










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkEcosistemas y recursos agropecuarios
versión On-line ISSN 2007-901Xversión impresa ISSN 2007-9028
Ecosistemas y recur. agropecuarios vol.2 no.6 Villahermosa sep./dic. 2015
Artículo científico
Análisis genético y bromatológico de mutantes de manzano (Malus x domestica Borkh) del cultivar golden delicius
Genetic and bromatological analyses of apple mutants (Malus x domestica Borkh) from golden delicious cultivar
1Mayra Alejandra Escobar-Saucedo, 1Marisol Cruz-Requena, 1Héctor Flores-Chávez, 2Alfonso Reyes-López, 1Cristóbal Aguilar-González, 1*Raúl Rodríguez-Herrera
1 Departamento de Investigación en Alimentos. Facultad de Ciencias Químicas. Universidad Autónoma de Coahuila. Blvd. Carranza and Ing. José Cárdenas V. s/n. Col. República Ote. Saltillo 25280, Coahuila. rrh961@hotmail.com
2 Universidad Autónoma Agraria Antonio Narro
Artículo recibido el 11 de marzo de 2011
Aceptado el 24 de febrero de 2015
RESUMEN
El manzano (Malus x domestica Borkh) es un frutal importante en el sur de Coahuila, México; que para su floración necesita la acumulación de unidades frío (UF), las cuales no se logran cubrir con las UF de la región, lo que ocasiona baja producción. En los últimos años en la región se han detectado mutaciones que necesitan menores UF. El objetivo del presente trabajo fue determinar diferencias genéticas y bromatológicas en los mutantes del sur de Coahuila. El análisis bromatológico mostró que el mutante Vigas I presentó el menor contenido de fibra (4.91 g) y el mayor contenido de azúcares (10.73 g), lo que le da ventajas comerciales. El análisis de polimorfismos en la longitud de fragmentos amplificados (AFLP's), mostró un mayor patrón de bandeado con la combinación: [EcoRI (+ACC +AGG) / Msel (+CTA)]. Obteniéndose con esta combinación 123 bandas amplificadas, las cuales fueron 100 % polimórficas. El mutante más parecido al control fue Vigas II con una distancia genética de 0.16 unidades, mientras que el mutante con mayor diferencia fue Primicia. Los mutantes estudiados pueden ser una alternativa a la problemática de UF de algunas regiones manzaneras de país.
Palabras clave: Brotador, Primicia, unidades frío, verde, vigas I-IV.
ABSTRACT
The apple tree (Malus x domestica Borkh) is an important fruit tree in southern Coahuila, Mexico. For flowering, it requires an accumulated number of cold air units (CAUs) which the CAUs of the region do not achieve to reach, leading to a low production. Mutations have been detected in the region that require less CAUs. The objective of this work was to determine the genetic and bromatological differences of the southern Coahuila mutants. The bromatological analysis showed that the mutant Vigas I has a lower fiber content (4.91 g) and the highest sugar content (10.73 g), which provide it with marketing advantages. The analysis of amplified length fragment polymorphisms (AFLPs), showed a larger banding pattern with the combination [EcoRI (+ACC +AGG) / Msel (+CTA)]. This combination yielded 123 amplified, 100 % polymorphic bands. The mutant closer to the control was Vigas II, with a genetic distance of 0.16 units, while the mutant with the greatest difference was Primicia. The studied mutants might be an alternative to the CAU issue in some apple tree regions of the country.
Key words: Brotador, Primicia, Cold air units, verde, vigas I-IV.
INTRODUCCIÓN
La producción de manzana es de gran importancia en la sierra de Arteaga, Coahuila, México; donde genera grandes beneficios económicos (Ramírez-Rodríguez y Cepeda-Si11er 1993). En Arteaga el principal cultivar de manzano es Golden Delicious, el cual requiere de 900 a 1200 UF para presentar una adecuada floración (Contreras de la Reé y Vázquez-Ramos 2010). El frío, es un factor determinante para que los árboles rompan el endo-letargo (Hernández-Herrera et al. 2006); sin embargo, en esta región frutícola sólo se acumulan entre 350 y 500 UF en la temporada invernal. Lo cual, se debe a que durante el invierno las noches son frías y los días cálidos y soleados, lo que ocasiona el calentamiento de los árboles, con efectos negativos en el letargo (Ramírez-Rodríguez y Cepeda-SiIler 1993). Además de una brotación y floración deficiente, lo que causa producción baja, si no se compensa la falta de UF con aceites vegetales o reguladores de crecimiento (Contreras de la Reé y Vázquez-Ramos 2010).
Una mutación es un cambio heredable en el material genético, que puede manifestarse en una o varias características morfológicas o fisiológicas de una especie (Schleif 1993). Estas mutaciones se estudian por medio de técnicas de marcadores moleculares (Azofeifa-Delgado 2006). La selección del tipo de marcador depende de los objetivos del estudio, de la especie de interés, y de la disponibilidad de recursos técnicos y financieros (Agarwal et al. 2008). Los polimorfismos en la longitud de fragmentos amplificados (AFLP's) son uno de los marcadores moleculares más utilizados en los análisis de variabilidad genética e identificación de genotipos, por su efectividad para detectar marcadores polimórficos con alta reproducibilidad, bajo costo y una mayor accesibilidad (Muzher et al. 2007). Se han detectado mutaciones del cultivar Golden Delicious en la sierra de Arteaga, Coahuila, las cuáles se caracterizan por presentar floración temprana, mayor producción, menos afectadas del paño o russeting, además de mayor vida de anaquel (Ramírez-Rodríguez y Cepeda-SiIler 1993). A estos mutantes se les han dado nombres regionales relacionados con el lugar donde se identificaron o por alguna característica morfológica, así han surgido nombres como Vigas I, II, III, IV, Brotador, Verde, Primicia, entre otros (Contreras de la Reé y Vázquez-Ramos 2010). Los cuales tienen la ventaja de que fueron seleccionados por los productores de acuerdo a su experiencia y necesidades, además de resover la problemática de la deficiencia de UF, lo que ha cauado que incremente su superficie de siembra (Ramírez-Rodríguez y Cepeda-SiIler 1993, Contreras de la Reé y Vázquez-Ramos 2010). Por lo anterior, el objetivo del presente estudio fue realizar el análisis genético y bromatológico de mutantes de manzano y del cultivar Golden Delicious.
MATERIALES Y MÉTODOS
Material vegetal
Se recolectaron al azar muestras de hojas jóvenes y de frutos de árboles de mutantes de manzano y del cultivar Golden Delicious. En la huerta Ingeniero Recio en Los Lirios, se recolectó el muíante Vigas I. Mientras que en la huerta Doctor Huereca, de San Antonio, se recolectó el muíante Vigas II y Golden Delicious. En tanto que en el poblado de San Antonio de las Alazanas se recolectaron los mutantes Primicia y Brotador. En tanto que en la huerta Víctor Sánchez de la localidad de Jamé, se recolectó el muíante Vigas III y en la huerta Nacho González el muíante Vigas IV y Verde. De los mutantes Vigas I, II, III y del cultivar Golden Delicious se colectaron hojas y frutos, mientras que de los mutantes Vigas IV, Primicia y Verde sólo se colectaron hojas.
Análisis bromatológicos
El análisis bromatológico se realizó en los mutantes Vigas I, Vigas II, Vigas III y Golden Delicious. Debido a que los mutantes tienen diferentes requerimientos de UF y maduran en diferentes fechas, fue difícil colectar frutas en similares condiciones de madurez de Primicia, Verde y Vigas IV. En este estudio se determinó el contenido de humedad por el método 934-01 (AOAC 1984), azúcares totales por la técnica de antrona (Kurnerth and Youngs 1984), y reductores por el método de fenol-sulfúrico (Dubois et ai 1956), proteínas de acuerdo con el método 467133, fibra cruda por el método 1-11, lípidos con la técnica 7-062, cenizas con el método 923-03, calcio, fósforo y sodio por el método 984.27, todos de la AOAC (1984). La unidad experimental fue de 100 g de fruto (pulpa y cáscara) y los tratamientos se distribuyeron bajo un diseño de bloques completos al azar con tres repeticiones. Para las diferentes variables, se realizaron los análisis de cada repetición en diferente día, considerando a los días como repeticiones. Con los datos se realizó un análisis de varianza (ANOVA), para las variables de proteínas y humedad, los datos se transformaron por medio de arcosen  antes de realizar el análisis de varianza. Cuando el ANOVA reporto diferencias entre tratamientos, los promedios de los tratamientos se compararon usando la prueba de rango múltiple de Tukey. Todos los análisis se realizaron con el software SAS® versión 9.0.
antes de realizar el análisis de varianza. Cuando el ANOVA reporto diferencias entre tratamientos, los promedios de los tratamientos se compararon usando la prueba de rango múltiple de Tukey. Todos los análisis se realizaron con el software SAS® versión 9.0.
Extracción de ADN
Las hojas recolectadas se lavaron con agua destilada para eliminar el polvo, la extracción de ADN se realizó con el método descrito por Della-porta et al. (1983). Una vez extraído el ADN se observó su integridad en un gel de agarosa al 1 %, con un transiluminador UV Spectroline® y mini cuarto oscuro marca UVP®. Posteriormente, se determinó la concentración de ADN con un Fluorómetro Turner BioSystems (TBS-380).
Polimorfismos en la longitud de fragmentos amplificados (AFLP)
Para la obtención de los AFLP's se empleó el protocolo de LI-COR® Biosciences, que se realizó en cuatro etapas, las cuales fueron: a) digestión del ADN genómico, en 1 µL de la mezcla de la enzima Msel de corte frecuente y de la enzima EcoRI corte poco frecuente, para ello las muestras se incubaron a 37 °C por 2 h. Para luego incubar de nuevo a 70 °C por 15 min, y posteriormente colocar en hielo, b) ligación de los adaptadores, a las muestras de ADN digerido se le agregaron 12 µ1 de la mezcla de los adaptadores específicos para cada extremo EcoRI (5'-CTCGTAGACTGCGTACC-3' y 3'-CTGACGCATGGTTAA-5'), Msel (5'-GACGATGAGTCCTGAG-3' y 3'-TACTCAGGACTC-AT-5'), y 0.5 µl de T4 ADN ligasa, en esta etapa la muestra se incubo durante 2 h a 20 °C. Para luego realizar una dilución 1:10 de la mezcla ligada, c) para la pre-amplificación, se agregaron primers de pre-amplificación AFLP® a la muestra. Posteriormente la muestra se sometió a reacción en cadena de la polimerasa (PCR) por 20 ciclos, donde cada ciclo consto de 30 s a 94 °C, 1 min a 56 °C, y 1 min a 72 °C, para luego mantener la muestra a 4 °C por 15 min. Finalizando con la dilución 1:40 de la muestra; d) selección de primers para la amplificación selectiva AFLP®, para la selección, se probaron 12 combinaciones de primers (Tabla 1) del IRDye Fluorescent AFLP Kit for Large Plant Genome Analysis, en este caso para cada combinación se utilizaron un primer Msel y dos EcoRI marcados con fluorescencia.
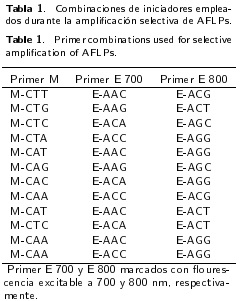
Para realizar la amplificación selectiva AFLP®, se siguió el protocolo PCR dúplex, un primer Msel con dos primers EcoRI (Tabla 1). En esta etapa se realizó una PCR utilizando un programa touchdown: un ciclo (30 s a 94 °C, 30 s a 65 °C y 1 min a 72 °C), y 12 ciclos (30 s a 94 °C, 30 s a 65 °C, con un decreciente de 0.7 °C por ciclo, y 1 min a 72 °C), y 23 ciclos (30 s a 94 °C, 30 s a 56 °C y 1 min a 72 °C), finalizando con temperatura de 4 °C. A cada tubo con las muestras se les adicionaron 5.0 µL de Blue stop solution, para después desnaturalizar las muestras, a 94 °C por 3 min y posteriormente fraccionar mediante electroforesis con geles de poliacrilamida al 6.5 %, con parámetros de la electroforesis de 1371 V, 29.1 mA a un temperatura de 53.8 °C.
Análisis de patrones de bandeo de AFLPs
Las bandas generadas por AFLPs a partir del ADN de los mutantes de manzano y el control, se codificaron como 0 y 1 (ausencia y presencia, respectivamente). Con los datos codificados se realizaron los siguientes análisis: diversidad genética, índice de contenido polimórfico, heterocigosis insesgada de Nei, número promedio de alelos, y número de alelos efectivos. Además, con la información de las bandas amplificadas de cada genotipo de manzano se construyó un dendrograma, para ello se empleó la distancia euclideana, debido a que fue la que presentó la mayor correlación cofenética con un valor de 0.981. Todos los análisis se realizaron con el software Info-Gen (Balzarini et al. 2009).
RESULTADOS
Análisis bromatológicos
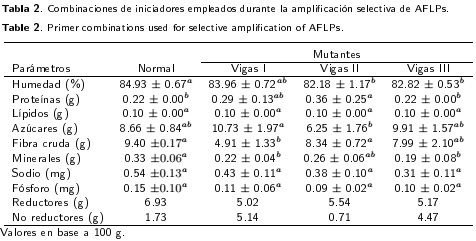
En la tabla 2 se muestra la caracterización bromatológica de los frutos de los mutantes de manzano y el control. Sólo se observaron diferencias en algunas variables bromatológicas de los diferentes mutantes con respecto al control, a pesar de que los frutos se recolectaron de diferentes huertos. El contenido de calcio del control fue mayor a la de los mutantes Vigas I, II y III. Mientras que el contenido de humedad y minerales de los frutos de Golden Delicious fue mayor que los de los mutantes Vigas II y Vigas III, y el contenido de fibra cruda del control sólo fue mayor que el contenido del mutante Vigas I. Por otra parte, los frutos del mutante Vigas II presentaron menor contenido de azúcares (6.25 g), mientras que los frutos de Vigas I presentaron el menor contenido de fibra (4.91 g).
Genotipificación de mutantes
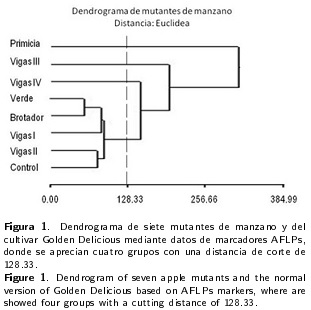
En el análisis de AFLP se probaron 12 combinaciones de primers (Tabla 1) de las cuales se seleccionó la combinación [EcoRI (+ACC +AGG) + Msel (+CTA)] debido a que con ella se obtuvo la mayor cantidad de bandas con mayor intensidad. Con esta combinación de primers se produjo un total de 123 bandas polimórficas. En el Dendrograma se observó que el mutante primicia presentó la mayor distancia genética (0.67) con respecto al control. Por otro lado, Vigas II fue el mutante que presentó la distancia más corta con respecto al control (0.16), lo que indica una mayor similitud entre ellos, esto puede ser debido a que ambas muestras presentan regiones similares del ADN, entre las cuales pudieran estar aquellas que condicionan las características bromatológicas del fruto y los requerimientos de frío (750 y 900 UF para Vigas II y Golden Delicious, respectivamente). Se observó que la diversidad genética fue de 0.50, esta mínima diferencia se atribuye a que todos los mutantes provienen de un mismo cultivar y que los genes que codifican para estas diferencias están asociados con la mutación. La heterocigosis insesgada de Nei tuvo un valor de 0.67, mientras que el número de alelos por locus fue de dos.
DISCUSIÓN
Análisis bromatológicos
Los frutos de los mutantes tienen contenidos similares de lípidos, azúcares, sodio y fosforo que el cultivar Golden Delicious. El contenido de calcio, humedad y minerales de Golden Delicious fue superior al de los mutantes. La similitud en las características bromatológicas de los mutantes de Vigas I-III con los frutos del cultivar Golden Delicious, les pueden dar ventajas comerciales y agronómicas, ya que requieren entre 750 y 800 UF para brotación, mientras que el cultivar Golden Delicious requiere entre 900 y 1200 UF, (Contreras de la Reé y Vázquez-Ramos 2010). Resultados similares fueron reportados por González-Portillo et al. (2008). El mutante Vigas I tuvo el porcentaje más alto de azúcares (Tabla 2), lo que pudede darle ventajas comerciales e industriales, ya que el contenido de azúcares determina el potencial de fermentación industrial. Al respecto Silveira et al. (2007) indican que dentro de los factores que determinan la cantidad de azúcares en la manzana, se encuentran el genotipo, la radiación solar y la humedad del suelo (Silveira et al. 2007).
El contenido de minerales (0.011 mg) en Golden Delicious fue mayor que el que presentaron los mutantes, aunque estos contenidos son inferiores a los reportados por Mancera-López et al. (2007), quienes indican que el contenido de calcio en la cáscara de manzana se encuentra entre 0.070 y 0.100 mg, mientras que en la pulpa oscila entre 0.025 y 0.050 mg. Altos contenidos de calcio y potasio dan mayor consistencia en la manzana, por lo que la falta de estos elementos, se vincula con desórdenes fisiológicos como corazón acuoso, mancha amarga y colapso interno del fruto, mientras que altos contenidos de calcio evitan la descomposición en postcosecha (Mancera-López et al. 2007).
Genotipificación de mutantes
En el análisis de AFLP's se probaron 12 diferentes combinaciones de primers (Tabla 1). El mayor número de bases amplificadas las tuvo la combinación EcoRI (+ACC +AGG) / Msel (+CTA), las cuales fueron 100 % polimórficas. Hay diferentes estudios donde se han utilizado combinaciones diferentes de primers, como el realizado por González-Portillo et al. (2008), quienes emplearon cuatro combinaciones de iniciadores, dos para la absorbancia de 700 nm y dos para 800 nm; con las cuales lograron detectar diferencias genéticas entre genotipos, lo que sugiere que la mejor combinación de primers está determinado por el genotipo, por lo que se deben probar diferentes combinaciones de iniciadores para identificar aquellas que ofrezcan mayor polimorfismo.
Se tuvo una una correlación cofenética de 0.981, lo cual indica una buena estructura jerárquica entre los genotipos estudiados (Zambrano et al. 2003). De acuerdo con las bandas AFLP's, los mutantes Verde y Brotador se unen a menor distancia que el resto de los genotipos, formado un grupo. Estos mutantes se encuentran a una distancia de 60.5, siendo esta la menor distancia entre todos los pares de genotipos, lo que indica que estas poblaciones tienen una similitud multivariada mayor que cualquier otro par de genotipos evaluados (Balzarini et al. 2010). A ese grupo se unen los perfiles de Vigas I, Vigas II y el cultivar Golden Delicious. Siendo Vigas II, el mutante con mayor similitud genética al cultivar Golden Delicious, Vigas también presento similitud a Golden Delicious en los contenidos de lípidos, azúcares, fibra, minerales, sodio y fosforo en el fruto. Los perfiles de los genotipos Vigas IV, Vigas III y Primicia se unen en forma decreciente a mayor distancia euclideana al grupo formado previamente. Siendo el mutante Primicia es el más alejado del resto de los genotipos evaluados, esta disimilitud puede deberse a que tiene un mayor número de mutaciones en su genoma y a las diferencias en brotación temprana, fruto grande y cónico, con mayor cantidad de lenticelas (Contreras de la Reé y Vázquez-Ramos 2010).
Por otra parte, si el corte de separación de grupos en el dendrograma (Figura 1) se realiza a una distancia euclideana de 128.33, se obtienen cuatro grupos, un grupo compacto formado por cinco genotipos los cuales se unen a una distancia de 128.33 y tres genotipos (Vigas IV, Vigas III y Primicia) que se alejan de éste grupo. Esta separación se realizó de esta forma dado que para definir el número correcto de grupos, no existen estadísticos exactos (Núñez-Colín y Escobedo-López 2011), solo algunas pruebas pseudoestadísticas como la t2 de Hotelling y el criterio cúbico de agrupación.
El análisis de las bandas de AFLP obtenidas, indicó que 100 % de los loci fueron polimórficos, por lo que son informativos y sirven para determinar la diversidad genética entre los genotipos evaluados. Se observó un valor de diversidad genética de 0.5, lo que sugiere que a pesar de que los mutantes provienen del cultivar Golden Delicious (Contreras de la Reé y Vázquez-Ramos 2010), tienen mutaciones que los diferencian entre si y de su contraparte normal. El género Malus se caracteriza por una gran diversidad, como resultado de la acumulación de mutaciones somáticas, que se incrementaron con la domesticación (Muzher et al. 2007). Para la heterocigosis insesgada de Nei se obtuvo un valor de 0.67 que es similar al valor reportado por Ott (1992) para considerar a un locus como altamente polimórfico. Por otra parte, se encontró un valor de dos para el número efectivo de alelos por locus, lo que sugiere la presencia de al menos dos alelos diferentes por locus en los mutantes estudiados. Sin embargo, para determinar el número de genes involucrados con estos mutantes, se requiere de estudios genéticos más detallados que involucren cruzas entre genotipos con altos requerimientos de frió y mutantes con bajos requerimientos y sus progenies (Rodríguez-Herrera et al. 2007), estudios con diferentes generaciones de una cruza entre un genotipo con altos requerimientos de UF con uno de bajos refquerimentos (Rodríguez-Herrera et al., 2000), o estudios de secuenciación de la última generación, para determinar el número de genes involucrados en las mutaciones (Velasco et al. 2010). Se han realizado estudios de caracterización molecular y bromatológica de mutantes de manzano (González-Horta et al. 2005, González-Portillo et al. 2008, Contreras de la Reé y Vázquez-Ramos 2010).
CONCLUSIONES
El mutante Vigas I tuvo el menor contenido de fibra y el mayor contenido de azucares, lo cual indica que sus frutos podrían llegar a competir en el mercado con los frutos de Golden Delicious. Los marcadores moleculares permitieron determinar diferencias en el número y posición de las bandas entre los mutantes y el cultivar Golden Delicious, lo que sugiere que los genotipos evaluados son genotipos diferentes y únicos.
AGRADECIMIENTOS
Al SNI-CONACYT, 2009 (Genotipificación y caracterización bromatológica de tres mutantes de manzano) y Gobierno del Estado de Coahuila-FONCYT-COECYT-CONACYT-2013-2014 (Caracterización bromatológica, genética/epigenética de mutantes de manzano para bajos requerimientos de frió). Los autores MAES, MCR y HFC agradecen al CONACYT-México por el apoyo financiero recibido para la realización de estudios de Postgrado.
LITERATURA CITADA
Agarwal M, Shrivastava N, Padh H (2008) Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Reports 27: 617-631 [ Links ]
Azofeifa-Delgado Á (2006) Uso de marcadores moleculares en plantas; aplicaciones en frutales del trópico. Agronomía Mesoamericana 17: 221-242. [ Links ]
AOAC (1984). Fruits and fruit products. Official Methods of Analysis. 14 th edition. Association of Official Analytical Chemists. Washington, D.C. U.S.A. pp: 391-393. [ Links ]
Balzarini M, Bruno C, Peña A, Teich I, Di Rienzo J (2010) Estadística en biotecnología. Aplicaciones en Info-Gen. Encuentro Grupo Editor. Córdoba, Argentina. 228p. [ Links ]
Balzarini M, Arroyo A, Bruno C, Di Rienzo J (2009) Análisis de datos de marcadores con Info-Gen. Editorial Brujas. Córdoba, Argentina. 120p. [ Links ]
Contreras de la Reé FJ, Vázquez-Ramos JA (2010). Cultivares y mutantes de manzana para el sureste de Coahuila. Boletín técnico 14. INIFAP. Campo Experimental Saltillo. México. 22p. [ Links ]
Dellaporta SL, Wood J, Hicks JB (1983) A plant DNA minipreparation: Version II. Plant Molecular Biology Reporter 1: 19-21. [ Links ]
Dubois M, Gilíes KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related substances. Analytical Chemistry 28: 350-356. [ Links ]
González-Horta A, Fernández-Montes MR, Rumayor-Flores A, Castaño-Tostado E, Martínez-Peniche RA (2005) Diversidad genética en poblaciones de manzano en Querétaro, México revelada por marcadores RAPD. Revista Fitotecnia Mexicana 28: 83-91. [ Links ]
González-Portillo M, Rocha-Guzmán NE, Simpson J, Rodríguez-Guerra R, Gallegos-Infante JA, Delgado E, Gil-Vega K (2008) Determinación de algunos atributos de calidad de manzana Starking y Golden Delicious en comparación con sus mutantes y su relación con las unidades frío. Ciencia y Tecnología Alimentaria 6: 27-32. [ Links ]
Hernández-Herrera A, Zermeño-González A, Rodríguez-García R, Jasso-Cantú D (2006) Beneficios del encalado total del manzano (Malus x domestica Borkh) en la sierra de Arteaga, Coahuila, México. Agrociencia 40: 577-584. [ Links ]
Kunerth WH, Youngs VL. (1984) Modification of the anthrone, carbazole, and orcinol reactions for quantitation of monosaccharides. Cereal Chemistry 61:344 - 349. [ Links ]
Mancera-López MM, Soto-Parra JM, Sánchez-Chávez E, Yáñez-Muñoz R. M, Montes-Domínguez F, Balandrán-Quintana RR (2007) Caracterización mineral de manzana 'Red Delicious' y 'Golden Delicious' de dos países productores. Tecnociencia 1: 6-17. [ Links ]
Muzher BM, Younis RAA, El-Halabi O, Ismail OM (2007). Genetic identification of some Syrian local apple (Malus sp.) cultivars using molecular markers. Research Journal of Agriculture and Biological Sciences 3: 704-713. [ Links ]
Núñez-Colín CA, Escobedo-López D (2011) Uso correcto del análisis cluster en la caracterización de germo-plasma vegetal. Agronomía Mesoamericana 22: 415-427. [ Links ]
Ott J (1992) Strategies for characterizing highly polymorphic markers in human gene mapping. American Journal of Human Genetics 51: 283-290. [ Links ]
Ramírez-Rodríguez H, Cepeda-Siller M (1993) El manzano. 2a Edición, Editorial Trillas, México, D.F. 208p. [ Links ]
Rodríguez-Herrera R., Rooney WL., Rosenow DT, Frederiksen RA (2000) Inheritance of grain mold resistance in grain sorghum without a pigmented testa. Crop Science 40: 1573-78. [ Links ]
Rodríguez-Herrera R, Rooney WL, Aguilar-González CN, Quero-Carrillo AR (2007) Genetic control and heritability of resistance to grain mold in F2:5 sorghum families with non-pigmented testa. Plant Breeding and Seed Science 55: 33-43. [ Links ]
Schleif R (1993) Genetics and Molecular Biology. 2nd. Edition. The Johns Hopkins University Press Baltimore Maryland, USA. 715p. [ Links ]
Silveira AC, Sautter CK, De Tonetto Freitas S, Galletta G, Brackmann A (2007) Determinación de algunos atributos de calidad de la variedad Fuji y sus mutantes al momento de cosecha. Ciencia y Tecnología Alimentaria 27: 149-153. [ Links ]
Velasco R, Zharkikh A, Affourtit J, Dhingra A, Cestaro A, (2010) The genome of the domesticated apple (Malus x domestica Borkh.). Nature Genetics 42: 833-839. [ Links ]
Zambrano AY, Demey JR, Fuenmayor F, Segovia V, Gutiérrez Z (2003) Diversidad genética de una colección de yuca a través de marcadores moleculares RAPDs. Agronomía Tropical 53: 157-173. [ Links ]

