











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En este trabajo se estudió el efecto de la temperatura y de las características de la semilla en la germinación de dos especies dominantes de diferentes matorrales xerófilos del valle de Tehuacán-Cuicatlán, Puebla:Dasylirion lucidum Rose y Beaucarnea gracilis Lemaire (Figura 1).

Nota: a) D. lucidum b) B. gracilis sin y con infrutescencias
Fuente: fotos por Dante A. Rodríguez T.
Figura 1 Imágenes de Dasylirion lucidum Rose y Beaucarnea gracilis Lemaire
D. lucidum
Los sotoles (Dasylirion spp.) son plantas que se distribuyen desde el sur de EE. UU. hasta Oaxaca, México. El género cuenta con 20 especies, todas presentes en el país, a lo largo de siete estados. La mayoría de estas especies son endémicas de México (Reyes-Valdés et al., 2012; Rivera-Lugo y Solano, 2012). Pertenecen a la familia Asparagaceae. Este género se encuentra en regiones de clima árido y semiárido, forma parte del matorral xerófilo (Rzedowski, 1998).
Dasyliron lucidum Rose, también conocido como sotolín, tehuizote o cucharilla, es una especie rosetófila acaule con tallos de hasta 1.5 m de alto, ocasionalmente ramificados. Es una especie amenazada, endémica del valle de Tehuacán-Cuicatlán, en Puebla y Oaxaca. Forma parte del matorral xerófilo ubicado entre 1500 y 2300 msnm, florece de mayo a junio y fructifica de abril a noviembre (Rivera-Lugo y Solano, 2012). Los Dasylirion son plantas dioicas (Reyes-Valdés et al., 2012), cuya semilla se encuentra en una cápsula trialada. Los Dasylirion del Valle de Tehucacán-Cuicatlán, antes eran considerados como D. acrotriche (Schiede) Zucc. Sin embargo, Rivera-Lugo y Solano (2012) establecen que se trata de D. lucidum, por ello, algunas de las referencias bibliográficas aquí utilizadas, de investigaciones realizadas en dicho valle, se consideraron para D. lucidum, si bien originalmente se refieren a D. acrotrhiche (Schiede) Zucc.
Los principales usos que se dan a D. lucidum en la región son: uso alimenticio y en ceremonias religiosas (Comisión Nacional de Áreas Naturales Protegidas [CONANP], 2013). La especie tiene importancia artesanal; se utilizan las porciones basales de las hojas para hacer arreglos florales con motivos religiosos, uso que ha contribuido en gran medida a la disminución de las poblaciones. Lo anterior, aunado a la baja germinación de las semillas en condiciones naturales, ha contribuido a que la especie se encuentre amenazada de acuerdo con la NOM-059 (Secretaría de Medio Ambiente y Recursos Naturales [SEMARNAT], 2010).
Existe escasa información referente a la capacidad germinativa de las semillas de esta especie y la ecología de la semilla ha sido poco estudiada. Asimismo, dada su dominancia en algunos matorrales xerófilos, se considera necesario estudiar su propagación para restaurar sus poblaciones en las áreas de distribución natural que han sido degradadas.
B. gracilis
El género Beaucarnea se distribuye desde el noreste de México hasta América Central. Cuenta con 11 especies, 10 de ellas se encuentran en México, de las cuales ocho son endémicas. En el Valle de Tehuacán-Cuicatlán, habitan tres especies, dos de ellas son endémicas (Hernández y Zamudio, 2003; Hernández et al., 2012; Rivera-Lugo y Solano, 2012). Este género pertenece a la familia Asparagaceae. Los nombres comunes de B. gracilis son “pata de elefante”, “sotolín” y “palma petacona”. Es una planta rosetófila arborescente, que alcanza entre 6 y 12 m de altura y sus tallos de 1.5 a 3 m de diámetro en la base, marcadamente ensanchados, globosos y hasta 2.5 m de altura. Corteza gruesa, gris a parda. Las hojas en rosetas apicales, con láminas de 24 a 68 cm de longitud. Plantas dioicas a poligamodioicas. Inflorescencias en panículas largamente ovoides, de 0.6 a 1.5 m de longitud, color anaranjado a amarillo pálido, 4 a 6 flores masculinas por nudo, la misma cantidad de flores femeninas por nudo. Los frutos son cápsulas de 0.7 a 1.0 cm de longitud y 0.6 a 0.9 cm de ancho. Semillas con 3.5 a 4 mm de longitud y 3 a 4 mm de ancho, esferoidales a elipsoidales, con tres lóbulos, con la cubierta pardo-rojiza (Rivera-Lugo y Solano, 2012; Hernández et al., 2012).
B. gracilis es una especie endémica y amenazada, que solamente se encuentra en el Valle de Tehuacán-Cuicatlán, México (CONANP, 2013; Rivera-Lugo y Solano, 2012), donde representa un elemento prominente dentro del matorral xerófilo (Rzedowski, 1998). B. gracilis florece anualmente y las semillas maduran en la época de lluvias (Hernández et al., 2012). La especie tiene 35 inflorescencias por individuo en promedio, así como 2 600 semillas por inflorescencia. Sin embargo, la supervivencia de semillas y plántulas es muy baja, pues las plántulas son comidas por cabras o mueren por la sequía, mientras que las semillas son comidas por roedores o arrastradas por el agua de lluvia. El bajo reclutamiento, natural o antropógeno, la extracción de individuos jóvenes o adultos para el comercio, el pastoreo y la destrucción del hábitat por la construcción de caminos y la urbanización, son una amenaza para la especie (Cardel et al., 1997; Flores, 2003; Hernández et al., 2012).
Beaucarnea gracilis se emplea como planta ornamental altamente cotizada, en algunas zonas como puesto para vigilar rebaños de chivos, también en la construcción. Con sus hojas se confeccionan sombreros y adornos artesanales. Algunas especies del género son posible fuente de precursores de hormonas esteroidales (CONANP, 2013; Hernández et al., 2012). La explotación y forma de uso de las especies del género en la actualidad, con extracción de individuos jóvenes, adultos y semillas, ha llevado a la reducción de las poblaciones, y de la diversidad genética, y a la alteración de la tasa de sexos (Hernández et al., 2012).
La propagación por semilla es relevante porque contribuye a la conservación de la riqueza genética. Asimismo, Stevenson (1980) señala que las plantas de B. recurvata Lem. procedentes de semilla forman un ensanchamiento en la base del tallo, que no se observa en las plantas propagadas vegetativamente. Sin embargo, no hay información de si las semillas de la especie son ortodoxas o recalcitrantes, de su capacidad germinativa sin la remoción del fruto y aunque hay investigaciones sobre el efecto de la temperatura en la germinación, todavía queda margen para ampliar estudios con dicho factor.
Latencia y especies del desierto
La latencia es un estado en el cual la semilla no germina, aunque cuente con los niveles adecuados de factores limitativos para hacerlo. Se han establecido varios tipos de latencia (individuales o en combinación): como la física (cubierta seminal dura e impermeable); la mecánica (cubierta o endocarpo duro mas no impermeable); la química (presencia de inhibidores químicos en la semilla o el endocarpo); la fisiológica (bloqueos metabólicos) y la morfológica (embrión no completamente desarrollado), si bien la latencia química ha tendido a ser considerada como parte de la latencia fisiológica (C.Baskin y J. Baskin, 2001; C. Baskin y J. Baskin, 2004). No obstante, la latencia también es dividida en exógena (física, mecánica y química) (Mayer y Plojakoff-Mayber, 2014) y endógena (fisiológica, morfológica o combinada) (Fenner y Thompson, 2005). Los factores naturales y prácticas de vivero que terminan con tales tipos de latencia, son: latencia física (paso por tracto digestivo, fuego/escarificación), latencia mecánica (remojo y secado alternos del pireno hasta ruptura en ambos casos), latencia química (remojo en ambos casos/remoción del fruto), latencia morfológica (horas frío/estratificación a bajas temperaturas) y latencia fisiológica (horas frío/estratificación a bajas temperaturas) (C. Baskin y J. Baskin, 2001).
Las especies vegetales de los desiertos cuentan con diferentes mecanismos de latencia en sus semillas, que permiten la germinación hasta el arribo de condiciones ambientales favorables, de humedad y, en especial, temperatura. Los principales tipos de latencia que se han registrado para las especies desérticas que cuentan con tal mecanismo son: física, fisiológica y química (C. Baskin y J. Baskin, 2001). Por ejemplo, la especie del desierto de Atacama, Nolana jaffuelii I. M. Johnst., mantiene una latencia físico-fisiológica, en tanto arriban condiciones favorables, en particular disponibilidad de agua, para la germinación (Cabrera et al., 2015). La semilla de Ochradenus baccatus Delile, del desierto de Arabia, cuenta con latencia fisiológica (Bhatt y Pérez-García, 2016), en tanto que la latencia físico-química fue reportada para la especie del desierto del norte de China, Corispermum patelliforme Lijin (Liu et al., 2015).
Todo proceso fisiológico tiene temperaturas óptimas para su ocurrencia (Lambers, Chapins y Pons, 2008) y la germinación no es la excepción. Por ejemplo, en varias especies desérticas o de selvas bajas de México, se ha registrado que temperaturas entre 20 a 30 °C optimizan la germinación, como se ha encontrado en Agave cupreata Trel. & Berger (23/19 °C) (Mendoza-Celino et al., 2017) y en Chloroleucon mangense (Jacq.) Britton & Rose (30 °C), en este último caso previa escarificación química para superar la latencia física (Rodríguez-Trejo y Guizar-Nolazco, 2016).
Asimismo, cuando la producción de semillas es abundante, el almacenamiento permite su utilización en años de escasez. Si bien los primeros meses o años las semillas de varias especies pueden entrar y salir del estado de latencia, dependiendo de las condiciones de almacenamiento, en general tanto semillas ortodoxas (con bajo contenido de humedad y larga viabilidad) como semillas recalcitrantes (con alto contenido de humedad y corta viabilidad) tenderán a reducir su viabilidad al aumentar el tiempo de almacenamiento (C. Baskin y J. Baskin, 2001). En especies de zonas áridas, sin latencia física, es escasa la información sobre el efecto del tiempo de almacenamiento en viabilidad y capacidad germinativa.
El objetivo del presente estudio, para ambas especies fue: establecer el efecto de la remoción del fruto y de diferentes regímenes térmicos en la germinación. Y para B. gracilis, determinar el efecto del almacenamiento en la capacidad germinativa de la semilla.
La hipótesis es que, debido a la presencia de inhibidores en el fruto, la remoción de éste favorecerá la germinación y que una temperatura fresca favorecerá una mayor germinación. Se espera que el almacenamiento de la semilla reduzca la capacidad germinativa de B. gracilis.
Materiales y métodos
D. lucidum
La semilla de D. lucidum fue recolectada en la Reserva de la Biosfera Tehuacán-Cuicatlán, Puebla, México, en noviembre de 2014. Se trabajó con 380 g en el laboratorio. El análisis se llevó a cabo conforme a las normas de International Seed Testing Association [ISTA], señaladas por Bonner et al. (1994). Se evaluó:
Pureza (Pu), con el modelo:
Donde: Pl = peso de frutos limpios, Pi = peso de frutos con impurezas.
El peso (P), se determinó para frutos y semillas. Para ello, se pesaron 10 muestras de 100 frutos cada una, los resultados se sumaron y se hizo la extrapolación a número de frutos por kilogramo. Después se extrajo la semilla, que fue pesada, para establecer el peso de ésta de la forma ya descrita.
Para estimar la viabilidad (V), se cortaron por la mitad 100 semillas y fueron colocadas en un recipiente, sumergidas en una solución de cloruro de 2,3,5-trifeniltetrazolio al 1 % durante 24 h. Se usó el modelo:
Donde: Sv = número de semillas viables y S = número total de semillas.
El diseño experimental fue en bloques completos al azar, con los bloques anidados dentro del factor régimen de temperatura. Los tratamientos fueron régimen de temperatura y remoción del fruto. En el primer caso los niveles fueron dos: 22/19 °C y 20/17 °C, para día/noche, con un fotoperiodo de 12 h, aplicados en cámaras de ambiente controlado marca Conviron. También se tuvieron dos niveles de remoción de fruto (con y sin éste). Cada caja germinadora constituyó un bloque. Las unidades experimentales constaron de 100 semillas o frutos cada una.
Para realizar la siembra, las semillas fueron colocadas sobre tela fieltro en las cajas, a su vez sobre rejillas de plástico, que fueron humedecidas previamente con 100 ml de agua para que se mantuvieran constantemente húmedas; la mayor parte del líquido quedó depositado bajo la rejilla, con la tela en contacto con el líquido para garantizar que se mantuviera húmeda. Se aplicó al agua el fungicida Captan (3 g L-1) como precaución ante posible afectación por hongos.
Se consideró germinada cualquier semilla cuya radícula alcanzaba la misma longitud que la semilla. La germinación se registró diariamente. Se evaluó la energía germinativa como el número de días para alcanzar 75 % de la capacidad germinativa. Para realizar los análisis estadísticos, los valores de capacidad germinativa (G), en por ciento, fueron transformados (Gt) mediante la función arco seno:
El modelo estadístico empleado, fue:
Donde: yijkl = observación, µ = media general, αi = efecto del i-ésimo nivel del factor régimen de temperatura, βj = efecto del j-ésimo nivel del factor remoción del fruto, αi (γ)k = efecto del k-ésimo bloque anidado dentro del i-ésimo nivel del factor régimen de temperatura, αiβj = el efecto de la interacción entre el i-ésimo nivel del factor régimen de temperatura y el j-ésimo nivel del factor remoción del fruto, αiβj(γ)k =efecto de la interacción entre el i-ésimo nivel del factor régimen de temperatura, el j-ésimo nivel del factor remoción del fruto y el k-ésimo bloque anidado dentro del factor régimen de temperatura, εijkl = error experimental.
Se realizó análisis de varianza mediante un procedimiento mixto, con el procedimiento PROC MIXED, del programa SAS para microcomputadoras, versión 9. Se consideró con efectos fijos a los factores temperatura y remoción del fruto, y su interacción. Los efectos de los bloques y todas las interacciones en las cuales participan se tomaron como efectos variables.
B. gracilis
Se recolectaron frutos de esta especie de dos poblaciones distintas en diferentes épocas del año, en la Reserva de la Biosfera de Tehuacán-Cuicatlán, la primera en diciembre de 2016 (0.25 kg) y la segunda en abril de 2017 (0.25 kg). Para diferenciar estas semillas se les designó con los nombres de semilla vieja y semilla joven, respectivamente. Fueron calculadas: pureza, peso, energía germinativa y viabilidad. Estas pruebas se llevaron a cabo con el mismo modelo que se describió para D. lucidum.
Se realizaron dos experimentos: el primero con los factores (y niveles) de remoción del fruto (con y sin remoción) y temperatura (30 °C y 25/20 °C); el segundo con los factores (y niveles) edad de la semilla (joven y vieja) y temperatura (30 °C y 30/25 °C). El primer experimento se realizó en febrero de 2017 con la muestra recolectada en 2016, y el segundo en junio del mismo año, con muestras recolectadas en 2016 y 2017. En el primer experimento, para cada nivel de temperatura y cada nivel de remoción del fruto, se establecieron cuatro repeticiones de 50 semillas cada una. Se sembraron 100 semillas en cada caja (la caja correspondió al bloque), la cual contenía semillas con y sin fruto, con una disposición al azar. Cuatro cajas fueron colocadas en una cámara de ambiente controlado a 30 °C y otras cuatro cajas fueron colocadas en otra cámara a 25/20 °C. En ambos casos el fotoperiodo fue de 12 h.
Para el segundo experimento se utilizaron 25 semillas jóvenes y 25 semillas viejas, por repetición, colocadas al azar dentro de las cajas de siembra. También se manejaron cuatro repeticiones (bloques), por cámara con las temperaturas referidas en el párrafo anterior. El diseño experimental para ambos experimentos fue en bloques completos al azar, con dos tratamientos, cada uno, con dos niveles y con los bloques anidados en el factor temperatura.
Se realizó un análisis de varianza con un procedimiento mixto, utilizando el procedimiento PROC MIXED, del programa SAS para microcomputadoras, versión 9. Los bloques y sus interacciones se consideraron con efectos aleatorios y los tratamientos con efectos fijos. Para el análisis se hizo la transformación de la variable germinación con la función arco seno (fórmula 3). También se llevó a cabo una comparación de medias mediante la prueba de Tukey.
El modelo estadístico usado fue:
Para el experimento 1:
Donde: 𝑦𝑖𝑗𝑘𝑙= observación, 𝜇 = media general, 𝛼𝑖 = efecto del i-ésimo nivel del factor temperatura, ( ) = efecto del j-ésimo bloque anidado dentro del i-ésimo nivel del factor temperatura, 𝛾𝑘 = efecto del k-ésimo nivel del factor remoción de fruto. Las combinaciones de letras denotan interacciones entre los factores que representan, 𝜀𝑖𝑗𝑘𝑙 = error experimental.
Para el experimento 2:
Donde: 𝑦𝑖𝑗𝑘𝑙 = observación, 𝜇 = media general, 𝛼𝑖 = efecto del i-ésimo nivel del factor temperatura, 𝛼𝑖 ( )= efecto del j-ésimo bloque anidado dentro del i-ésimo nivel del factor temperatura, 𝛾𝑘 = efecto del k-ésimo nivel del factor edad de la semilla. Las combinaciones de letras denotan interacciones entre los factores que representan, 𝜀𝑖𝑗𝑘𝑙 = error experimental.
Resultados y discusión
D. lucidum
Análisis de la semilla
Se halló una pureza para frutos de 94.1 %. La especie tiene 91 798 frutos limpios kg-1 y 141 614.9 semillas kg-1, es decir, 1000 semillas pesan 7.08 g. La viabilidad fue de 94 %. El fruto y semilla se muestran en la Figura 2. Cabe señalar que Missouri Botanical Garden (2019) refiere que las semillas de la familia Asparagaceae, subfamilia Nolinoideae, a la que pertenece el género Dasylirion, cuentan con endospermo como tejido de reserva.
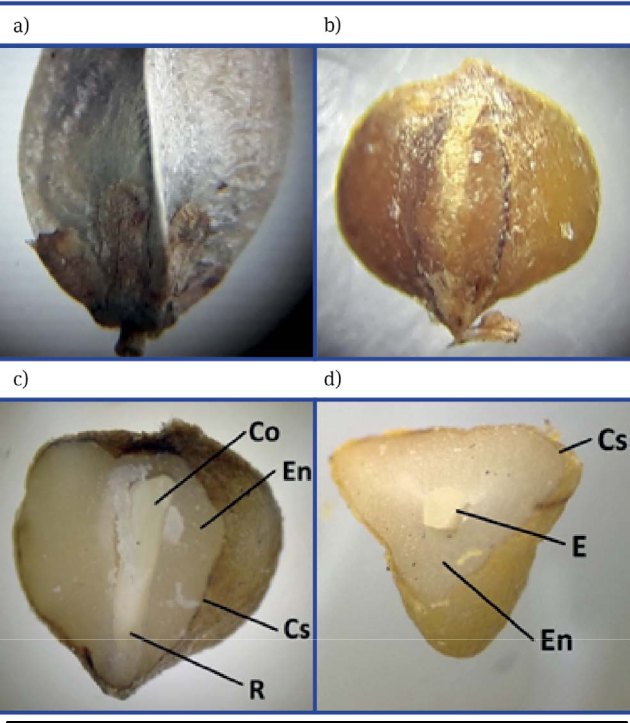
Germinación
El máximo de germinación se alcanzó entre 14 y 19 días para los diferentes tratamientos. El tratamiento a temperatura baja, sin fruto, fue el que alcanzó el máximo en menos tiempo, 14 días (Figura 3), aunque con baja capacidad germinativa.
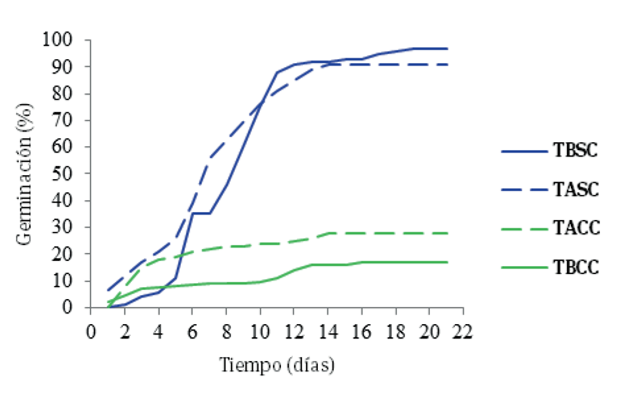
Nota: TBSC = temperatura baja sin fruto, TASC = temperatura alta sin fruto, TACC = temperatura alta con fruto, TBCC = temperatura baja con fruto. Fuente: elaboración propia.
Figura 3 Germinación acumulada de D. lucidum
Para la capacidad germinativa, resultaron significativos el factor remoción de fruto (p ≤ .0001), así como la interacción remoción de fruto y régimen de temperatura (p ≤ .0001). La germinación de semillas con el fruto, fue igual a 22.3 %, mientras que la de la semilla removida del fruto alcanzó 94.3 %. Respecto a la energía germinativa, fue de 7 días para la semilla con fruto, y de 10 días para la semilla sin fruto. La capacidad germinativa es similar a la referida por Sierra-Tristán y Morales (2002), para D. leiophyllum Engelm. ex Trel. como se aprecia en la Figura 4, donde se representa la interacción entre los factores estudiados, la semilla que fue removida del fruto, alcanzó una mayor germinación en ambos regímenes térmicos, pero el mayor valor se alcanzó en el régimen térmico bajo. Contrasta que la semilla con fruto, germinó mejor en el régimen de temperatura alto.

Fuente: elaboración propia.
Figura 4 Interacción entre régimen de temperatura y remoción de fruto, para la capacidad germinativa de D. lucidum
Hay investigaciones con otras especies de Dasylirion, en las cuales la remoción de las brácteas (fruto) promueve la germinación. En D. leiophyllum, Sierra-Tristán y Morales (2002), refieren que la capacidad germinativa sin remover la bráctea tuvo un promedio de 3 %, mientras que removiéndola alcanzó 95.8 %. Con los mismos tratamientos, D. sereke Bogler tuvo <2 % y 80 % de capacidad germinativa, respectivamente (Vega-Cruz, Melgoza-Castillo y Sierra-Tristán, 2006).
El aumento en la germinación de la semilla de Dasylirion, gracias a la remoción de las brácteas, deja ver la presencia de latencia. Sin embargo, al parecer no se ha logrado concluir de qué tipo.
Las brácteas retrasan la germinación por algunos meses y, dado que las semillas maduran hacia fines de verano u otoño, esto puede favorecer que la germinación de las semillas coincida con la siguiente temporada de lluvias (Vega-Cruz et al., 2006; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias [INIFAP] y Centro de Investigación Regional Norte Centro [CIRNOC], 2008 y Reyes-Valdés et al., 2012). El que la semilla de las especies de Dasylirion mejore significativamente su germinación solo si se remueven las brácteas, indica que potencialmente podría haber los siguientes tipos de latencia: física y química. Sin embargo, las brácteas no son muy duras ni impermeables, se remueven fácilmente con las manos. Si bien es posible una latencia física leve y temporal, conferida por las brácteas, es más probable la presencia de químicos inhibidores de la germinación (latencia química) en éstas, que con las primeras lluvias se lavan y ello facilita, junto con la mayor humedad disponible, la germinación y establecimiento de la plántula. Esta estrategia existe en especies del desierto.
El mecanismo específico de latencia puede relacionarse con la presencia de productos como el ácido abscísico, inhibidor de la germinación, que se ha registrado por Slabbert, Motsa y Averbeke (2015) en los embriones de Brassica rapa ssp. chinensis y en S. retroflexum. Estas especies mejoraron su germinación, respectivamente, de 74 a 95 % y de 46 a 95 %, luego de ser remojadas en KNO3 y aplicarles giberelinas, lo cual se relaciona con la reducción de los niveles de ácido abscísico en los embriones y con el efecto exógeno de las giberelinas. La especie australiana Persoonia longifolia R. Br. requiere de remojo en agua para solubilizar los inhibidores químicos, pero también de ácido giberélico para promover su germinación (Mullins, Koch y Ward, 2002). En China, Corispermum patelliforme tiene latencia física y química. Se debe remover la cubierta seminal o escarificarla, además de remojar la semilla en ácido giberélico, para lograr una buena germinación (Liu et al., 2015). La especie naturalizada en México el pirúl, Schinus molle L., también presenta latencia química por la presencia de inhibidores en la cubierta seminal, que son solubilizados mediante remojo en agua (Camacho, 2011). Este último autor, refiere que entre las sustancias que inhiben la germinación en la latencia química, se ha hallado al ácido acetilsalicílico y a la cafeína, entre otras.
Aunque con ligera diferencia (6 %), la mejor germinación de la semilla sin fruto a temperatura baja, puede relacionarse con mayor cercanía a las temperaturas día/noche óptimas para la germinación; en tanto que la tendencia es contraria, la diferencia de 11 % y con menor germinación en presencia del fruto. En este último caso, el agua utilizada para activar la germinación pudo lixiviar parte de los inhibidores del delgado fruto. Diversas especies de zonas semiáridas muestran mejor germinación a temperaturas cercanas al régimen 20/17 °C utilizado, ya que de esa manera tienden a conservar más la humedad necesaria para germinar y se mantienen cerca del intervalo de temperaturas óptimas para la germinación. Astragalus arpilobus Kar. & Kir., del desierto central de Asia, cuya germinación fue probada por Long et al. (2012) en regímenes térmicos de entre 5/2 y 30/15 °C, tuvo las mayores germinaciones a 20/10 y 25/15 °C. Tendencia similar a la de Nolina parviflora (H.B.K.) Hemsl., de zonas semiáridas del centro de México, cuya germinación alcanzó 87.5 y 88.1 % a 20 y 25 °C, respectivamente (en ambos casos sin luz), mientras que a 30 °C declinó a 15.8 %. Con luz la germinación declinó a mayores temperaturas (85 % a 20 °C, 42.3 % a 25 °C y 0.6 % a 30 °C (Reyes-Bautista y Rodríguez, 2005).
Sin embargo, al parecer no todas las especies de Dasylirion tienen latencia, pues Francisco-Francisco et al. (2012) y Reyes-Valdés et al. (2012), hallaron que algunos sotoles, como D. cedrosanum Trel., germinan bien (>90 a 98 %) incluso sin tratamientos pré-germinativos, siempre que haya buena iluminación y disponibilidad de humedad en el sustrato. No obstante, el agua de riego podría haber contribuido a la solubilización de los inhibidores de haber latencia química moderada en esa especie.
La aplicación de ácidos fúlvicos aumenta la velocidad de emergencia (Cruz, 2011), y el extracto de raíz de Agave lechuguilla Torr. (0.05 mg), favoreció una germinación de 96.8 % en D. cedrosanum. Esto puede deberse a la acción de los grupos funcionales de las substancias orgánicas, que incrementan actividad enzimática, lo cual favorece el desdoblamiento de sustancias de reserva y la multiplicación celular (Reyes-Valdés et al., 2012).
La capacidad germinativa y la velocidad de germinación, varían entre localidades, como hallaron Sierra-Tristán y Morales (2002) para D. leiophyllum. La gran producción de semillas por individuo, así como su elevada capacidad germinativa, posiblemente aseguran el establecimiento inicial de numerosas plántulas (Reyes-Valdés et al., 2012), si bien bajo condiciones naturales, diversos factores limitativos como sequía y forrajeo por fauna o ganado la limitan (Vega-Cruz et al., 2006).
B. gracilis
Análisis de la semilla
De B. gracilis, se obtuvieron 33 251.1 frutos limpios kg-1; 33 745.4 frutos kg-1 de frutos con impurezas, así como 46 152.2 semillas kg-1. Fruto y semilla se muestran en la Figura 5. Missouri Botanical Garden (2019) señala que las semillas de la familia Asparagaceae, subfamilia Nolinoideae, a la que pertenece el género Beaucarnea, tienen endospermo como tejido de reserva.
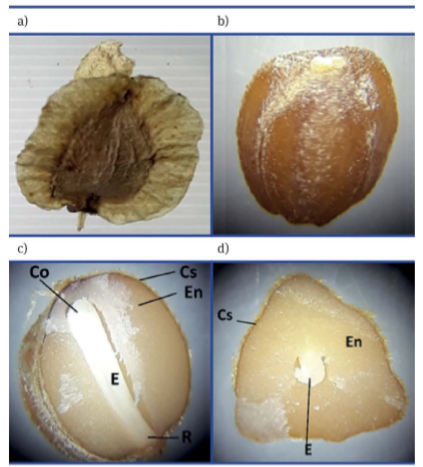
Primer experimento
La remoción del fruto (p = .0323) fue significativa en este experimento, no así la temperatura (p = .1887) ni la interacción de ambos factores (p = .1887). La germinación sin remoción del fruto fue nula, pero con su remoción fue igual a 20.5 %. Este lote mostró una germinación menor que la del empleado en el segundo experimento. Es bien conocida la variabilidad en la capacidad germinativa entre poblaciones y entre procedencias, como hallaron Sierra-Tristán y Morales (2002) para D. leiophyllum.
Segundo experimento
Para la variable capacidad germinativa, ni temperatura (p = .9772), ni edad de la semilla (p = .8471), ni su interacción (p = .3838) resultaron significativos (Cuadro 1). La capacidad germinativa promedio alcanzó 89.3 % y la energía germinativa media fue de 15 días. La Figura 6 muestra la curva de germinación acumulada.
Cuadro 1 Comparación de medias de Tukey, para la germinación de B. gracilis
| Factor | Media (%) |
|---|---|
| Temperatura | |
| 30 °C | 90.5a |
| 25/20 °C | 88.0a |
| Edad | |
| Semilla joven | 88.5a |
| Semilla vieja | 90.0a |
Nota: Letras iguales en los niveles de cada factor, denotan que no hay diferencias significativas.
Fuente: elaboración propia.

La semilla de la especie estudiada se considera no latente en algunas fuentes (Cardel et al., 1997; C. Baskin y J. Baskin, 2001). Sin embargo, los resultados del presente trabajo y los resultados con otras especies de Beaucarnea, sugieren lo contrario. Jardim-Rosa et al. (2014), hallaron para B. recurvata Lem., que la remoción del pericarpio produjo una germinación de 60 %, diferente estadísticamente del 37 % cuando no fue removido. Lo mismo sucedió para el índice de velocidad de emergencia, cuyos valores fueron, respectivamente de 0.95 y 0.61. El lavado de inhibidores debe ocurrir de manera semejante a como se discutió para D. lucidum.
Las semillas responden bien a la escarificación química con ácido sulfúrico; al aumentar la concentración de éste de 40 hasta 90 %, la germinación aumenta de 20 a 100 %. Semillas sumergidas en agua durante 24 h también alcanzaron 100 % de germinación (Cardel et al., 1997). Aunque no se especifica el tipo de latencia, los resultados de este tratamiento se pueden usar para confirmar la presencia de latencia, física leve y química. La fácil remoción manual (o por desgaste en condiciones naturales), o por el ácido, dejan ver la posibilidad de existencia de latencia física leve. No obstante, es más probable la presencia de latencia química, con sustancias inhibidoras de la germinación en las brácteas, que, al ser removidas con las manos, con el ácido o solubilizadas con el agua de lluvia, se elimina el bloqueo a la germinación. Cabe recordar que la germinación en época de lluvias es una estrategia adaptativa común en plantas del desierto, como también se mencionó para D. lucidum.
Flores y Briones (2001) registraron 95 % de germinación en cámaras de ambiente controlado a 26 °C, contra 2 % a 20 °C y 0 % a 12 °C. En las dos pruebas del presente estudio no se halló influencia de la temperatura sobre la germinación, lo que obedece a que los regímenes térmicos usados por dichos autores son más variados.
Comparando dos especies arbustivas [(Cercidium praecox (Ruiz & Pavón), Prosopis laevigata (Humb. & Bonpl. ex Willd.)] con dos especies columnares suculentas [(Neobuxbaumia tetetzo (F.A.C. Weber) Backeb var. tetetzo, Pachycereus hillianus (F.A.C. Weber) F. Buxb.] y otra especie arborescente semisuculenta, Yucca periculosa F. Backer, se encontró que el potencial hídrico igual a -0.41 MPa, no representó un umbral por debajo del cual (a -0.61 MPa) se redujera la germinación en P. laevigata ni P. hollianus, pero sí para el resto de especies, incluida B. gracilis. Es decir, no es una de las especies que pueda germinar con una elevada tensión hídrica, como otras sí lo pueden hacer. Por formas de vida, se tuvieron diferencias significativas entre ellas, con la siguiente secuencia en la germinación: arbustos (82.5 %), arborescentes semisuculentas (52 %) y columnares suculentas (42.6 %) (Flores y Briones, 2001).
Si bien en las semillas recalcitrantes la capacidad germinativa declina más rápido que en las ortodoxas, eventualmente hay una reducción en tal capacidad conforme la semilla de cualquier especie permanece almacenada por más tiempo (C. Baskin y J. Baskin, 2001). No obstante, el tiempo de almacenamiento del presente estudio, seis meses, resultó breve como para arrojar diferencias con respecto a la semilla recolectada recientemente.
Conclusiones
Hay evidencia de latencia química tanto en D. lucidum como en B. gracilis, que en el ambiente natural cesa con las primeras lluvias y en laboratorio con la remoción de las brácteas que constituyen el fruto alado. Sin embargo, todavía debe probarse que el lavado de estos productos promueve la germinación, así como la identificación de los agentes químicos que provocan dicha latencia.
La remoción de la semilla del fruto es recomendable, a efecto de incrementar significativamente la capacidad germinativa de las especies estudiadas.
El régimen día/noche 20/17 °C y la remoción del fruto, maximizaron la germinación de D. lucidum. En B. gracilis, ni la temperatura ni la edad probadas influyeron en la capacidad germinativa. No obstante, es necesario probar edades más viejas de la semilla, para establecer su longevidad, además de incluir almacenamiento en diferentes condiciones, por ejemplo, a temperaturas cercanas al punto de congelación.
Las diferencias en la germinación entre las dos pruebas realizadas para B. gracilis, dejan ver la existencia de diferencias de germinación entre poblaciones.
El almacenamiento de la semilla de B. gracilis, en condiciones de cuarto fresco, mantiene la capacidad germinativa de la especie durante seis meses por lo menos.

