, taxonomy, variability.
INTRODUCCIÓN
Las cactáceas originarias de América constituyen una de las familias de mayor distribución y variación e incluye 98 géneros y más de 1 500 especies. El género Opuntia presenta gran distribución y representatividad, con más de 200 especies desde Canadá hasta la Patagonia (llloldi-Rangel et al. 2012). Uno de los principales centros de origen del nopal es México (Maki-Díaz et al. 2015). La clasificación del género Opuntia se complica por la diversidad de especies y ecotipos, producto de procesos de hibridación natural debido a su proceso de propagación sexual o asexual, que permite una gran cantidad de variantes (Gallegos-Vázquez y Mondragón-Jacobo 2011), así como por su plasticidad morfológica, resultado de la constitución genética e interacción con el ambiente (Mad-dams 1972). Las clasificaciones desarrolladas por Sánchez-Mejorada (1982) y Barthlott y Hunt (1993) se basan en diferencias fenotípicas que son la base para identificar diversos géneros y especies. Sin embargo, presentan debilidades para una identificación taxonómica correcta en el género Opuntia, ya que con cierta frecuencia existe coincidencia limitada entre las características de las plantas in situ y la descripción de los sistemas de clasificación. Por lo que es preferible realizar la clasificación mediante la aproximación de caracteres (Peña-Valdivia et al. 2008). El uso de los métodos de clasificación comunes generan numerosas sinonimias; además, los problemas se acentúan por las descripciones que se realizan con ejemplares únicos de jardines y herbarios, de tal forma que la variabilidad natural de las poblaciones respectivas se ha excluido, a pesar de que es fundamental para la descripción apropiada de las especies y conformación de grupos (Schneider 1999).
La preservación de Opuntia, como recurso genético, es importante. Se requiere de investigación fisiológica, morfológica, genética, bioquímica y taxonómica para expandir la información y el conocimiento acerca de la diversidad de especies, el efecto de los cambios ambientales y las modificaciones que resultan de su domesticación (Montiel et al. 2012). En este sentido, la clasificación de variedades y especies silvestres de nopal se han basado en características morfológicas de los frutos y cladodios (Valdez-Cepeda et al. 2003, Reyes-Agüero et al. 2005, Peña-Valdivia et al. 2008, Gallegos-Vázquez et al. 2011, Gallegos-Vázquez et al. 2012, Espinoza-Sánchez et al. 2014, Samah y Valadez-Moctezuma 2014), tolerancia a las heladas (Parish y Felker 1998), nomenclatura y clasificación vernácula, la ubicación de nopaleras silvestres, el uso y manejo de Opuntia spp. (Ávalos-Huerta et al. 2013). Otras investigaciones han definido e identificado variedades de Opuntia spp. y otras cactáceas al usar marcadores moleculares como isoenzimas (Chessa et al. 1997, Uzun 1997), RAPDs (Wang et al. 1998, Mondragón 2003, Luna-Paez et al. 2007, Zoghlami et al. 2007, Bendhifi et al. 2013, Valadez-Moctezuma et al. 2014b), cpSSR (Labra et al. 2003, Caruso et al. 2010), ISSR (Luna-Paez et al. 2007, Valadez-Moctezuma et al. 2014b, Ganopoulos et al. 2015, Contreras-Negrete et al. 2015), AFLP (Labra et al. 2003, Espinoza-Sánchez et al. 2014), análisis filogenético Bayesiano de nrITS de secuencias de ADN (Griffith 2004, Lyra et al. 2013). Los marcadores moleculares basados en la amplificación por PCR de genoma específico de secuencias, se han propuesto como herramientas directas y eficaces para estimar la relación intergenérica e interespecífica entre los diferentes taxones (Labra et al. 2003). En este sentido, el análisis de los caracteres para la clasificación de las plantas avanza y se profundiza paulatinamente en diferentes campos del conocimiento como el anatómico, morfológico (Scheinvar et al. 2010), genético (citogenético) y bioquímico. Para los aspectos bioquímicos se ha recurrido a la electro-foresis y a la secuencia de moléculas químicas como ADN, proteínas e isoenzimas en diferentes órganos de la planta (Uzun 1997, Henry 2012, Lyra et al. 2013).
En nopal, el análisis de isoenzimas se utiliza para su clasificación taxonómica, a la vez que puede ampliar y complementar la descripción varietal de la especie. Además, las isoenzimas son marcadores ideales debido a que son colineales con el gen, por lo común codominante en efecto y poco afectadas por el ambiente. El fortalecimiento de la caracterización del nopal con herramientas alternativas, permite conocer de forma más eficiente la diversidad genética existente, apoyar su ubicación taxonómica, contribuir en programas de mejoramiento genético para la creación de nuevas variedades, así como enriquecer los bancos de germoplasma (Chessa et al. 1997), en este caso del nopal, el cual es reconocido por su importancia como recurso natural, pues colecciones de valor comercial o agronómico que se describen desde un punto de vista morfológico y antropocéntrico se protegen en bancos de germoplasma (Bellon et al. 2009). En este sentido, el objetivo del presente trabajo fue detectar polimorfismo en nueve cultivares de nopal caracterizados por producir brotes (nopalito) y frutas (tuna) de interés comercial de las especies O. amyclaea Ten. (O. albicarpa), O. megacantha, O. crassa y O. ficusindica (L.) Mili.
MATERIALES Y MÉTODOS
Área de estudio
La presente investigación se realizó en cultivares de nopal establecidos en la zona desértica de la Reserva de la Biosfera Mapimí, Durango, en la plantación experimental de Opuntia de la Unidad Regional Universitaria de Zonas Áridas de la Universidad Autónoma Chapingo (URUZA-UACh) en Bermejillo, Durango, México. Ubicada en las coordenadas geográficas 25° 52' 28" LN y 103° 37' 07" LO, a 1 119 msnm, con una temperatura anual promedio de 21 °C y precipitación anual promedio de 200 mm, registrándose el 50 % durante la estación de verano (García 1988).
Las plantas de nopal utilizadas pertenecen al banco de germoplasma de la URUZA-UACh que incluyen 55 cultivares de Opuntia, los cuales se obtuvieron de diferentes centros de investigación, universidades públicas y plantaciones comerciales de México, establecidas a 1.20 m entre hileras y 1.50 m entre plantas, con densidad de plantación de 5 555 plantas ha-1, las cuales no se riegan, fertilizan, ni se realiza manejo fitosanitario, sólo se lleva a cabo la eliminación de malezas de forma manual, así como la poda para regular el desarrollo de las plantas.
Obtención de raíces y extracción de proteínas
Se utilizaron nueve cultivares de Opuntia (Tabla 1) que corresponden a las especies O. amyclaea Ten. (O. albicarpa) (cvs. 1, 4 y 6), O. megacantha (cvs. 2, 3 y 9), O. crassa (cv. 5) y O. ficus-indica (L.) Mili. (cvs. 7 y 8). Las raíces de tres cladodios de cada cultivar se utilizaron para determinar las enzimas. Para la obtención de las raíces, los cladodios de cada cultivar se colocaron en bandejas con sustrato (vermiculita) a capacidad de campo, mantenidas en el laboratorio a temperatura ambiente (25 a 30 °C) durante 20 d, periodo en que emitieron las raíces, manteniéndose en estas condiciones mientras se colectaron las raíces, las cuales se extrajeron de la base del cladodio mediante cortes con bisturí, se lavaron con agua potable y luego con agua destilada. La extracción de proteínas se realizó mediante la maceración del tejido de dos segmentos de raíz con longitudes entre 2 y 3 cm, para luego poner por tres segundos en homogenizador con 1.5 mL de solución extractora compuesta por 20 mL de Tris-HCI pH 0.5 M y 7.5, 3.0 M de sacarosa, 1.6 M de ácido ascorbico, 1.6 M de polivinil pirrolidona, 0.1 M de metabisulfito de sodio y 22 µL de 2-mercaptoetanol. Del sobrenadante que se obtuvo por centrifugación a 14 000 rpm durante 10 min a -10 °C, se tomaron alícuotas para la separación de las proteínas (Bradford, 1976).
Electroforesis en gel de almidón
La separación de las proteínas por electroforesis en gel de almidón y la detección de las isoenzimas se realizó con la técnica descrita por Stuber et al. (1982). La solución de bromofenol se usó como indicador de la migración de las proteínas y la mezcla de ácido bórico: hidróxido de litio y L-histidina: ácido cítrico para la electroforesis. La separación se realizó en cámara horizontal con 140 V durante 5 h y 200 V, durante 6 h. En el primero y último carril del gel se aplicó un extracto de raíz de frijol (Phaseolus vulgaris L.) como muestra de referencia. Los sistemas enzimáticos evaluados fueron fosfoglu-comutasa (PGM, E.C. 2.7.5.1), 6-fosfogluconato deshidrogenasa (6-PGD, E.C. 1.1.1.44), glutamato oxaloacetato transaminasa (GOT, E.C. 2.6.1.1), aconitasa (ACO, E.C. 4.2.1.3), enzima málica (ME, E.C. 1.1.1.40), malato deshidrogenasa (MDH, E.C. 1.1.1.37), ácido shikímico deshidrogenasa (SAD, E.C. 1.1.1.25), fosfatasa ácida (ACP, E.C. 3.1.3.2) y fosfoglucosa isomerasa (PHI, E.C. 5.3.1.9). La localización e identificación de bandas de los sistemas enzimáticos y sus variantes isoenzimáticas se realizó a partir de su Rf (movilidad relativa de la proteína) en el gel, utilizando los dibujos de los zimogramas previamente fotografiados.
Análisis estadístico
Cada banda de cada gel se consideró como un carácter o locus independiente, que se codificó visualmente como polimorfica ausente (0) o presente (1) para cada uno de los cultivares y anotadas como variables discretas, las diferencias cualitativas entre bandas no se consideraron. Para tener una mayor confiabilidad y precisión de los datos, se utilizaron tres repeticiones independientes de cada cultivar, además del testigo (Phaseolus vulgaris L.) que se colocó en la primera y última banda. Sólo las bandas con actividad satisfactoria, buena resolución y resultados consistentes se seleccionaron y se utilizaron para los análisis respectivos. La matriz básica de datos se generó con la información de cada enzima. La similitud genética se estimó mediante el coeficiente de Jaccard y para el agrupamiento se utilizó el método UPGMA. Los dendogramas se realizaron con el programa MVSP v. 3.1 (Kovach 1999).
RESULTADOS
Identificación de zonas de actividad enzimática
De los nueve sistemas enzimáticos solo seis presentaron definición aceptable para la identificación de zonas de actividad enzimática, los cuales fueron fosfoglucomutasa (PGM), 6-fosfogluconato deshidrogenasa (6-PGD), glutamato oxaloacetato transaminasa (GOT), aconitasa (ACO), enzima málica (ME) y malato deshidrogenasa (MDH). Los sistemas ácido shikímico deshidrogenasa (SAD), fosfatasa ácida (ACP) y 6-fosfoglucosa isomerasa (PHI) mostraron poca definición y fue imposible la identificación correcta de las bandas de actividad enzimática atribuibles a los diferentes cultivares. En la Figura 1 se muestra el bandeo obtenido de las seis enzimas de mayor definición. Los sistemas enzimáticos ACO y ME se distinguieron porque muestran resultados similares en la expresión enzimática (monomórficos), mientras que los demás exhibieron polimorfismo en mayor o menor grado en todos los cultivares. Las enzimas ACO y ME del primer grupo no presentaron diferencias en su expresión, es decir, no aportaron información importante para la identificación de los cultivares. En el segundo grupo, PGM, 6-PGD, GOT y MDH mostraron diferencias en el patrón de bandeo atribuibles a los cultivares, por lo que es conveniente utilizarlas en estudios futuros de identidad y evaluación de la variabilidad genética.

Enzimas de nopal
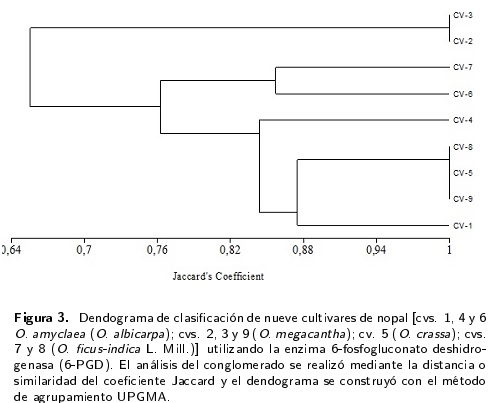
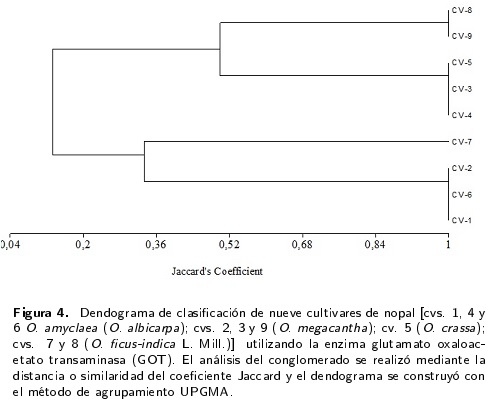
El análisis particular de cada enzima muestra que entre las enzimas monomórficas, la aconitasa (ACO) se encontró sólo en una zona de actividad enzimática en todos los cultivares y está codificada por un locus (ACO1) con alelos en condición homocigota (Figura 1). La enzima mélica (ME), básica en el ciclo de carboxilación-descarboxilación de las plantas con metabolismo ácido de las crasuláceas (CAM) como el nopal, se caracterizó por la presencia de un locus con tres bandas en todos los cultivares, a una distancia considerable (Figura 1); ello dificulta la circunspección pues se trata de un locus con tres alelos. De los sistemas isoenzimáticos que exhibieron polimorfismo, el sistema GOT presentó zonas de actividad definidas con claridad (Figura 1) e indica dos locus, GOT1 y GOT2 con dos alelos en condición homocigota o heterocigota de acuerdo con el cultivar de nopal. Por la distribución de las bandas, se estableció la presencia de alelos nulos. Los sistemas isoenzimáticos PGM, 6-PGD y MDH presentaron el mayor número de zonas de actividad enzimática (Figura 1), lo que no facilita su interpretación adecuada. Para PGM (Figuras 1 y 2) se detectaron de tres a cinco bandas por cultivar, las cuales de acuerdo a sus distancias se agrupan en cinco loci con dos alelos por locus, todos en condición homocigota dominante o recesiva según el cultivar; los cultivares 1 y 9 presentaron alelos nulos. En la enzima 6-PGD (Figura 1) se mostraron de cinco a ocho bandas por cultivar (siete bandas fue lo más frecuente). De acuerdo a las distancias del patrón de bandeo, se manifiesta la posibilidad de cuatro loci, con dos y tres alelos según el cultivar. En el sistema enzimático MDH (Figuras 1 y 5) se detectaron entre cinco y ocho bandas por cultivar (siete bandas fue lo más frecuente). De acuerdo a las distancias del patrón de bandeo, puede presentarse cinco loci con dos alelos por locus por loci.


Especies de nopal
El análisis de las especies en estudio mostró que la similitud en el patrón isoenzimático de los cultivares ubicados en una sola especie fue inexistente en los sistemas PGM, 6-PGD y MDH (Figura 1), es decir, en la misma especie se presentan diferencias en su patrón de bandeo (polimorfismo). El dendograma de la isoenzima PGM (Figura 2) muestra dos ramificaciones, la primera, representada por los cultivares 1 y 2, clasificados como O. amyclaea y O. megacantha, respectivamente. La segunda ramificación se subdivide en cinco grupos; el primero agrupa a los cultivares 7 y 8, los cuales corresponden a la misma especie (O. ficus-indica); en el segundo se mantiene independiente el cultivar 5 (O. crassa); en el tercero y cuarto grupo se encuentran los cultivares 6 (O. amyclaea) y 9 (O. megacantha) como independientes; mientras que en el grupo cinco se ubican los cultivares 3 y 4 que corresponden a O. megacantha y O. amyclaea, respectivamente.
El dendograma formado por la enzima 6-PGD (Figura 3) muestra dos ramificaciones; la primera por los cultivares 2 y 3, ambos clasificados como O. megacantha. En la segunda ramificación aparecen cuatro grupos; el primero por los cultivares 6 (O. amyclaea) y 7 (O. ficus-indica)] el segundo por el cultivar 4(0. amyclaea) como independiente; el tercero formado por los cultivares 5 (O. crassa), 8 (O. ficus-indica) y 9 (O. megacantha). El cultivar 1 (O. amyclaea) aparece en el cuarto grupo de forma independiente. El dendograma de la enzima GOT (Figura 4) presentó dos ramificaciones, ambas con dos grupos. En el primer grupo con los cultivares 1 y 6 (O. amyclaea) y 2 (O. megacantha) y en un segundo con el cultivar 7 (O. ficus-indica) como independiente. La segunda ramificación mostró un primer grupo con los cultivares 3 (O. megacantha), 4 (O. amyclaea) y 5 (O. crassa) y un segundo con los cultivares 8 (O. ficus-indica) y 9 (O. megacantha).
El dendograma de la enzima MDH (Figura 5) presentó dos ramificaciones principales. La primera que se subdivide en dos grupos, en el primero se agruparon los cultivares 1 y 6 (O. amyclaea) y en el segundo el cultivar 5 (O. crassa) como independiente. La segunda ramificación se subdivide en cuatro grupos, el primero formado por los cultivares 3 (O. megacantha) y 4 (O. amyclaea), el segundo por los cultivares 7 y 8, ambos de O. ficus-indica, el tercero formado por el cultivar 9 (O. megacantha) y el cuarto por el cultivar 2 (O. megacantha).
DISCUSIÓN
Enzimas de nopal
Un estudio con isoenzimas en polen de nopal (Chessa et al. 1997) demostró que la enzima mélica presentó un patrón único de bandeo para todos los genotipos estudiados, por lo que se descartó como útil para la identificación de genotipos; lo cual coinciden con los resultados encontrados en el presente estudio. El sistema isoenzimático GOT presentó zonas de actividad definidas con claridad, indicando dos locus, GOT1 y GOT2 con dos alelos en condición homocigota o heterocigota, lo cual difiere de lo reportado por Uzun (1997), que encontró solo una banda isoenzimática con actividad definida para frutos y cladodios de nopal. También difieren del resultado obtenido por Chessa et al. (1997), quienes encontraron que sólo un patrón de banda para GOT se presentó en todos los genotipos de nopal cuando se utilizó tejido de cladodios, mientras que cuando se utilizó tejido de polen, se detectaron tres bandas isoenzimáticas; sin embargo, la actividad de GOT no fue diferenciada de forma clara para algunos genotipos, por lo que no se utilizó para la identificación de genotipos en esa etapa de investigación.
En cuanto a la presencia de alelos nulos en algunos cultivares, cabe señalar que el alelo nulo está ausente y se considera que su expresión en la síntesis del producto proteico es reducida, lo que puede indicar un homocigoto nulo. Al respecto, la interpretación de los resultados de los ensayos en los que se involucran alelos nulos debe confirmarse con estudios de mejoramiento genético (Murphy et al. 1990). Mientras que Chessa et al. (1997) reportaron sólo un patrón de bandeo del sistema enzimático PGM cuando se utilizó tejido de cladodios para todos los genotipos de nopal estudiados, en tanto que al utilizar tejido de polen, el sistema enzimático PGM mostró una variabilidad considerable con tres diferentes patrones de bandeo.
Los resultados mostrados en la enzima 6-PGD difieren de los reportados por Chessa et al. (1997) quienes encontraron sólo un patrón de bandeo, cuando se utilizó tejido de cladodios para todos los genotipos de nopal estudiados, mientras que al utilizar tejido de polen, este sistema enzimático mostró dos diferentes patrones de bandeo. Los resultados del presente estudio con la enzima MDH difieren de aquellos reportados en tejidos de cladodios en los que se presentaron cuatro bandas bien definidas (Uzun 1997), tres bandas en tejido verde (Mukerji y Ting 1969) y tres bandas en tejido de frutos (Uzun 1997). Sin embargo, los resultados reportados por Chessa et al. (1997) para el caso de polen de nopal, coinciden en parte con los del presente estudio, pues ellos reportaron que el sistema enzimático MDH mostró alta variabilidad, con nueve diferentes patrones de bandeo. La cantidad de loci detectada en MDH apoya la sugerencia de una estructura compleja. Al respecto Hardisty y Claire (1989) encontraron en O. cymochila y O. phaeacantha dos o más loci en el control de la expresión de la MDH, por lo que las diferencias en el patrón de bandas de esta enzima, entre estas dos especies de Opuntia, permiten sugerir su uso como marcador taxonómico. Al respecto se sabe que puede haber la posibilidad de que la enzima 6-PGD de nopal, como en el caso de MDH, produzca heterodímeros (Stuber et al. 1982). Los sistemas enzimáticos MDH, PGI y PGM han mostrado una variabilidad considerable en tejido de polen de nopal y son especialmente útiles para la identificación de genotipos de Opuntia, debido a sus distintas bandas y el número de patrones proporcionados; sin embargo, la identificación de un cultivar no es posible usando sólo sistemas isoenzimáticos (Chessa et al. 1997). La enzima malato deshidrogenasa (MDH) ha mostrado que es altamente polimorfica, lo que significa que tiene múltiples formas moleculares; encontrándose que el polimorfismo de MDH en cada especie está determinado genéticamente por varios loci con alelos múltiples, por lo que una alta actividad de MDH en diversos órganos y tejidos, hace de esta enzima un marcador genético confiable, conveniente y se puede utilizar en estudios de ecología y genética de poblaciones (Yudina 2012).
Las isoenzimas fueron los primeros marcadores moleculares desarrollados, resultan de la variación en la secuencia de nucleótidos que repercute en la substitución de un aminoácido por otro; este reemplazo puede implicar una alteración de la carga eléctrica neta de una proteína. La diferencia de cargas se detecta como una alteración de la tasa de migración de una proteína a través de un campo eléctrico. La separación electroforética se utiliza para medir la variación de la movilidad de la proteína dentro de una población (Klug y Cummings 2000). Así, las distintas formas electroforéticas de una proteína pueden implicar una codificación de diferentes alelos, es decir, variación genética. La electroforesis y el análisis de isoenzimas se han aplicado exitosamente en una amplia variedad de especies de plantas y se utilizan para varios propósitos, entre los que destacan la identificación de cultivares de manzana (Weeden y Lamb 1985), verificar el parentesco de los híbridos de melocotón y almendro (Carter y Brock 1980), como marcador genético en melocotón (Durham et al. 1987), evaluar la relación filo-genética entre Agave fourcroydes y A. angustifolia (Colunga-GarcíaMarín et al. 1999), valorar la diversidad genética de Pilosocereus tillianus (Figueredo et al. 2010) y Stenocereus pruinosus (Parra et al. 2008), identificar la variabilidad genética de maíz (Sánchez et al. 2000) y aguacate (Torres y Berg 1980). Sin embargo, en la actualidad, las isoenzimas han sido superadas por los marcadores de ADN, debido al número reducido de marcadores que generan, además, pueden ser afectadas por el tipo de tejido y etapa de desarrollo de una planta, ya que son producto de la expresión de genes. Las proteínas, también, pueden modificarse después de la traducción y alterar su movilidad electroforética (Kumar 1999); sin embargo, no todas las sustituciones cambian la carga eléctrica neta en la molécula, ya que aproximadamente 30 % de la variación real asociada a sustituciones de aminoácidos se detecta por electroforesis (Klug y Cummings 2000). Los patrones electroforéticos de proteínas de semillas y tallos de dos especies de Opuntia (O. rzedowskii Scheinvar y O. robusta Wenland ssp. larreyi Bravo) se estudiaron, se distinguieron bandas comunes a todos los taxones examinados, por lo que podrían ser características del género Opuntia, ya que los taxones con bandas similares son interpretados como filogenéticamente cercanos, por lo que se concluye que la electroforesis es una herramienta auxiliar en la identificación y clasificación de Opuntia (Estrada-Galván et al. 2000). Recientes estudios utilizando patrones electroforéticos de proteínas de semillas de Opuntia demostraron que no existe una separación entre las especies de nopales dulces y nopales ácidos o xoconostles. En base a los marcadores bioquímicos utilizados, se sugiere una revisión de la asignación taxonómica de los genotipos que pertenecen al género Opuntia (Samah et al. 2015), además, estos resultados soportan la hipótesis de la existencia de un reducido número de especies de Opuntia en México (Valadez-Moctezuma et al. 2014a).
Especies de nopal
Los resultados de los dendogramas son una clara evidencia de las diferentes especies que pueden presentar el mismo patrón isoenzimático, lo que refleja polimorfismo y probable relación genómica menor entre las especies utilizadas, esto sugiere que pueden compartir ciertos rasgos fenotípicos comunes a pesar de ser especies diferentes (Guerrero-Muñoz et al. 2006). El polimorfismo se ha observado en especies como O. robusta, que es endémica de México, sexualmente dimórfica, con una ecología e historia de vida desconocida (Janczur et al. 2014). También se ha observado polimorfismo en O. ficusindica (Ganopoulus et al. 2015, Bendhifi et al. 2015). Lo que corrobora la discrepancia en los diferentes métodos de clasificación de nopal, por lo que se fortalece la hipótesis de que al efectuar la clasificación del nopal por aproximación de caracteres se han generado numerosas sinonimias. Estos problemas se han acentuado por las descripciones realizadas con genotipos únicos de jardines y herbarios, pues no se consideró la variabilidad natural de las poblaciones, la cual es fundamental para la descripción apropiada de las especies y conformación de grupos (Schneider 1999, Reyes-Agüero et al. 2005). La interpretación de la presencia de isoenzimas en plantas poliploides, como el nopal, puede conllevar cierto sesgo debido a que existen copias múltiples de cromosomas y su combinación depende del nivel de ploidía. El nivel octaploide (2n=8x= 88) de los cultivares de nopal estudiados, propicia la duda de que exista duplicidad de bandas en la separación electroforética y error en su interpretación. Otras posibles explicaciones es la posibilidad de recombinación genética, o que los individuos utilizados para el estudio son atípicos y no representan a las variedades referidas y, en última instancia, puede existir un error de clasificación en el banco de germoplasma de donde se tomaron los individuos de las especies. Similares resultados se han obtenido en conglomerados obtenidos a partir de RAPD, ISSR y una combinación de ambos, los cuales no coinciden con la clasificación taxonómica real, ya que las variedades clasificadas en la misma especie no se encuentran en el mismo grupo, donde O. ficus-indica, O. albicarpa y O. megacantha mostraron variación amplia sin una definición adecuada, por lo que es probable que tengan un ancestro común, lo que apoya la hipótesis sobre la existencia de un menor número de especies de Opuntia, con alta variación genética intraespecí-fica (Valadez-Moctezuma et al. 2014a). Lo anterior, es una evidencia del uso de las herramientas moleculares para la clasificación de cultivares y para la estimación de la diversidad o la variabilidad genética compleja de especies (Taylor y Barker 2012, Bendhifi et al. 2013, Shilpha et al. 2013) no siempre ha sido útil ni garantiza la caracterización de especies. La variación morfológica continua en el género Opuntia, los sinónimos, los diferentes niveles de ploidía y los descriptores morfológicos inadecuados, han llevado a errores de clasificación (Majure et al. 2012). Al respecto Espinoza-Sánchez et al (2014) encontraron que el dendrograma no mostró una clara relación con la taxonomía conocida de Opuntia. Mientras que Ganopoulos et al (2015) utilizando marcadores ISSR, encontraron alta diversidad genética entre el germoplasma de O. ficusindica, sugiriendo que este puede ser de beneficio en programas de mejoramiento genético y actividades de manejo de germoplasma de la especie. Lo anterior, es evidencia para señalar la necesidad de otros estudios que permitan detectar con precisión y confiabilidad las semejanzas y diferencias para reclasificación del género Opuntia (Wang et al. 1998), ya que estudios en O. ficus-indica mostraron una divergencia entre los cultivares estudiados y exhibieron variación continua independiente del origen geográfico, no agrupándose por separado de las otras especies de nopal, lo que indica que su clasificación taxonómica actual no está alineada con su variabilidad genética (Bendhifi et al. 2015), resultados que coinciden con los de Lyra et al. (2013) quienes argumentan la necesidad de realizar estudios moleculares y morfológicos en Opuntia, ya que la filogenia que encontraron sugiere una nueva revisión en la taxonomía, encontrando híbridos que causaron dificultad para caracterizar de una manera segura la familia Opuntiodeae. Al respecto MihaIter et al. (2011) en estudios del DNA de Opuntia, observaron discrepancias entre la caracterización molecular y la clasificación taxonómica.
CONCLUSIONES
Las enzimas fosfoglucomutasa (PGM), 6-fosfogluconato deshidrogenasa (6-PGD), glutamato oxaloacetato transaminasa (GOT) y malato deshidrogenasa (MDH) expresaron reacción para identificar bandas de actividad enzimática al mostrar diferencias o polimorfismo en su patrón de bandeo de manera indeterminada, aunque sí se consideran relevantes para la identificación taxonómica y evaluación de la variabilidad genética de nopal. Las enzimas málica (ME) y aconitasa (ACO), no se recomiendan para la clasificación de cultivares de nopal debido a su falta de polimorfismo. Los sistemas enzimáticos PGM, 6-PGD, GOT y MDH, al mostrar diferencias en el patrón de bandeo atribuibles a los cultivares estudiados, pueden ser utilizadas en estudios de identidad y evaluación de la variabilidad genética de nopal.
AGRADECIMIENTOS
Se agradece al Dr. Porfirio Ramírez Vallejo por las facilidades brindadas para la evaluación de las isoenzimas en el laboratorio de marcadores moleculares del Programa de Genética del Colegio de Postgraduados, Montecillos, Estado de México. Al Centro de Investigaciones Biológicas del Noroeste (proyecto AGROT1) por la publicación y difusión de la presente investigación.
LITERATURA CITADA
Ávalos-Huerta I, Sánchez-López DL, López-González C (2013) Nomenclatura vernácula, uso y manejo de Opuntia spp. en Santiago Bayacora, Durango, México. Revista Chapingo Serie Horticultura 19: 367-380. [ Links ]
Barthlott W, Hunt DR (1993) Cactaceae. In: Kubitzki K (Ed.). The families and genera of vascular plants. Vol. II: Flowering Plants. Springer Verlag, Berlin/Heidelberg, Germany, pp: 161-197. [ Links ]
Bellon MR, Barrientos-Priego AF, Colunga-García Marín P, Perales H, Reyes Agüero JA, Rosales-Serna R, et al. (2009) Diversidad y conservación de recursos genéticos en plantas cultivadas, en capital natural de México, vol. II: Estado de conservación y tendencias de cambio. CONABIO, México, pp: 355-382. [ Links ]
Bendhifi M, Baraket G, Zourgui L, Souid S, Salhi-Hannachi A (2013) Assessment of genetic diversity of Tunisian Barbary fig (Opuntia ficus-indica) cultivars by RAPD markers and morphological traits. Scientia Horticulture 158: 1-7. [ Links ]
Bendhifi M, Baraket G, Zourgui L, Souid S, Salhi-Hannachi A (2015) Genetic diversity and phylogenetic relationship among Tunisian cactus species (Opuntia) as revealed by random amplified microsatellite polymorphism markers. Genetic Molecular Research 14: 1423-1433. [ Links ]
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-Dye binding. Analytical Biochemistry 72: 248-254. [ Links ]
Carter GE, Brock MM (1980) Identification of peach cultivars through protein analysis. HortScience 15: 292-293. [ Links ]
Caruso MS, Curro S, Las Casas G, La Malfa S, Gentile A (2010) Microsatellite markers help assess genetic diversity among Opuntia ficus-indica cultivated genotypes and their relation with related species. Plant Systematic Evolution 90: 85-97. [ Links ]
Chessa I, Nieddu G, Serra P, Inglese P, La Mantia T (1997) Isozyme characterization of Opuntia species and varieties from Italian germplasm. Acta Horticulturae 438: 45-48. [ Links ]
Colunga-García Marín P, Coello-Coello J, Eguiarte LE, Piñero D (1999) Isozymatic variation and phylogenetic relationships between henequen (Agave fourcroydes) and its wild ancestor A. angustifolia (Agavaceae). American Journal of Botany 86: 115-123. [ Links ]
Contreras-Negrete G, Ruíz-Durán ME, Cabrera-Toledo D, Casas A, Ofelia Vargas O, Parra F (2015) Genetic diversity and structure of wild and managed populations of Polaskia chende (Cactaceae) in the Tehuacan-Cuicatlan Valley, Central Mexico: insights from SSRand allozyme markers. Genetic Resources Crop Evolution 62: 85-101. [ Links ]
Durham HE, Moore GA, Sherman WB (1987) Isozyme banding pattern and their usefulness as genetic markers in peach. Journal of the American Society for Horticultural Science 112: 1013-1018. [ Links ]
Espinoza-Sánchez EA, Silos Espino H, Flores-Benítez S, Valera-Montero LL, Rodríguez-Salazar E, Gallegos-Vázquez C, et al. (2014) Agrupamiento de genotipos de nopal (Opuntia spp.) de México por medio de la técnica de AFLPs y características del fruto. Phyton. Revista Internacional de Botánica Experimental 83: 299-306. [ Links ]
Estrada-Galván EB, Scheinvar L, Alagón A, Olalde-Parra G (2000) Resultados preliminares de electroforesis en proteínas como método auxiliar en la taxonomía de Opuntia spp. Acta Científica Potosina 15: 7-24. [ Links ]
Figueredo CJ, Nassar JM, García-Rivas AE, González-Carcacía JA (2010) Population genetic diversity and structure of Pilosocereus tillianus (Cactaceae, Cereeae), a columnar cactus endemic to the Venezuelan Andes. Journal of Arid Environments 74: 1392-1398. [ Links ]
Gallegos-Vázquez C, Barrientos-Priego AF, Reyes-Agüero JA, Núñez-Colín CA, Mondragón-Jacobo C (2011) Clusters of commercial varieties of cactus pear and xoconostle using UPOV morphological traits. Journal of the Professional Association for Cactus Development 13: 10-22. [ Links ]
Gallegos-Vázquez C, Mondragón-Jacobo C (2011) Cultivares selectos de tuna de México al mundo. Servicio Nacional de Inspección y Certificación de Semillas, Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SNICS-SAGARPA) y Universidad Autónoma Chapingo. Chapingo, México. 159p. [ Links ]
Gallegos-Vázquez C, Scheinvar L, Núñez-Colín CA, Mondragón-Jacobo C (2012) Morphological diversity of xoconostles (Opuntia spp.) or acidic cactus pears: a Mexican contribution to functional foods. Fruits 67: 109-120. [ Links ]
Ganopoulos I, Kalivas A, Kavroulakis N, Xanthopoulou A, Mastrogianni A, Koubouris G, et al. (2015) Genetic diversity of Barbary fig (Opuntia ficus-indica) collection in Greece with ISSR molecular markers. Plant Gene 2: 29-33. [ Links ]
García E (1988) Sistema de clasificación de Kóppen modificado. UNAM, México. 217p. [ Links ]
Griffith MP (2004) The origins of an important cactus crop, Opuntia ficus-indica (cactaceae): new molecular evidence. American Journal of Botany 91: 1915-1921. [ Links ]
Guerrero-Muñoz P, Zavaleta-Mancera HA, Barrientos-Priego AF, Gallegos-Vázquez C, Núñez-Colín CA, Valadez-Moctezuma E, et al. (2006) Técnica para el estudio de la micromorfología interna de semillas duras en Opuntia. Revista Fitotecnia Mexicana 29: 37-43. [ Links ]
Hardisty RM, Claire W (1989) Electrophoretic band patterns of phosphoglucoisomerase and malate dehydrogenase in the prickly pear cactus Opuntia cymochila E. and Opuntia phaeacantha (cactaceae) from southwestern Oklahoma. Proceedings of the Oklahoma Academy of Science 69: 5-9. [ Links ]
Henry RJ (2012) Molecular Markers in Plants. John Wiley & Sons. 216p. [ Links ]
Illoldi-Rangel P, Ciarleglio M, Sheinvar L, Linaje M, Sánchez-Cordero V (2012) Opuntia in Mexico: Identifying priority areas for conserving biodiversity in a multi-use landscape. PLoS ONE 7: e36650. [ Links ]
Janczur MK, León-Solano HJ, Solache-Rámos LT, Mendoza-Reyes CH, Oro-Cerro MC (2014) Chemical and physical defense traits in two sexual forms of Opuntia robusta in central eastern Mexico. PLoS ONE 9: e89535. [ Links ]
Klug WS, Cummings MR (2000) Concepts of genetics, 6th Edition. Prentice Hall International, New Jersey, USA. pp: 713-733 [ Links ]
Kovach WL (1999) MVSP-A multivariate statistical package for windows, ver. 3.1. Kovach computing services, Pentraeth, Wales, UK. 137p. [ Links ]
Kumar LS (1999) DNA markers in plant improvement: An overview. Biotechnology Advances 17: 143-182. [ Links ]
Labra M, Grassi F, Bardini M, Imazio S, Guiggi A, Citterio S, Banfi E, Sgorbati S (2003) Genetic relationships in Opuntia Mill, genus (Cactaceae) detected by molecular marker. Plant Science 165: 1129-1136. [ Links ]
Luna-Páez A, Valadez-Moctezuma E, Barrientos-Priego AF, Gallegos-Vázquez C (2007) Caracterización de Opuntia spp. mediante semilla con marcadores moleculares RAPD e ISSR y su posible uso para diferenciación. Journal of the Professional Association for Cactus Development 9: 43-59. [ Links ]
Lyra MCCP, Santos DC, Mondragon-Jacobo C, Silva MLRB, Mergulhao ACES, Martínez-Romero E (2013) Molecular characteristics of prickly-pear cactus (Opuntia) based on internal transcribed spacer sequences (ITS) of Queretaro State-Mexico. Journal of Applied Biology & Biotechnology 1: 6-10. [ Links ]
Maddams WF (1972) The problem of variability. Cactus and Succulent Journal Great Britain 34: 74-78. [ Links ]
Majure LC, Puente R, Griffith MP, Judd WS, Soltis PS, Soltis DE (2012) Phylogeny of Opuntia s.s. (cactaceae): clade delineation, geographic origins, and reticulate evolution. American Journal Botany 99: 847-864. [ Links ]
Maki-Díaz G, Peña-Valdivia CB, García-Nava R, Arévalo-Galarza ML, Calderón-Zavala G, Anaya-Rosales S (2015) Características físicas y químicas de nopal verdura (Opuntia ficus-indica) para exportación y consumo nacional. Agrociencia 49: 31-51. [ Links ]
Mihalte L, Sestras RE, Gyorgy-Feszt G, Tamas E (2011) Assessment of genetic variation on four genera of Cactaceae using taxonomic, cytological and molecular markers methods. Plants Omics Journal 4: 142-148. [ Links ]
Mondragón JC (2003) Caracterización molecular mediante RAPDs de una colección de nopal de (Opuntia spp. cactaceae) del centro de México, como base del mejoramiento genético. Revista Chapingo Serie Horticultura 9: 97-114. [ Links ]
Montiel D, Valadez-Moctezuma E, Palomino G, Bermudez MA, Fernández FJ (2012) DNA extraction from roots of xoconostle. Journal of the Professional Association for Cactus Development 14: 35-40. [ Links ]
Mukerji SK, Ting IP (1969) Malate dehydrogenase isoenzymes in green tissue of Opuntia: isolation and characterization. Archives of Biochemistry and Biophysics 131: 336-351. [ Links ]
Murphy WR, Sites WJ, But GD, Haufler HC (1990) Proteins: isozyme electrophoresis. In: Hillis, DM, Mortis C (Eds). Molecular systematics. Sinaver Associated Inc. Publish. Massachusetts. USA. pp. 45-126. [ Links ]
Parish J, Felker P (1998) Fruit quality and production of cactus pear (Opuntia spp.) fruit clones selected for increased frost hardiness. Journal of Arid Environments 37: 123-143. [ Links ]
Parra F, Pérez-Nasser N, Lira R, Pérez-Salicrup D, Casas A (2008) Population genetics and process of domestication of Stenocereus pruinosus (Cactaceae) in the Tehuacan Valley, Mexico. Journal of Arid Environments 72: 1997-2010. [ Links ]
Peña-Valdivia CB, Luna-Cavazos M, Carranza-Sabas JA, Reyes-Agüero JA, Flores-Hernández A (2008) Morphological characterization of Opuntia spp: A multivariate analysis. Journal of the Professional Association for Cactus Development 10: 1-21. [ Links ]
Reyes-Agüero JA, Aguirre-Rivera JR, Flores-Flores JL (2005) Variación morfológica de Opuntia (cactaceae) en relación con su domesticación en la altiplanicie meridional de México. Interciencia 30: 476-484. [ Links ]
Samah S, Valadez-Moctezuma E (2014) Morphological seeds descriptors for characterize and differentiate genotypes of Opuntia (Cactaceae, Opuntioideae). Annual Research & Review in Biology 4: 3791-3809. [ Links ]
Samah S, Ventura-Zapata E, Valadez-Moctezuma E (2015) Fractionation and electrophoretic patterns of seed protein of Opuntia genus. A preliminary survey as a tool for accession differentiation and taxonomy. Biochemical Systematics and Ecology 58: 187-194. [ Links ]
Sánchez J J, Goodman MM, Stuber CW (2000) Isozymatic and morphological diversity in the races of maize in Mexico. Economic Botany 54: 43-59. [ Links ]
Sánchez-Mejorada H (1982) Consideraciones generales sobre la clasificación de las cactáceas. Cactáceas y Suculentas Mexicanas 27: 3-9. [ Links ]
Schneider L (1999) Opuntia albicar Scheinvar, una nueva especie para la ciencia del Estado de México. Revista del Jardín Botánico Nacional XX: 167-169. [ Links ]
Scheinvar L, Olalde-Parra G, Olvera-Shule D, Martinez-Gonzalez C (2010) Base de datos de especies silvestres de nopales Mexicanos. Laboratorio de Cactologia, Jardín Botánico IBUNAM. http://www.unibio.unam.mx/minero/index.jsp?accion=sc&colecciones=MEXU,Herbario. Fecha de consulta 9 diciembre de 2013. [ Links ]
Shilpha J, Silambarasan T, Pandian SK, Ramesh M (2013) Assessment of genetic diversity in Solanum trilobatum L., an important medicinal plant from South India using RAPD and ISSR markers. Genetic Resource Crop Evolution 60: 807-818. [ Links ]
Stuber CW, Goodman MM, Moll RH (1982) Improvement of yield and ear number resulting from selection at allozyme loci in a maize population. Crop Science 22: 737-740. [ Links ]
Taylor CL, Barker NP (2012) Species limits in Vachellia (Acacia) karroo (Mimosoideae: Leguminoseae): Evidence from automated ISSR DNA "fingerprinting". South African Journal Botany 83: 36-43. [ Links ]
Torres AM, Bergh BO (1980) Fruit and leaf isozymes as genetic markers in avocado. Journal of the American Society of Horticultural Science 105: 614-619. [ Links ]
Uzun I (1997) Fruit and cladode isozymes in cactus pear. Acta Horticulturae 438: 45-53. [ Links ]
Valadez-Moctezuma E, Samah S, Luna-Paez A (2014a) Genetic diversity of Opuntia spp. varieties assessed by classical marker tools (RAPD and ISSR). Plant Systematic Evolution 301: 737-747. [ Links ]
Valadez-Moctezuma E, Ortiz-Vásquez Q, Samah S (2014b) Molecular based assessment of genetic diversity of xoconostle accessions (Opuntia spp.). African Journal of Biotechnology 13: 202-210. [ Links ]
Valdez-Cepeda RD, Blanco-Macias F, Gallegos-Vazquez C (2003) Ordering and numerical classification in prickly pear cactus using fruit attributes. Revista Chapingo Serie Horticultura 9: 81-95. [ Links ]
Wang X, Felker P, Burow DM, Paterson HA (1998) Comparison of RAPD marker patterns to morphological and physiological data in the classification of Opuntia accessions. Journal of the Professional Association for Cactus Development 3: 3-14. [ Links ]
Weeden NF, Lamb RC (1985) Identification of apple cultivars by isozyme phenotypes. Journal of the American Society of Horticultural Science 110: 509-515. [ Links ]
Yudina RS (2012) Malate dehydrogenase in plants: Its genetics, structure, localization and use as a marker. Advances in Bioscience and Biotechnology 3: 370-377. [ Links ]
Zoghlami N, Chrita I, Bouamama B, Gargouri M, Zemni H, Ghorbel A, et al. (2007) Molecular based assessment of genetic diversity within barbary fig (Opuntia ficus-indica L. Mill.) in Tunisia. Scientia Horticulture 113: 134-141. [ Links ]

