











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
La altitud influye en la temperatura y en la humedad ambiental y del suelo, lo que puede manifestarse como una reducción de los crecimientos anuales típicos de cada especie, reducir la supervivencia del renuevo, afectar la viabilidad de la semilla y menguar el vigor de la vegetación. Si los cambios son muy extremos o la capacidad adaptativa o de desplazamiento de la especie son muy bajas, incluso puede haber pérdida de la vegetación (Champo-Jiménez, Valderrama-Landeros, y España-Boquera, 2012; Gutiérrez y Trejo, 2014; Islas Báez et al., 2015; Villanueva-Díaz et al., 2017).
El desfase climático entre los rangos altitudinales y las diversas coberturas vegetales ocasionado por el calentamiento global puede traer repercusiones en la dinámica, distribución, salud, adaptabilidad y sostenibilidad de los diversos organismos y ecosistemas del planeta (Estrada-Contreras et al., 2015; Monterroso-Rivas, Gómez-Díaz y Tinoco-Rueda, 2013; Pérez-Miranda et al., 2014; Sáenz-Romero et al., 2016; Vose, Peterson y Patel-Weynand, 2012).
La temperatura disminuye, en promedio, entre 0.4 y 0.7 °C por cada 100 metros de incremento en la altitud, dependiendo de la humedad relativa del aire (Lamprecht, 1990; Sáenz-Romero et al., 2012a), lo que puede generar cambios dentro de una misma masa forestal a lo largo de su distribución vertical en el terreno. Aunado a esto, Sáenz-Romero et al. (2012a) concluye que, en el centro de México, el aumento de la temperatura promedio será de 1.5 °C para el 2030, 2.3 °C para el 2060 y hasta 3.7 °C para el 2090; y que la precipitación se reducirá 6.7, 9.0 y 18.2 %, para los mismos periodos, respectivamente, intensificando estos cambios altitudinales.
Es importante estudiar las consecuencias del cambio climático en los ecosistemas forestales, como son: el estrés hídrico, mayor incidencia de plagas y enfermedades, declinación forestal, frecuencia e intensidad de incendios, erosión del suelo, reducción del crecimiento forestal y la regeneración natural. Ya que, en su conjunto, son consecuencias de afectaciones ecosistémicas anteriores, y también intensifican las tasas de degradación y deforestación actuales, lo que las transforma de ser efectos, a ser una parte causal y dinámica de la problemática ambiental (Comisión Nacional Forestal [Conafor], 2013a; Granados-Sánchez y López-Ríos, 2001; Pardos, 2010).
Una especie que resulta frecuentemente afectada por los fenómenos causados por el incremento de la temperatura en el planeta es el Abies religiosa (Kunth) Schltdl. et Cham [oyamel], pues es parte de una comunidad vegetal que se asienta en zonas húmedas y con temperaturas bajas. Las afectaciones causadas por la modificación de las variables climáticas óptimas para este, o cualquier tipo de vegetación, se pueden manifestar a nivel molecular, morfológico y fisiológico, así como reducir su vigor y regeneración natural (Castellanos-Acuña et al., 2014; Flores-Nieves et al., 2011; Granados-Sánchez y López Ríos, 2001; Vose et al., 2012).
Starker Leopold (1950) clasifica a las comunidades de oyamel dentro de los bosques boreales por su similitud con los de coníferas de las partes septentrionales de Norteamérica, norte de Europa y Asia. De igual forma, Rzedowski (1978) menciona que las poblaciones de oyameles que actualmente se localizan en México se desplazaron desde el hemisferio norte. Critchfield (1984) propone que, durante los periodos glaciares, comunidades de este tipo de vegetación se trasladaron a los actuales desiertos del norte de México; finalmente se establecieron en las zonas altas, como sierras y formaciones volcánicas, donde las temperaturas tienden a disminuir por efecto de la altitud.
Se han presentado casos de pérdida de arbolado por declinación forestal prácticamente en bosques de todo el mundo, como lo mencionan Sáenz-Romero et al. (2016); particularmente, para la especie Abies religiosa, se reporta el caso del Parque Nacional Desierto de los Leones (Granados-Sánchez y López-Ríos, 2001; Castillo-Flores et al., 2013; Rodríguez-Trejo, 2014), y más específicamente, aunque menos severa, en las masas forestales de oyamel en el monte Tláloc en el Estado de México (Flores-Nieves et al., 2011).
Los cambios en el clima pueden representar una reducción de la superficie que cuente con las condiciones apropiadas para ciertos tipos de vegetación (Sáenz-Romero et al. 2012a); estos autores proyectan la posible variabilidad (la alza o la baja) de la superficie de nichos ecológicos, entre ellos la del oyamel. Sus estudios, realizados en la Reserva de la Biosfera de la Mariposa Monarca, concluyen que la superficie de distribución de esta especie disminuirá, aproximadamente, 69.2 % para el 2030 y 96.5 % para el 2090; también prevén un desfase altitudinal de su nicho ecológico respecto al clima prevaleciente de hasta 300 m para el 2030.
El oyamel, con un rango altitudinal de distribución entre las cotas 2 900 y 3 600 m s. n. m. en la zona oriente del Estado de México, por su cercanía con las zonas de la cumbre, donde incide más la radiación solar, no cuenta con superficies óptimas para desplazarse a alturas mayores. Esto, aunado a que tiene un rango climático de distribución limitado y, por tratarse de un ecosistema fragmentado, suma las condiciones para que en un panorama futuro (70-80 años) de incremento en el calentamiento global la especie sufra reducciones en su vigor y una menor regeneración natural, lo que podría, incluso, repercutir hasta en su prevalencia (Castellanos-Acuña et al., 2014; Pérez-Miranda et al., 2014; Sáenz-Romero et al., 2012b). La hipótesis de la que se partió plantea que la regeneración natural y el vigor del oyamel disminuyen en los rangos de distribución altitudinal más bajos, debido al incremento de la temperatura y a la disminución de la humedad ambiental y del suelo. Asimismo, se plantea que la regeneración natural incrementa conforme aumenta el rango altitudinal, hasta que los factores de la cumbre (exposición a mayor radiación solar, suelos someros, bajas temperaturas) limitan el establecimiento del oyamel.
Lo anterior lo afirman diversos estudios realizados en torno al cambio del clima, desde el Holoceno medio hasta la actualidad (Manzanilla-Quiñones, Martínez-Adriano y Aguirre-Calderón, 2019), y sus efectos potenciales en la distribución de la vegetación, creando desfases entre los nichos ecológicos y las variables climáticas, obligando a las especies de climas templados a desplazarse a zonas altas y con menores temperaturas (Castellanos-Acuña et al., 2014; Pérez-Miranda et al., 2014; Sáenz-Romero et al., 2012b).
Cabe mencionar que, en la zona de estudio, el patrón de distribución de las distintas comunidades de vegetación está determinado, principalmente, por el gradiente altitudinal y sus respectivos rangos de temperatura y precipitación (Sánchez-González y López-Mata, 2003).
El objetivo del presente estudio fue evaluar el vigor, la mortalidad y regeneración natural del oyamel, mediante variables dasométricas, sobre un gradiente altitudinal en la zona oriente del Estado de México.
METODOLOGÍA
Área de estudio
El estudio se realizó en el monte Tláloc, el cual cuenta con una altitud máxima de 4 125 m s. n. m. y se localiza en la Sierra de Río Frío junto con otras cimas como el Telapón (Sánchez-González y López-Mata, 2003). La Sierra de Río Frío se ubica en el extremo septentrional de la Sierra Nevada, cuyo parteaguas es la línea divisoria entre las cuencas hidrográficas del Pacífico (cuenca del Balsas) y del Golfo (cuenca del Pánuco). Cuenta con lomeríos y sierras con bosques de coníferas, encino y mixtos del centro de México, se encuentra cubierto por bosque templado de coníferas con predominancia de pinos (Pinus montezumae Lamb., Pinus hartwegii Lindl), encinos (Quercus rugosa Neé, Q. laurina Humb et Bonpl.), ailes (Alnus jorullensis Humboldt, Bonpland et Kunth, A. acuminata Humboldt, Bonpland et Kunth), cedro blanco (Cupressus lusitanica Mill.) y oyamel (A. religiosa) (Lomas-Barrié, Terrazas-Domínguez y Tchikoué Maga, 2005).
El monte Tláloc y cimas aledañas son producto de actividad volcánica que creó extensos abanicos volcánicos. La zona de estudio también abarca parte del área natural protegida con la categoría de Reserva Estatal Sistema Tetzcotzingo y se encuentra en las coordenadas extremas UTM Zona 14, 522568.43 latitud N, 2146384.596 longitud O y 528805.759 latitud N y 2153486.335 longitud O. En la Figura1 se presenta la macrolocalización de la poligonal del bosque de oyamel reportada en la Serie v del Instituto Nacional de Estadística y Geografía [Inegi] (2013), misma que se halla en la región occidente del monte Tláloc en Texcoco, Estado de México.

Nota: conjunto de datos vectoriales de uso del suelo y vegetación, Inegi (2013), y sitios de muestreo, los cuales se numeraron con la cota altitudinal y número consecutivo de SO a NE (p. ej. 3000-1).
Fuente: elaboración propia con base en Inegi (2013)
Figura 1 Macrolocalización de la poligonal de A. religiosa reportada en la Serie V
La zona de estudio presenta un clima de montaña C(E)(w2)(w)b(i)g, según la clasificación de Köppen modificada por Enriqueta García (2004); clima semifrío, subhúmedo, con porcentaje de precipitación invernal menor a 5 %. El verano es largo, isotermal y la temperatura más elevada se presenta durante abril, mayo y junio, antes del solsticio de verano; la temperatura media anual se encuentra por debajo de los 8 °C, con una precipitación anual entre 1 000 y 1 200 mm (Casa A., 1997, citado por Comisión Nacional para el Conocimiento y Uso de la Biodiversidad [Conabio], 2020a).
El área cuenta con un tipo de suelo Luvisol umbrihúmico (LVhhu) con textura media, poroso y permeable. Pertenece a la unidad hidrogeológica del Cenozoico Superior Volcánico (mioceno a reciente), con rocas volcánicas (lavas, brechas y tobas), principalmente basálticas y andesíticas con una permeabilidad media a alta (Marín-C y Torres-Ruata 1990, citado por Conabio, 2020b); un régimen de humedad de suelo ústico, con 180 a 270 días de humedad, a údico, con 270 a 330 días de humedad Inegi (2013).
En la zona de estudio los rangos altitudinales donde está establecido el bosque de oyamel van de los 3 100 a los 3 500 m s. n. m. (Sánchez, 2004), y cubre una superficie de aproximadamente 3709 ha (Inegi, 2013). El estudio se centró en una poligonal de una masa forestal de A. religiosa que abarca una superficie de 1 360.870 ha que va de los 2 900 a los 3 600 m s. n. m.
Diseño del muestreo
La evaluación de las condiciones de vigor forestal y regeneración natural del oyamel en la zona de estudio se realizó mediante muestreo forestal enfocado al arbolado adulto, joven y de renuevo, considerando las cotas altitudinales entre los 2 900 a los 3 600 m s. n. m. para la ubicación de los sitios de muestreo. Se empleó una modificación del método utilizado para bosques y selvas, especificado en el Manual y procedimientos para el muestreo de campo. Re-muestreo 2011, del Inventario Nacional Forestal y de Suelos, de la Comisión Nacional Forestal [Conafor] (2011) y la Metodología para realizar y presentar los informes de sobrevivencia inicial (ISI) de las plantaciones forestales comerciales (Conafor, 2013b). Se instrumentó un muestreo dasonómico aleatorio y estratificado, enfocado al oyamel, realizado entre noviembre de 2018 y abril de 2019.
En la Figura 2 se presenta la forma, dimensión y orientación de los sitios de muestreo empleados para bosques y zonas áridas (Conafor, 2011), la cual contempla el registro y medición de arbustivas y herbáceas, mientras que en esta investigación el muestreo se concentra solo en individuos de oyamel y se registran las asociaciones vegetales. La disposición en “Y” invertida de los sitios y subsitios se mantuvo, a excepción de los sitios cuadrados de 1 m²; también cambia la dimensión y distancia entre ellos, estableciéndolos como se indica en la Figura 3, así como las variables consideradas para la medición de los individuos, las cuales se establecen más adelante.
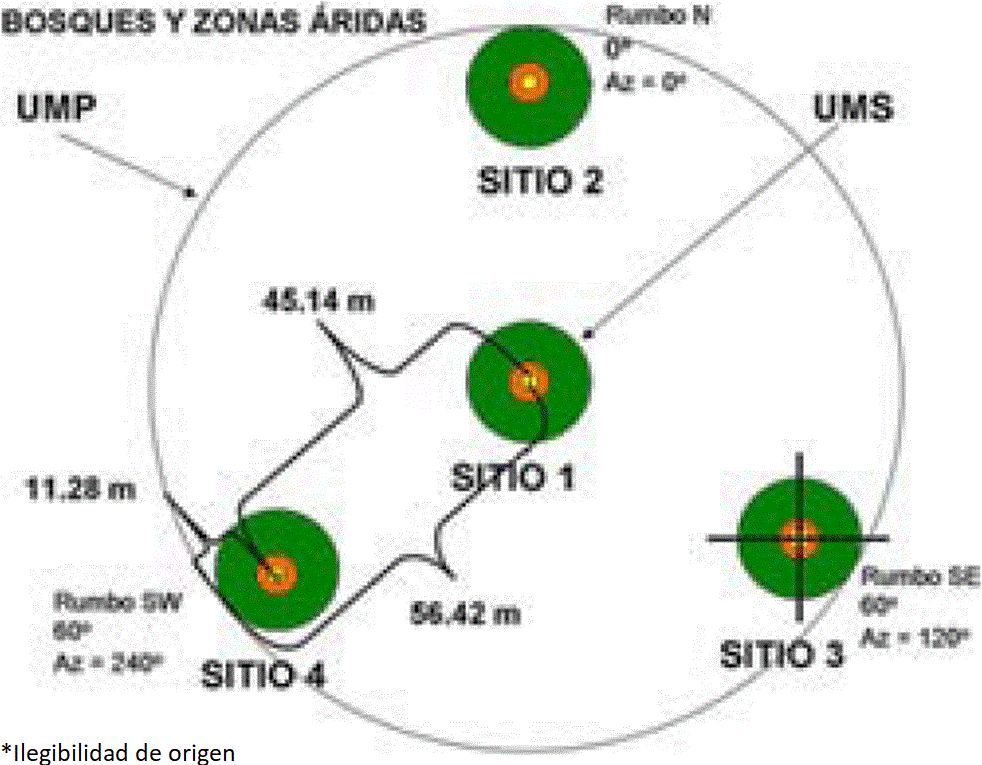
Nota: en los círculos verdes (400 m²) se mide el arbolado con DAP (diámetro a la altura del pecho) mayor a 7.5 cm. En los círculos naranjas (12.56 m²) se miden las plantas y árboles que tengan como mínimo 25 cm de altura y DAP menor a 7.5 cm. En los cuadros amarillos (1 m²) se registran hierbas, helechos, musgos y líquenes.
Fuente: Conafor (2011).
Figura 2 Forma de la unidad de muestreo utilizada

Nota: círculo mayor para arbolado adulto, círculos medianos para arbolado joven y círculos menores para renuevo.
Fuente: Conafor (2011).
Figura 3 Forma de los sitios de muestreo
De la Metodología para realizar y presentar los informes de sobrevivencia inicial (ISI) de las plantaciones forestales comerciales (Conafor, 2013b) se tomaron los sitios circulares de 100 m² para medir la regeneración natural de oyamel. De igual forma, se emplea una retícula de puntos para la distribución sistemática y aleatoria de los sitios. En dicha fuente se recomienda una intensidad de muestreo de 0.5 % para superficies mayores a 50 ha, pero debido a que la poligonal (objeto de estudio) comprende 1 360.870 ha, la intensidad de muestreo aplicada fue mayor a 0.1 %.
Distribución de muestras
Para la ubicación de los sitios de muestreo se empleó la herramienta digital ArcMap versión 10.0 de Environmental Systems Research Institute [ESRI] (2010). De las capas de uso de suelo y vegetación serie v (Inegi, 2013) se seleccionaron y recortaron las poligonales de oyamel ubicadas en la zona de estudio. La capa de curvas de nivel con cotas cada 20 m (de altitud) se extrajo del conjunto de datos vectoriales de información topográfica escala 1:50 000, serie iii, de la carta E14B31 (Inegi, 1982).
Con la herramienta Create Fishnet de ArcMap se distribuyó sobre la poligonal de oyamel una red de puntos equidistantes (600 m). Esta red se alineó perpendicularmente a la pendiente, lo que dio como resultado 39 sitios de muestreo (Figura 1). Cada punto de la red se alineó a la cota más cercana en el rango altitudinal de 2 900 a 3 600 m s. n. m. (una cota cada 100 m de altitud), se obtuvieron 8 cotas; cabe mencionar que se establecieron menos sitios en las cotas de los 2 900 y 3600 m, debido a que el área de distribución del oyamel a estas alturas es más reducida, restringiéndose sólo a aquellos sitios con características particulares, como son los fondos de cañadas, sitios con mayor contenido de humedad o menor exposición a la radiación solar.
Durante el muestreo se reubicaron algunos puntos buscando la cota y que no fueran sitios con condiciones particularmente húmedas favorecidas por la topografía, dando preferencia a laderas menos pronunciadas y más expuestas. Cinco puntos se desplazaron hacia un rango altitudinal mayor fuera de la poligonal, lo que amplía y ajusta el registro de su distribución; y cuatro más, hacia un rango menor fuera de la poligonal, uno de ellos por debajo de la cota 2 900 m s. n. m. hasta el límite inferior donde se observaron oyameles. En total se muestrearon 1.56 (ha) de las 1 360.870 ha del polígono (de oyamel) seleccionado para el presente estudio, lo que corresponde a una intensidad de muestreo de 0.1146 %.
Muestreo en campo
Para el muestreo forestal se clasificó al oyamel en tres estratos: arbolado adulto (estrato A), individuos mayores o iguales a 12.5 cm de diámetro a la altura del pecho (DAP); arbolado joven (estrato B), individuos menores a 12.5 cm. de DAP y mayores a 1 m de altura; y renuevo (estrato C), individuos menores o iguales a 1 m de altura (en esos casos su diámetro se midió en la base del tallo).
Unidades muestrales
El estrato A se muestreó en sitios circulares de 11.28 m de radio y 400 m² de área, donde a partir del punto central y en dirección al norte (0°) se midieron 7 metros para establecer el primero de los tres subsitios de muestreo del estrato B; el segundo 7 m al Sureste (120°) y el tercero 7 m hacia el Suroeste (240°), todos parten del centro del sitio de 400 m2. Los centros de los subsitios circulares de 5.64 m de radio y 100 m² de área del estrato B también fueron los centros de los subsitios de muestreo del estrato C, siendo estos circulares de 1.78 m de radio y 10 m² de área como se muestra en la Figura 3. Para los sitios de 11.28 m de radio se usaron cuerdas para compensar la pendiente del terreno.
Variables medidas
Los datos registrados para la descripción de los sitios fueron: número de sitio, coordenadas UTM, altitud (con geoposicionador satelital Garmin, modelo etrex Venture Cx), pendiente (con clinómetro), profundidad del suelo (con una varilla enterrada en el suelo), pedregosidad, exposición (con brújula), paraje y fecha del levantamiento.
Se utilizó un relascopio de Bitterlich [Pat NR. 172305] para determinar la altura de los árboles del estrato A y B; un calibrador vernier para medir los diámetros del estrato C, cinta diamétrica para medir el DAP de los estratos A y B, cinta métrica para medir alturas del estrato C, y cuerdas compensadas y estacas para delimitar los sitios de muestreo.
Aparte de las variables medibles ya mencionadas, se estimó de forma visual la pedregosidad del terreno (%), la densidad de la copa (%), el vigor (rango entre 3 y 1, 3 el más vigoroso), el estado sanitario de los árboles de los estratos A y B (rango entre 3 y 1, 3 el más sano), el color del follaje (3 verde obscuro, 2 verde claro-amarillento, 1 café), la sobrevivencia (por estrato) y la asociación vegetal.
Análisis estadístico
Los datos obtenidos del muestreo se procesaron en el programa de análisis estadístico SAS v. 9.4 del Statistical Analysis System [SAS]. Inicialmente se realizó una prueba de normalidad de los datos mediante Proc Univariate. Se realizaron tablas de frecuencia mediante Proc Freq, para observar las densidades de cada estrato y se empleó Proc GLM para establecer si había correlación entre las variables diámetro, altitud y densidad forestal (para los diferentes estratos) con la altitud.
Se usó la cota como variable clase y como variables dependientes la altura y diámetro de los individuos, los valores anteriores en escala logarítmica (AltLOG y DiamLOG), así como la densidad forestal, y se buscaron interacciones con la caracterización del vigor forestal y la mortalidad para los tres estratos.
RESULTADOS Y DISCUSIÓN
Se encontró oyamel fuera de las altitudes reportadas para el Valle de México por Rzedowski (1971) (2 700 a 3 500 m s. n. m.), desde los 2 850 en los fondos de las cañadas y hasta los 3 612, en una agrupación rodeada de pastos, establecida en un afloramiento rocoso (Sitio 3600-1).
La densidad de árboles de los tres estratos tiende a reducirse en las cotas inferiores e incrementa con la altitud y muestra una ligera disminución hacia las cotas más altas. Es nula la cantidad de oyamel registrada para el estrato C, en la cota inferior, en la cual se realizaron tres sitios ubicados a 2 850, 2 900 y 2 918 m s. n. m. (Figura 4), mientras que en la siguiente elevación (3 000 m s. n. m.) solo se registró un renuevo. En contraste, la cota 3 400 tuvo una densidad promedio de 186 800 /ha, es la altitud que presentó la mayor cantidad de oyameles del estrato C.
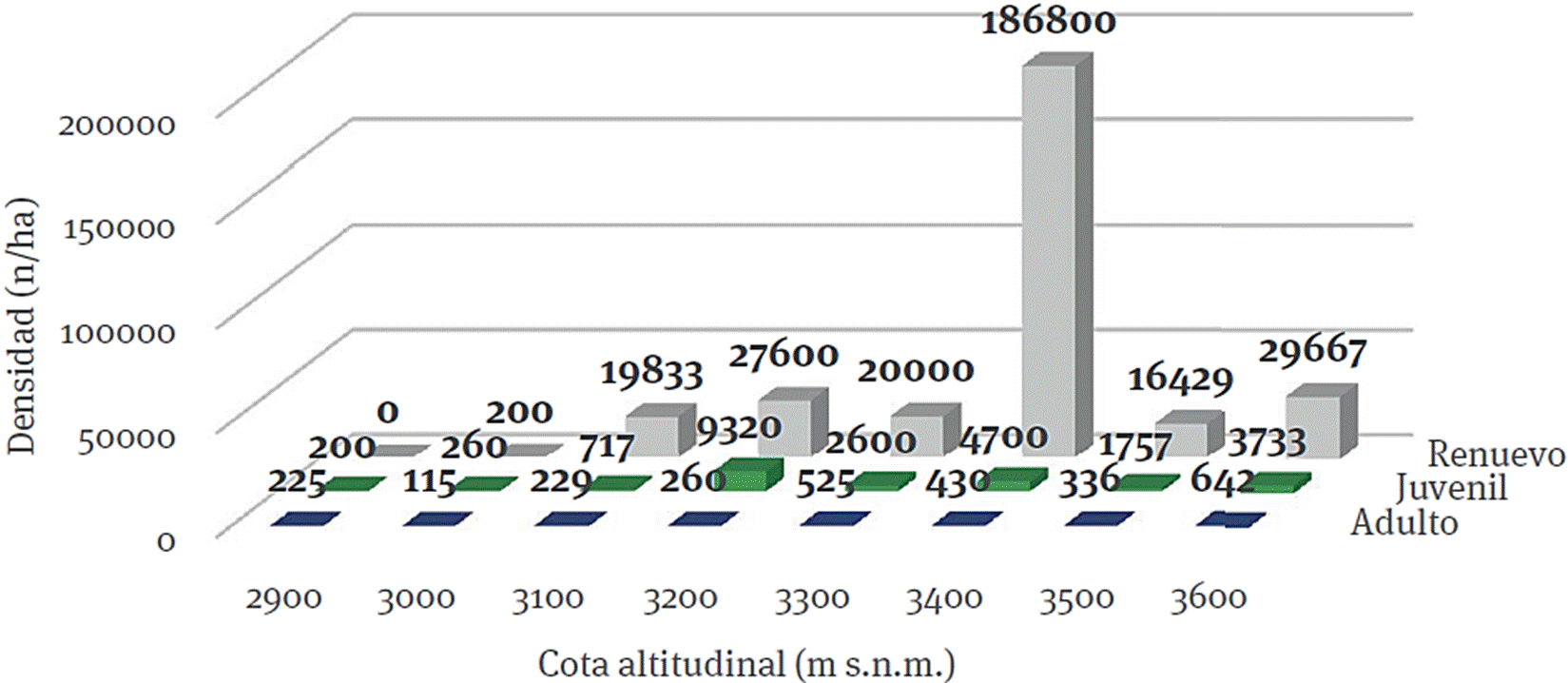
Los valores fuera de los rangos comunes observados para el arbolado juvenil y el renuevo corresponden a sitios alterados por el hombre; en el caso del estrato B, en la cota altitudinal de los 3 200 m s. n. m., se trata de un sitio (3200-5) donde ocurrió un incendio forestal hace algunos años, lo que permitió el establecimiento del renuevo y su posterior desarrollo a juveniles, favorecido por las características del lugar, un área bajo influencia de la sombra de un cerro aledaño y con mayor humedad del suelo, lo que dio lugar a juveniles dispersos de pino y oyamel que alcanzaron densidades de 41 000 /ha. Abundante regeneración post incendios catastróficos de la especie bajo estudio fue referida por Ángeles-Cervantes y López-Mata (2009) luego del incendio de 1998 en el Parque Nacional El Chico, Hidalgo.
Referente al estrato C, en la cota altitudinal de los 3 400 m s. n. m., el sitio 3400-5 presentó actividades de extracción maderable, lo que ocasionó que se abrieran algunos claros y se escarificara el suelo, eso facilitó el establecimiento del renuevo. Se registró una densidad de 823 000 /ha para ese sitio lo que elevó la densidad promedio para la cota 3 400 (186 800 /ha). El renuevo de oyamel se puede establecer bajo claros (principalmente de 44 a 88 m2 y 66 m2 en promedio) o bajo dosel (983 y 4 239 /ha respectivamente), como establecieron Lara-González, Sánchez-Velásquez y Corral-Aguirre (2009) para el Parque Nacional Cofre de Perote, Veracruz.
El Cuadro 1 presenta los porcentajes de vigor por estrato y rango altitudinal. En el estrato A se puede observar que la densidad se incrementa conforme aumenta la elevación y la cota que presenta una mayor proporción de individuos muertos es la 2 900, con 18.52 %, mientras que las que presentan una menor mortalidad en el arbolado adulto son las tres de mayor altitud, con 5.81, 6.38 y 5.19 %, respectivamente. De igual forma, el vigor del estrato A incrementa conforme aumenta el rango altitudinal.
Cuadro 1 Porcentajes de vigor por estrato y por cota altitudinal
| Cota (m s. n. m.) | Densidad de árboles (n/ha) | Vigor 3 (%) | Vigor 2 (%) | Vigor 1 (%) | Mortalidad (%) |
|---|---|---|---|---|---|
| Estrato A | |||||
| 2900 | 225 | 77.8 | 3.7 | 0.0 | 18.5 |
| 3000 | 115 | 52.2 | 34.8 | 4.3 | 8.7 |
| 3100 | 229 | 67.3 | 21.8 | 0.0 | 10.9 |
| 3200 | 260 | 88.5 | 1.9 | 0.0 | 9.6 |
| 3300 | 525 | 77.1 | 9.5 | 1.0 | 12.4 |
| 3400 | 430 | 82.6 | 11.6 | 0.0 | 5.8 |
| 3500 | 336 | 85.1 | 8.5 | 0.0 | 6.4 |
| 3600 | 642 | 92.2 | 2.6 | 0.0 | 5.2 |
| Estrato B | |||||
| 2900 | 200 | 33.3 | 50.0 | 0.0 | 16.7 |
| 3000 | 260 | 84.6 | 15.4 | 0.0 | 0.0 |
| 3100 | 717 | 90.7 | 9.3 | 0.0 | 0.0 |
| 3200 | 9,320 | 95.9 | 2.8 | 0.0 | 1.3 |
| 3300 | 2,600 | 50.0 | 33.1 | 6.9 | 10.0 |
| 3400 | 4,700 | 73.6 | 20.0 | 1.7 | 4.7 |
| 3500 | 1,757 | 40.7 | 27.6 | 13.0 | 18.7 |
| 3600 | 3,733 | 42.0 | 31.3 | 12.5 | 14.3 |
| Estrato C | |||||
| 2900 | - | - | - | - | - |
| 3000 | 200 | 100.0 | 0.0 | 0.0 | 0.0 |
| 3100 | 19,833 | 95.8 | 3.4 | 0.8 | 0.0 |
| 3200 | 27,600 | 95.7 | 4.3 | 0.0 | 0.0 |
| 3300 | 20,000 | 96.0 | 4.0 | 0.0 | 0.0 |
| 3400 | 186,800 | 87.3 | 10.4 | 1.3 | 1.1 |
| 3500 | 16,429 | 89.6 | 10.4 | 0.0 | 0.0 |
| 3600 | 29,667 | 92.1 | 6.7 | 0.0 | 1.1 |
Este mayor porcentaje de individuos muertos en las cotas bajas, así como la disminución del vigor forestal, se atribuyen a las condiciones de fragmentación y efecto borde, una mayor intensidad de vientos, caída de rayos, actividades antropogénicas, pero también en gran medida al cambio climático, que supone un incremento de las temperaturas y una reducción de la precipitación (Sáenz-Romero et al., 2012a), presentando escenarios de mayor sequía y teniendo repercusiones sobre el vigor y distribución de la vegetación debido a un desfasamiento de las variables climáticas y las variables requeridas para cada organismo.
En el estrato B el mayor porcentaje de individuos muertos y con menor vigor se registró para las cotas altas, esto se relaciona con actividades de extracción de recursos y también con las condiciones de los sitios, donde si la densidad del estrato A es alta, evita el paso de la luz, lo que dificulta el desarrollo de los individuos del estrato B, y se favorece el desarrollo de hongos y líquenes. Lo anterior también limita el establecimiento del sotobosque y del estrato C (Aunós, Martínez y Blanco, 2007; Manzanilla, 1974; Santillán-Peréz, 1991). Otra causa son sitios con suelos someros, pendientes pronunciadas y fuertes vientos que derriban a los individuos del estrato B.
Referente al estrato C, cabe destacar que el vigor disminuye hacia las cotas altas, siendo la cota 3 400 m la que presenta los porcentajes más bajos de vigor, posiblemente debido a la alta densidad del estrato A y a la competencia, pues también es una de las cotas con mayor cantidad de renuevo.
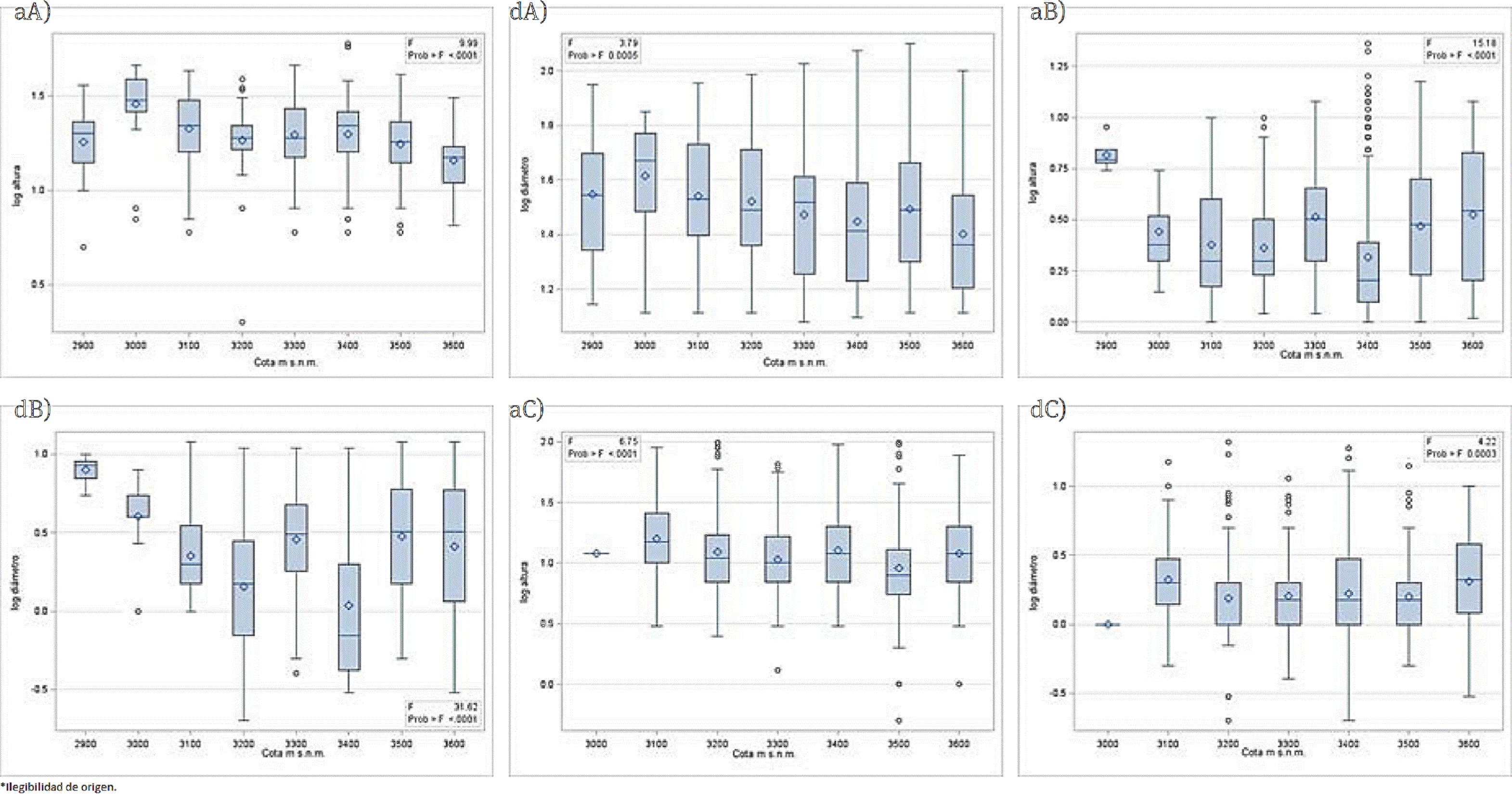
En la Figura 5 (aA) se observa la correlación entre la altura logarítmica del estrato A y la cota altitudinal, hay una estrecha correlación entre dichas variables, como resultado los individuos con mayor altura, en promedio, se presentaron en las cotas inferiores, en parte debido a que se trata de árboles dispersos con menor competencia, aunque las mayores alturas registradas por individuo se localizaron en la cota 3 400 m. Esta tendencia de reducción de altura al incrementar altitud también puede relacionarse con las menores temperaturas, profundidad del suelo y disponibilidad de humedad a mayores elevaciones, como Klinka et al. (1996) hallaron para Abies lasiocarpa (Hook.) Nutt. en un ambiente subalpino de la Columbia Británica, en los Estados Unidos.
Fuente: elaboración propia.
Figura 5 Correlación entre la altura (a) y diámetro (d) logarítmicos de los estratos A (superior), B (en medio) y C (inferior) y la cota altitudinal
La altura se reduce en las cotas superiores debido a diversos factores como el gradiente adiabático, una mayor densidad forestal y una menor profundidad del suelo. A manera de comparación, en la cota 3 300 m se registra una densidad promedio de 525 /ha, en la cota 3 400 m una densidad promedio de 430 /ha y en la cota 3 600 una densidad promedio de 642 /ha; mientras que en las cotas inferiores se registran densidades medias de 225/ha para los sitios de la cota 2 900 m, 115/ha para la cota 3 000 m y 229/ha para la cota 3 000 m y 229 /ha para la cota 3 100 m.
En la Figura 5 (dA) se muestra el efecto de la cota altitudinal sobre el DAP en escala logarítmica para el estrato A; como lo establece Colohua-Citlahua (2015), en las masas forestales del Parque Nacional Pico de Orizaba, con la proyección de modelos de densidad encontró que a medida que disminuye el número de árboles, aumenta la categoría diamétrica.
Para el estrato B, en la Figura 5 se muestra el efecto de la variable cota sobre las variables log altura y log diámetro (aB y dB), respectivamente. Se observa una menor correlación en comparación con el estrato A; para el caso de la altura se obtuvieron valores más bajos que para el DAP y, de manera similar, disminuyen los valores conforme se incrementa la cota altitudinal, con una correlación negativa más marcada para el DAP que para la altura; se limita su crecimiento por el dosel superior, la competencia con su propio estrato, el gradiente adiabático y contenido de humedad, así como el contenido de materia orgánica y pedregosidad en el suelo, denotando que es la cota 3 400 m la que presenta mayor cantidad de individuos en el estrato B, con un promedio de 4 700 /ha.
Esta disminución en los diámetros y alturas de los árboles, con relación al aumento en altitud, se observó de manera similar por Linares et al. (2015) en España, donde estudiaron los efectos del cambio climático sobre el crecimiento de A. pinsapo en el sur de la península ibérica y notaron que los individuos con diámetros mayores se localizan en los gradientes altitudinales inferiores, mientras que en el rango superior se encuentran individuos con diámetros menores a 10 cm, demostrando dendrocronológicamente que se han establecido más recientemente. También reportan una mayor área basal de individuos muertos en el rango inferior y que decrece conforme se incrementa la altitud.
En la Figura 5 (aC y dC) se muestra el efecto sobre las variables log altura y log diámetro para el estrato C, presentándose una disminución de los valores de estas variables conforme se incrementa la altitud, salvo en las cotas 3 400 y 3 600 m, donde aumenta el diámetro, lo que demuestra que se tiene una regeneración natural un poco más vieja o vigorosa hacia las partes altas.
Desde luego, puede haber influencia de la edad de la regeneración en sus dimensiones.
En la Figura 5 (aC y dC) la cota 3 600 m tiene uno de los valores más altos de logaritmo de altura, y el más alto para logaritmo de diámetro para el estrato C, la regeneración. Se trata de la segunda cota más elevada con mayor densidad de renuevo, como se verá más adelante. Las altas dimensiones de la regeneración a mayor altitud, dejan ver un ambiente favorable para establecerse donde debería haber limitaciones.
Estos resultados contrastan con los reportados por Sáenz-Romero, Beaulieu y Rehfeldt (2011), quienes recolectaron 14 poblaciones de Pinus patula en el estado de Oaxaca, México, a lo largo del rango altitudinal de 2 400 a 3 050 m s. n. m. con intervalos de 50 m de altitud. Las poblaciones de altitudes menores registraron en promedio plántulas más altas y de mayor peso que aquellas originadas a mayor altitud, con condiciones más extremas para este pino.
Si se considera la densidad del arbolado para el estrato A en relación a la elevación, se puede observar (Figura 6, izq.) que la densidad tiene una relación directa con la altitud hasta la cota 3 300 m, donde alcanzó la media más alta, se estabiliza para las siguientes dos cotas, y repunta en el límite superior con la densidad promedio por cota más alta con 642 /ha, aunque los sitios con una mayor densidad son el 3 400-1 con 950 /ha, el 3 500-7 con 900 /ha y dos sitios de la cota 3 600 con 875 /ha y 850 /ha.

Fuente: elaboración propia.
Figura 6 Gráfico de cajas con la interacción entre la densidad forestal del estrato A (izq.), B (centro) y C (dcha.) con la cota altitudinal
La densidad del estrato B, que se presenta en la Figura 6 (centro), se comportó de manera similar que el estrato A, es mayor conforme se aumenta la altitud, alcanza su valor más alto en promedio en la cota 3 400 m y después disminuye para volver a incrementar en la mayor elevación estudiada. Se destaca el valor extremo del sitio 3 200-5 en la cota 3 300 m, con una densidad de 41 000 /ha; si se excluye, los que presentan una mayor densidad son el 3400-5 con 15 000 /ha, 3 300-4 y 3 500-7 con 6 000 /ha cada uno y el 3 600-3L (límite superior) con 5 200 /ha; las densidades más bajas se registran en las cotas inferiores.
Para el estrato C (Figura 6, dcha.), no se registraron individuos en el límite inferior y sólo un individuo en la cota 3 000 m. Los valores de densidad de renuevo forestal más altos están en la cota 3 400 m en los sitios 3 400-5 y 3 400-3, con 823 000 y 94 000 /ha, pero los valores de la cota 3 100 m son importantes, debido a que es la menor elevación que presenta abundancia de renuevo, a una densidad de 71 000 /ha, mientras que, en el límite altitudinal superior, particularmente el sitio 3 600-1, registra una densidad de 57 000/ha.
El renuevo se puede establecer en las cotas inferiores, pero requiere de condiciones específicas para su establecimiento y desarrollo, como mayor concentración de humedad, menor exposición a la luz solar y estar fuera de zonas fácilmente accesibles para el ser humano.
En el Cuadro 2 se presentan los valores de F, p y r² de las correlaciones entre la altura y diámetro logarítmicos y la densidad de los tres estratos con la cota altitudinal.
Cuadro 2 Valores de F, p y r² para la altura y diámetro logarítmicos y la densidad por estrato
| Valor | Estrato A | Estrato B | Estrato C | |
|---|---|---|---|---|
| log altura | F | 9.99 | 15.18 | 6.75 |
| p | 0.0001 | 0.0001 | 0.0001 | |
| r² | 0.119 | 0.086 | 0.026 | |
| log diámetro | F | 3.79 | 31.62 | 4.22 |
| p | 0.0005 | 0.000 | 0.0003 | |
| r² | 0.049 | 0.164 | 0.016 | |
| densidad | F | 41.17 | 545.97 | 763.71 |
| p | 0.0001 | 0.0001 | 0.0001 | |
| r² | 0.36 | 0.773 | 0.754 |
Fuente: elaboración propia.
En las cotas inferiores (2 900 y 3 000 m s. n. m.) prácticamente no se encontró regeneración natural en los sitios muestreados, esto debido a la mayor presencia de actividades antropogénicas, pero también se muestrearon sitios de difícil acceso y poco perturbados donde tampoco se presentó regeneración natural, lo que se podría explicar por el desfase adaptativo del oyamel con las condiciones climáticas propicias, como sucede también con otras coníferas (Castellanos-Acuña et al., 2014).
El individuo más joven del estrato B registrado en la cota inferior tiene una altura de 5.5 m y un DAP de 8 cm, mientras que la altura promedio para este mismo estrato es de 6.6 m de altura y un DAP de 8.1 cm, por lo que no se ha registrado el establecimiento de regeneración natural desde que se establecieron los árboles del estrato B, presentes en la cota 2 900 m. Los individuos del estrato B alcanzan en promedio una altura de 3 m y un DAP de 4.4 cm para la cota 3000 m.
En la cota 3 400 m, donde el estrato A tiene una mayor densidad, las cantidades de renuevo se reducen, debido a lo expresado por Manzanilla (1974), quien menciona la importancia de una correcta dosificación de luz a través de podas por lo alto, para esta especie considerada tolerante a la sombra. Los rodales muy densos no presentan regeneración, y en rodales muy iluminados, la densidad del sotobosque compite con las plántulas de oyamel, siendo el óptimo de cobertura de sotobosque, para una mayor regeneración natural, de entre 41 y 81 %, según este último autor.
CONCLUSIONES
La cota altitudinal superior de la zona de estudio donde se registró al oyamel, es más alta que la referida por la literatura de hace algunas décadas, para dicha especie en el valle de México, lo que constituye una primera evidencia de que está migrando a sitios más altos a causa del cambio climático global.
La menor densidad en las cotas bajas, y la mayor densidad en las cotas media y alta, para los tres estratos de arbolado de oyamel (adultos, juveniles, y regeneración) estudiados, así como la elevada mortalidad para el estrato de arbolado maduro en la cota altitudinal más baja, constituyen otro indicio de que el oyamel se está desplazando a mayores altitudes, como respuesta al cambio climático. Otra evidencia que apunta a la misma conclusión, es el menor vigor que se observó en el arbolado maduro y juvenil, en las cotas más bajas.
Por otra parte, la altura y diámetro de los árboles maduros tienden a reducirse a mayores altitudes, pues a causa del gradiente adiabático se encuentran temperaturas más frescas en ellas, así como menor disponibilidad de agua por tender a congelarse. Asimismo, la competencia intraespecífica tiende a incrementar debido a la relación directa observada entre densidad de oyameles adultos y altitud.
La escarificación del piso forestal y la presencia de claros pequeños, producto de aprovechamiento forestal, han favorecido la regeneración de la especie bajo estudio. Las zonas no aprovechadas retienen más humedad y en ellas son más comunes musgos, helechos y hongos, que también son parte de la biodiversidad.
Es conveniente estudiar si otras especies relevantes de este ecosistema, y otros bosques de oyamel, también están migrando a mayores altitudes, así como el efecto de otros procesos ecológicos, como un diferente régimen de fuego y áreas con poca o mínima alteración antropógena.
Los resultados del presente trabajo, y observaciones en campo, permiten considerar algunas actividades que facilitarían tanto la migración como la migración asistida del oyamel a mayores altitudes. Se puede utilizar P. hartwegii para que actúe como especie nodriza, proporcionando sombra al oyamel en las localidades más altas. El apilamiento de rocas, materiales leñosos e incluso de residuos de aprovechamiento, generarían micrositios con mayor humedad en el piso forestal, para propiciar el establecimiento del oyamel. Asimismo, en varias zonas, en especial las de mayor altitud, puede pensarse en un control temporal del zacatón (Muhlenbergia spp.) para que tenga más éxito la regeneración natural del oyamel.

