











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En Tamaulipas, la producción pesquera anual promedio es de más de 52 000 T, con un valor superior a los 930 millones de pesos aportando el 8.86 % del Producto Interno Bruto, dentro del sector agropecuario (Rodríguez-Castro y col., 2010). En orden decreciente, las especies que más contribuyen a la producción pesquera de Tamaulipas son: camarón, Farfantepenaeus aztecus, Ives, 189i (13 404 T); lisa, Mugil cephalus, Linnaeus, 1758 (3 990 T); mojarra-tilapia, Oreochromis aureus, Steindachner, 1864 (3 235 T); jaiba azul, Callinectes sapidus, Rathbun, 1896 (2 733 T); ostión, Crassostrea virginica, Gmelin, 1791 (2 303 T); carpa, Cyprinus carpio, Linnaeus, 1758 (2 261 T) y el grupo denominado tiburón cazón que abarca principalmente a las especies: Rhizoprionodon terraenovae, Carcharhinus acronotus, Sphyrna lewini, Carcharhinus leucas, Carcharhinus falcifor-mis y Carcharhinus porosus (2 033 T).
Actualmente, la pesquería de la jaiba azul (JA) de la Laguna Madre, Tamaulipas (LMT), al igual que en el resto del Golfo de México, se encuentra aprovechada al máximo sustentable (Arreguín-Sánchez y Arcos-Buitrón, 2011; SAGARPA, 2012). En dicha actividad económica participan más de 3 500 pescadores, y dependen de ellos más de 15 000 personas ubicadas en los municipios de Matamoros, San Fernando y Soto La Marina. Esta situación de sobreexplotación reviste crucial importancia en la LMT, en donde se realiza el 82 % de la captura estatal de esta pesquería (SAGARPA, 2012).
Arreguín-Sánchez y Arcos-Huitrón (2011), indicaron que: "en muchos países, un problema común es la insuficiencia de información clave y continua para la aplicación de modelos que permitan realizar un diagnóstico confiable del estado de la pesca, y representar escenarios de manejo con niveles de incertidumbre y riesgo calculables. Por lo general, el único dato que parece ser relativamente consistente son las capturas registradas. Aún con las deficiencias que pudieran señalarse para los registros de captura de las flotas ribereñas, la tendencia de las capturas, especialmente cuando se trata de recursos plenamente explotados, suele ser un indicador relativo de la abundancia del recurso; y la tendencia puede ofrecer información sobre el estado de la pesca; ya que en muchos casos, es la única información disponible". Por ello, en la evaluación de los recursos pesqueros, partiendo de la información de series de tiempo de captura, como única fuente de información, es importante analizar su tendencia.
En el análisis de la tendencia de las series de tiempo se incluye, en diferentes métodos, la utilización de funciones matemáticas para arribar a proyecciones confiables. En tal situación se ubican los modelos de regresión lineal y no lineal (o curvilínea). Bajo el marco de la selección de modelos, hay una gama de modelos que se ajustan a un grupo de datos en particular, tal es caso de las series temporales, por lo que es necesario establecer un criterio de decisión para elegir el mejor modelo, como es el de información (Burnham y Anderson, 2004). La inferencia multimodelo (IMM) postula que cuando los datos soportan evidencia para más de un modelo, en lugar de estimar los parámetros a partir de sólo "el mejor modelo", es posible estimarlos a partir de varios modelos, e incluso todos los modelos considerados. Obtener el valor promedio de la variable predicha, a partir de varios modelos, es una ventaja para alcanzar una inferencia robusta, que no esté condicionada a un sólo modelo y que integre la aportación ponderada de cada modelo ajustado (Burnham y Anderson, 2002).
Con base en las series de tiempo de la captura pesquera, se han desarrollado modelos generales de clasificación de las fases de desarrollo de una pesquería (Caddy, 1984; Csirke, 1984), con poca variación en las descripciones de las fases de crecimiento, y que definen la vida por etapas de una pesquería. La captura (C) es una cantidad considerada como índice, que es proporcional a parámetros importantes de la pesquería, como la mortalidad por pesca y la densidad de la proporción explotada (Gulland, 1971).
En el plano de la evaluación y manejo de las pesquerías, en primera instancia, se tiene que revisar la pesquería a través de la tendencia de la C, ya que determinará la fase de desarrollo en que se encuentra, y según la fase de desarrollo actual, establece la definición de técnicas y objetivos de la ordenación pesquera que se deberán de aplicar. Csirke (1984), señala que: "por lo general, todo cambio en gran escala en un recurso explotado determinará una reacción equivalente en la pesquería. Si las poblaciones de peces aumentan, lo harán también las tasas de captura, la captura total y el esfuerzo de pesca. En cambio, si la población disminuye, las capturas también lo harán, pero el esfuerzo de pesca se reducirá más lentamente y es posible incluso que se aumente, en el intento de mantener altos niveles de captura, con lo cual la mortalidad por pesca real aumentará. Esta forma de comportamiento hace que las pesquerías que son afectadas por importantes fluctuaciones, debidas al ambiente, tengan mayores posibilidades de llegar a una situación de sobreexplotación".
La presente investigación tuvo como objetivo modelar la captura anual de la Jaiba azul en la Laguna Madre, Tamaulipas, mediante el ajuste de funciones matemáticas de tipo lineal y no lineal (o curvilínea) a la serie de tiempo, de 1998 a 2012, e identificar las fases de desarrollo de la pesquería, de acuerdo a los modelos analizados.
MATERIALES Y MÉTODOS
Recopilación de información
La información que se utilizó para este estudio fue obtenida de la Subsecretaría de Pesca y Acuacultura de la Secretaría de Desarrollo Rural del Gobierno del Estado de Tamaulipas. El origen de esta información son los avisos de arribo para embarcaciones menores de las oficinas de pesca de los municipios de Matamoros, San Fernando y Soto La Marina, dependientes de la Subdelegación de Pesca de la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA), en Tamaulipas. Los avisos de arribo son formatos oficiales generados por la SAGARPA, a través de la Comisión Nacional de Pesca y Acuacultura, para que todos los permisionarios reporten sus capturas de forma mensual. Las especificaciones de dichos formatos incluyen, entre otras, el nombre de la especie, el volumen y el valor de la captura en kilogramos por mes, principalmente.
Ajuste de modelos para el análisis de captura (C)
En una hoja de cálculo del software Numbers versión 3.5.3 (2015), se construyó la base de datos, las gráficas correspondientes y se estimaron los estadísticos básicos (totales y promedios). Se realizaron ajustes de modelos lineales y no lineales (estimación curvilínea) a la serie de tiempo de la C de la JA en la LMT durante el periodo 1998 a 2012, utilizando el software Statistical Package for the Social Sciences (SPSS, Version 18, 2009). Los modelos aplicados se indican en la Tabla 1. De cada modelo se obtuvo el coeficiente de determinación (R2), la prueba F de Fisher, la probabilidad de significancia; las estimaciones de los interceptos (β 0) y coeficientes de regresión (β 1, β 2, β 3), y las probabilidades asociadas a la prueba t de estos parámetros en cada modelo de regresión.
Tabla 1 Modelos lineales y no lineales aplicados a la serie de tiempo de 1998 a 2012 de la captura total anual de la jaiba azul Callinectes sapidus en la Laguna Madre, Tamaulipas, México.
Table 1. Linear and nonlinear models applieds to the time series of 1998 to 2012 of the total annual catch of blue crab (Callinectes sapidus) in Laguna Madre, Tamaulipas, Mexico.

y = variable dependiente; x = variable independiente; ß0 = constante (intercepto); ßl, ß2 y ß3 = coeficientes de regresión; Y = variable dependiente; b0, b1 y b2 = coeficientes de regresión; t = variable tiempo; ln = logaritmo natural; e = función exponencial; u = valor del límite superior.
Selección de modelos
La selección de los modelos se basó en la teoría de la información, específicamente en el Criterio de Información Akaike (1973) (CIA) y Criterio de Información Bayesiano (CIB), propuesto por Schwarz (1978). De cada modelo se obtuvieron los valores CIAc (CIAc = CIA corregido, más adelante se indican las razones de esta corrección) y CIB, así como las diferencias (∆i) y el peso (wi) para CIAc y CIB respectivamente. Con estos resultados se procedió a encontrar los valores más bajos CIAc y CIB, y los más altos de los wi CIAc y wi CIB, para identificar el mejor o mejores modelos, del total de los modelos candidatos. Para complementar estos criterios de selección, también se utilizó la significancia estadística del β 0, β 1, β 2, β 3 , de cada uno de los modelos candidatos, con base en la probabilidad asociada a la prueba t.
Criterio de Información Akaike (CIA)
Este criterio se basa en la distancia Kull-back-Leibler, que mide la aproximación del modelo calculado con los datos reales, seleccionando así el mejor modelo candidato (Katsanevakis, 2006; Katsanevakis y Maravelias, 2008). El enfoque CIAc permite jerarquizar los modelos, al ordenarlos de forma ascendente, en función de los valores CIAc. El modelo con el menor valor CIAc será el mejor (Cailliet y col., 2006; Romine y col., 2006; Griffiths y col., 2010). Guzmán-Castellanos y col. (2014) establecieron que: "la premisa más importante del método del CIAc es penalizar la cantidad de parámetros de cada uno de los modelos, basándose en el principio de parsimonia; es decir, existe un criterio basado en la bondad de ajuste del modelo a los datos, definido a través de la función objetivo de máxima verosimilitud o SSQ (suma de cuadrados), y al mismo tiempo existe una penalización asociada a la cantidad total de parámetros del modelo. En consecuencia, se esperaría un mejor ajuste del modelo a los datos, en la medida que aumenta el número de parámetros de los modelos. El CIAc equilibra adecuadamente estos dos componentes, y su resultado finales un criterio cuantitativo para la selección de modelos".
Formulación CIA
La expresión original del CIA es la siguiente:
Donde -lnL es el logaritmo natural negativo de verosimilitud estimado por una función logarítmica, y θ es el número de parámetros estimados en cada modelo candidato de crecimiento.
Sin embargo, en este estudio se utilizó el enfoque de los mínimos cuadrados, y el parámetro del logaritmo natural negativo de verosimilitud fue sustituido por la varianza residual de acuerdo al criterio de Burnham y Anderson (2002), quienes proponen la versión siguiente:
Donde n = tamaño muestral (número de datos), 2 = varianza residual (cociente de la suma de cuadrados residuales y el número de datos), ô 2 y K = número de parámetros del modelo.
Para esta versión se asumió una distribución normal del error, una media cero y una varianza constante.
CIA corregido (CIAc)
De acuerdo al criterio de Burnham y Anderson (2002), si el tamaño de la muestra es pequeña, es decir, si n/K < 40, que es el presente caso, entonces se debe usar una forma de estimación de CIA que pueda corregir el sesgo, y la expresión se escribe como:
Donde CIAc = CIA corregido, CIA = valor de CIA, K = número de parámetros y n = número de datos.
Delta (∆i) CIAc
Se siguió el método descrito por Guzmán-Castellanos y col. (2014), quienes establecen que: "para conocer el soporte estadístico que tienen los modelos candidatos, que no fueron considerados como el modelo más adecuado, se estiman las diferencias de Akaike definidas como ∆i; las cuales de acuerdo con Burnham y Anderson (2002), si ∆i > 10 denota modelos candidatos sin apoyo estadístico que no deben ser tomados en cuenta. Por el contrario, si ∆i < 2 entonces tienen una alta evidencia como funciones alternativas; mientras que modelos en los que 4 < ∆i <7 pueden ser tomados en cuenta, aunque con menor soporte estadístico que los anteriores. De tal forma que, ∆i es un indicador del desempeño de cada modelo candidato. La formulación para estimar ∆i se representa como: ∆i = CIAci - CIAcmin; donde: CIAcmin es el modelo candidato con el valor más bajo de CIA que representa el modelo candidato más adecuado, y CIAci representa el valor de CIA estimado para los otros modelos candidatos (i = 1...n)".
Pesos CIA (wi)
Adicionalmente, para cuantificar la evidencia a favor que tiene cada modelo candidato, también se estiman los factores ponderados de Akaike (wi) "pesos Akaike"; cuya expresión Akaike:
donde ∆i = diferencia Akaike y K = número de parámetros.
K (Cantidad de parámetros)
La varianza residual ô 2 se incluyó como un parámetro más de cada modelo, dado que se utilizó la función de mínimos cuadrados, recomendada por Burnham y Anderson (2002).
Criterio de Información Bayesiano (CIB)
También se aplicó el CIB para la selección del modelo de mejor ajuste, bajo la fórmula siguiente:
Donde L = estimador de máxima verosimilitud, K = número de parámetros (incluyendo la varianza residual como un parámetro más del modelo) y n = número de datos.
De igual forma, el estimador de máxima verosimilitud fue sustituida por la varianza residual. Una vez estimados los valores CIB, se procedió a estimar el valor de la diferencia ∆i y el valor de wi para cada uno de los modelos, de acuerdo al procedimiento indicado para los correspondientes del CIA, según lo proponen Wagenmakers y Farrel (2004). También se consideró a ô 2 como un parámetro del modelo en aplicación. Los pesos CIB pueden ser interpretados como probabilidades a posteriori del modelo, dado los datos, el conjunto de modelos y la probabilidad a posteriori (1/k) de cada modelo, siendo k el número de modelos candidatos.
Criterio de IMM
Se utilizó el criterio IMM, dado que varios modelos presentaron ajustes adecuados de predictibilidad. Este criterio consiste en rehacer la estimación completa de los valores de CIAc y/o CIB y de interceptos y coeficientes de regresión, para el subgrupo de modelos que cumplieron con el criterio de ∆i < 2 de plausibilidad. Este carácter de plausibilidad significa que dichos modelos disponen de soporte estadístico similar, es decir, que poseen un grado de explicación aceptable, en términos del ajuste de los modelos de los datos. En consecuencia, los parámetros de interés (β 0, β 1 y β 2) de los modelos bajo este criterio (∆i < 2) pueden ser y fueron mejorados. Para lograr esto, se estimó el valor esperado del parámetro de interés (Ôi), usando el siguiente estimador:
De acuerdo a Guzmán-Castellanos y col. (2014) "este estimador asume que el valor del parámetro Ōi corresponde a un valor esperado, que cruza sobre todos los modelos, donde el valor Ôi debe ser estimado, de manera individual, para cada modelo candidato que resulta con valores satisfactorios ∆i < 2. Por consiguiente, este procedimiento explica que el valor de Ôi es mejor que aquel estimado para Ōi, sin importar si el valor de Ōi correspondió al modelo candidato con el mejor valor de CIAcmin o CIB. Esto implica una corrección al valor de Ōi que mejora el valor del parámetro". Una vez estimados Ôi se estimaron sus intervalos de confianza al 95 %, con base en lo siguiente:
Donde la variable Ōi/g1 corresponde a la varianza estimada de Ōi, de acuerdo con el ajuste del modelo candidato gi. De esta manera, además de estimar el mejor valor del parámetro de interés (β 0 , β 1 y β 2 ), definido como Ōi. También se estimaron sus interva los de confianza al 95 %".
Indice de Sustentabilidad Pesquera (ISP)
Se identificaron y analizaron los periodos de expansión y contracción de la captura de la JA en la LMT durante el periodo 1998 a 2012, mediante el ISP (Ponce y col., 2006), a través de la ecuación siguiente: ISP = ln (CXiCXmean-1) Donde: CXi = captura en el año i, CXmean = captura promedio del total del periodo.
Fases de la pesquería
Se utilizó la propuesta de Csirke (1984), que refiera a la evolución usual de una pesquería a lo largo del tiempo, donde se describen las fases siguientes: (1) predesarrollo, (2) crecimiento, (3) explotación plena, (4) sobreexplotación, en algunos casos (5) colapso y, cuando fue viable (6) recuperación. Asimismo, representa dos tipos de evolución típica de algunas pesquerías neríticas: un aumento rapidísimo del esfuerzo de pesca, hasta alcanzar niveles excesivamente altos, y un aumento moderado, que llega también a niveles altos, pero a ritmo más lento. También se aplicó el modelo generalizado de una pesquería propuesto por Caddy (1984), en la cual, en el curso de su desarrollo, debe pasar a través de cuatro fases: (I) sin desarrollo, (II) de desarrollo, (III) de madurez, y (IV) de decadencia. En este modelo van implícitos los conceptos de la capacidad de carga y el esfuerzo pesquero, que corresponde este último a la tasa de extracción.
La fase sin desarrollo se caracteriza porque los recursos son explotados de forma limitada, por debajo de su potencial, con técnicas de pesca poco eficientes y muchas veces artesanales. Es importante considerar que, de acuerdo con Grainger y García (1996): "la fase de desarrollo o de crecimiento se caracteriza por un ritmo rápido de aumento de las capturas, concomitante con un crecimiento del número de embarcaciones y pescadores y con el perfeccionamiento y modernización de los sistemas de pesca. Con las ganancias que se obtienen, al iniciarse esta fase, se estimula el desarrollo de nuevas inversiones y un incremento aún mayor del esfuerzo de pesca. La fase de madurez se caracteriza porque todavía se puede apreciar un aumento de las capturas; el aproximarse, en la mayoría de los casos, a la captura máxima sostenible, determina que, aunque las capturas totales pueden aumentar, la tasa de incremento de las capturas disminuye rápidamente. Sin embargo, por la propia inercia del proceso inversionista, resulta difícil reducir el esfuerzo pesquero de manera inmediata y, por consiguiente, puede pasarse rápidamente a la fase de decadencia, que conlleva una disminución de las capturas debido a la sobrepesca, a veces exagerada por cambios ambientales desfavorables. La duración de las diferentes fases y su nivel de inclinación, dependerán tanto de la tasa del incremento de las capturas, como de la capacidad de carga del ecosistema, para cada una de las especies".
RESULTADOS Y DISCUSIÓN
Tendencia de la captura (C)
Además de la regresión lineal, en este estudio también se utilizó la estimación curvilínea (regresiones no lineales), bajo el supuesto de que los factores que han ocasionado patrones o tendencias en el pasado y en el presente, reflejados en una serie de tiempo, continuarán haciéndolo, más o menos, de la misma forma en el futuro (Bello y Martínez, 2007). En la Tabla 2 se muestran las estimaciones de R2, prueba F, gl1, gl2 (gl = grados de libertad) y de la probabilidad de significancia de las regresiones; así como las estimaciones de β 0, β 1, β 2, β 3, y de sus respectivas probabilidades asociadas a la prueba t, de los 11 modelos aplicados. El valor más alto de R2 lo obtuvo el modelo cúbico (0.262), mientras que el valor más bajo correspondió al sigmoideo (0.013). Después del modelo cúbico, los modelos compuesto, crecimiento, exponencial y logístico, ocuparon el segundo lugar, respecto al valor de R2, con un valor similar de 0.142 cada uno. Estos cuatro modelos mostraron el mismo valor en la prueba F(F = 2.155), siendo el más alto de todos los modelos. Los 11 modelos ajustados a los datos de C de la JA, de la LMT, presentaron valores de β 0 con significancia estadística, de acuerdo a la prueba t (todos diferentes de cero) (P < 0.05), mientras que con respecto al parámetro β 1 solo los modelos compuesto y logístico presentaron valores significativamente diferentes de cero (P < 0.05), según la prueba de t. Estos dos últimos modelos fueron los únicos en donde ambos parámetros de la regresión (β 0 y β 1) fueron significativos estadísticamente (P < 0.05). Finalmente, los valores de los parámetros β 2, de los modelos cúbico y cuadrático, y β 3 del modelo cúbico, no fueron diferentes de cero (P < 0.05).
Tabla 2 Resumenes de los modelos lineales y no lineales, estimaciones de sus coeficientes y sus probabilidades asociadas a la prueba t, respecto al ajuste de la curva de captura anual de la jaiba (Callinectes sapidus) en la Laguna Madre, Tamaulipas, México, durante el periodo 1998 a 2012.
Table 2. Summaries of linear and nonlinear models, estimates of their coefficients and their probabilities associated with the t test, compared to curve setting annual catch of crab (Callinectes sapidus) Laguna Madre, Tamaulipas, Mexico, during the period from 1998 to 2012.

R2 = Coeficiente determinación; F = Prueba F; gl = Grados de libertad; Sig. = Probabilidad asociada a la regresión; En negrita se indican los modelos que cumplieron con la significancia estadística en sus parámetros β 0 y β 1
En la Tabla 3 se observan los valores estimados de CIAc y CIB, y sus correspondientes valores de ∆i y wi, para cada uno de los 11 modelos aplicados. Los modelos compuesto, crecimiento exponencial y logístico presentaron los menores valores de ∆i de CIAc y CIB, y los mayores de wi para ambos criterios, siendo ∆i de CIAc = 0.000, ∆i de CIB = 1.387; wi de CIAc = 18.2 %, y wi de CIB = 12.6 %. Considerando el nivel de ∆i < 2 de plausibilidad, que indica soporte estadístico similar para los modelos que lo cumplan, los modelos que quedaron incluidos en este nivel fueron el compuesto, potencial, sigmoideo, crecimiento, exponencial y logístico, tanto para el criterio CIAc como el criterio CIB; por lo que dichos modelos fueron los que se utilizaron para aplicar el criterio de IMM. Estos seis modelos sumaron el 99.6 % de los wi, de la totalidad de los 11 modelos utilizados inicialmente. La suma del wi de los cuatro modelos que presentaron mejor ajuste (compuesto, crecimiento, exponencial y logístico) fue del 73 %. En consecuencia, es recomendable el nivel Δi <2de plausibilidad para incrementar el wi y utilizar los modelos en el enfoque IMM.
Tabla 3 Selección de modelos lineales y no lineales mediante el uso del Criterio de Información Akaike (1973) (CIA) y del Criterio de Información Bayesiano (CIB) Schwarz (1978), para la modelación de la tendencia de la captura de la jaiba azul (Callinectes sapidus) durante el periodo 1998 a 2012 en la Laguna Madre, Tamaulipas.
Table 3. Selection of linear and nonlinear models using the Akaike Information Criterion (1973) (AIC, for its acronym in English) and Bayesian Information Criterion (BIC, for its acronym in English Schwarz (1978), for modeling the trend of catching blue crab (Callinectes sapidus) during the period from 1998 to 2012 in the Laguna Madre, Tamaulipas.

K = número de parámetros; CIAc = valor Akaike corregido; Δi = Delta; ωi = peso Akaike o CIB. Nota: En el número de parámetros (K) de cada modelo candidato se incluye el parámetro de
En la Figura 1 se presentan las series de tiempo de la C de la JA de la LMT, con datos observados y datos calculados. Estos últimos incluyen los valores promedio, producto de la sumatoria de los valores ponderados de los parámetros de los modelos compuesto, potencial, sigmoideo, crecimiento, exponencial y logístico; y valores máximos y mínimos de los intervalos de confianza al 95 %, obtenidos al utilizar el procedimiento IMM, basado en la aplicación de los criterios de información CIAc y CIB, como criterios de selección de modelos. En la gráfica se aprecia que los valores promedio y de datos observados, quedan incluidos en los intervalos de conianza de los valores estimados, al aplicar ambos criterios de información, CIAc y CIB; lo que redunda en una robustez de la modelización de la tendencia de la captura de la JA en la LMT.
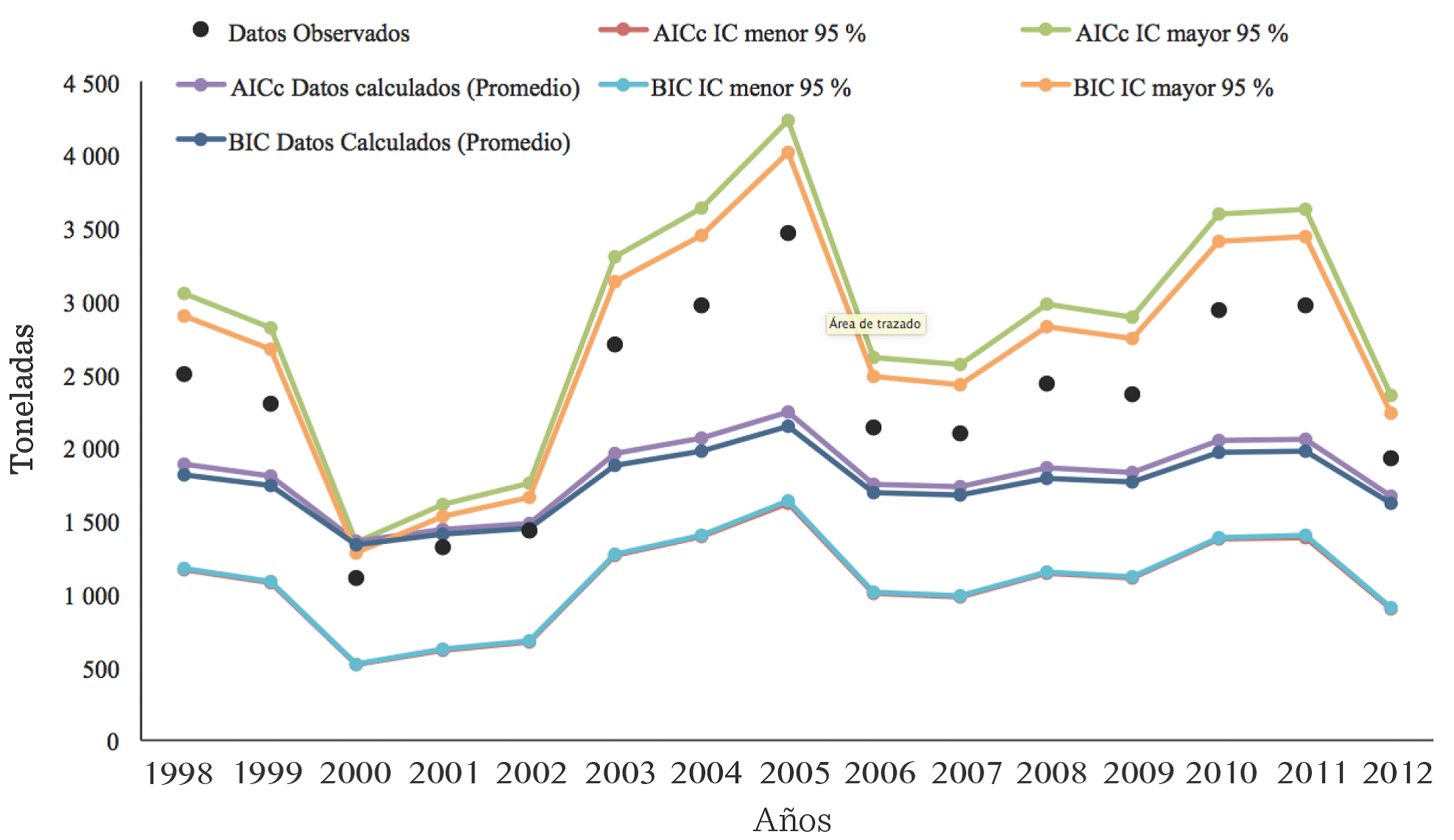
Figura 1 Modelización de la serie de tiempo de la captura de jaiba azul (Callinectes sapidus) en la Laguna Madre, Tamaulipas, México, durante el periodo 1998 a 2012. Se grafican los datos observados, los valores estimados promedios y valores menores y mayores de los intervalos de confianza al 95 %, obtenidos por el procedimiento Inferencia Multimodelo (IMM). Para este procedimiento se utilizaron los valores estimados por los modelos compuesto, potencial, sigmoideo, crecimiento, exponencial y logístico, que fueron elegidos por los Criterios de Información Akaike corregido (CIAc) y Schwarz (CIB).
Figure 1. Modelling time series catching blue crab (Callinectes sapidus) Laguna Madre, Tamaulipas, Mexico, during the period from 1998 to 2012. The observed data are plotted, the estimated values averages and lowest and highest values of confidence intervals 95 %, obtained by the Inference Multimodel (IMM). For this procedure, compound, potential, sigmoid, growth, exponential and logistic models were used, which were selected by the corrected Akaike Information Criterion (AICc) and Schwarz (BIC) values.
En la Tabla 4 se muestran los resultados del procedimiento IMM con base en CIAc y en la Tabla 5 se observan con base en CIB. En este procedimiento se reajustaron los modelos elegidos bajo el nivel Δi< 2 de plausibilidad, en ambos criterios (CIAc y CIB). Excepto los modelos potencial y sigmoideo, el total de los modelos (compuesto, crecimiento exponencial y logístico) presentaron valores similares de los Criterios de Información para el caso CIAc, Δi y ωi (- 7.874, 0.000 y 0.183 respectivamente); y para el criterio CIB, Δi y ωi (1.387, 0.000 y 1.69 respectivamente). Sin embargo, los seis modelos quedaron incluidos en el rango de Δi < 2, por lo que se procedió nuevamente a estimar sus respectivos valores de β0 y β1, calcular sus errores estándar (EE) y valores mayores y menores de sus intervalos de confianza al 95 %.
Tabla 4 Valores de CIAc, Δi, ωi; y valores estimados, error estándar (EE) e intervalos de confianza (95 %) de los interceptos y coeficientes de regresión, dados de forma individual por modelo y en promedio, obtenidos de los modelos ajustados a la captura anual de jaiba azul (Callinectes sapidus), elegidos por dos criterios: mayores valores de ωi CIA, e intercepto y coeficiente de regresión estadísticamente diferente de cero.
Table 4. Values of BIC, Δi, ωi; and estimated values, standard error (SE) and conidence intervals (95 %) of the intercepts and regression coefficients, given individually by model and on average, obtained from the models adjusted annual catch of blue crab (Callinectes sapidus), chosen by two criteria: higher values of wi AICc and statistical significance of the parameters of the regression models (regression coefficient and intercept).

K = cantidad de parámetros; CIAc = Valor Akaike corregido; Δi = Delta; ωi = peso Akaike; EE = Error Estándar; IC = intervalo de confianza; β 0 = intercepto; β 1 = coeficiente de regresión (pendiente).
Tabla 5 Valores de CIB, Δi, wi; y valores estimados, error estándar (EE) e intervalos de confianza (95 %) de los interceptos y coeficientes de regresión, dados de forma individual por modelo y en promedio, obtenidos de los modelos ajustados a la captura anual de jaiba azul (Callinectes sapidus), elegidos por dos criterios: mayores valores de wi CIB y la significancia estadística de los parámetros de los modelos de regresión (intercepto y coeiciente de regresión).
Table 5. Values of BIC, Δi, wi; and estimated values, standard error (SE) and conidence intervals (95 %) of the intercepts and regression coefficients, given individually by model and on average, obtained from the models adjusted annual catch of blue crab (Callinectes sapidus), chosen by two criteria: higher values of wi BIC and statistical signiicance of the parameters of the regression models (regression coefficient and intercept).

K = cantidad de parámetros; CIAc = Valor Akaike corregido; Δi = Delta; wi = peso Akaike; EE = Error Estándar; IC = intervalo de confianza; β 0 = intercepto; β 1 = coeficiente de regresión (pendiente).
Los valores de β0 (939 T y 952 T), de acuerdo a los criterios CIAc (Tabla 4) y CIB (Tabla 5) respectivamente, representan el promedio teórico de las capturas anuales de jaiba, asumiéndose la certeza de que en algún año no existiera captura. El valor se considera teórico porque este escenario no es factible, excepto si se aplicara una veda. El coeficiente β1 con valores de 0.377 y 0.344 (es decir, 37.7 % y 34.4 %), según los criterios CIAc y CIB respectivamente, representan la tasa de cambio anual; es decir, que cada año la captura varía en promedio 377 T o 344 T de JA en la LMT. Los intervalos de confianza para β0 fueron - 0.70 a 2.58 y - 0.71 a 2.61, y los correspondientes para β1 fueron - 0.467 a 1.222 y - 0.471 a 1.158, de acuerdo a los criterios CIAc y CIB respectivamente. Bajo el marco de la incertidumbre estadística, a menor EE mayor precisión de estimadores obtenidos. En este caso, no tiene sentido ubicar el menor EE de los parámetros de la regresión, dado que se contrapone con los ωi: a menor EE menor ωi; y esto ocurre porque el presente análisis es bajo el enfoque IMM que sopesa a cada modelo y genera las bases para una sumatoria de valores, más que la individualización de cada uno.
Es recomendable el uso de criterios de información, en particular CIAc y CIB, dado que ofrecen las ventajas de que pueden ordenar jerárquicamente a los modelos según su ajuste a los datos, y la obtención de sus parámetros promedio. La ordenación jerárquica de los modelos se da en función de los valores CIAc y/o CIB; comparando este valor entre los modelos, y aquel que resulta con el valor más bajo (también reflejado en el menor valor de Δi y en el mayor valor del wi) se considera el mejor modelo (Cailliet, y col., 2006; Romine y col., 2006 y Griffiths y col., 2010). Cabe señalar que la estimación del o los parámetros promedios deseados se estiman obteniendo la ponderación Akaike y Schwarz (Burnham y Anderson, 2002). Esto significa que la contribución de cada parámetro es ponderada antes de obtener el promedio de los parámetros deseados. En este caso, la estimación de β 0 y β 1 y promedios, siguiendo el enfoque IMM, fue de 0.939 según CIAc y 0.952 de acuerdo a CIB, pero no corresponde a un promedio aritmético de los β 0 y β 1 de todos los modelos. En su lugar, es la sumatoria de cada β 0 y β 1 multiplicada por la ponderación (wi %), y con esto se otorga el peso de cada modelo a la estimación de /0 y /1 promedio. Es importante indicar que el uso de R2, ajustada o no, y la suma de cuadrados (SRC), no toma en cuenta la estructura del modelo que mejor se ajusta a los datos, sino que la decisión de seleccionarlo se basa en la complejidad del mismo (Aragón-Noriega, 2013). De hecho, R2 ajustada o no, y la suma de cuadrados, tienden a seleccionar el modelo más complejo (Zhu y col., 2009). Por el contrario, el CIAc se rige bajo el principio de parsimonia y selecciona el modelo más simple, porque penaliza la adición de más parámetros al modelo (Burnham y Anderson, 2002), lo mismo ocurre con el CIB. En este sentido, Burnham y Anderson (2002), expresan que R2 es una medida de la descripción y de la variación del ajuste del modelo, sin embargo, no es un criterio útil para seleccionar un modelo que compita por describir los datos observados. En el presente estudio se mostró esta tendencia: el mejor modelo, de acuerdo a R2 fue el modelo cúbico (R2 = 0.262), pero fue el que presentó una mayor cantidad de parámetros (5), por el contrario: los que fueron incluidos en el enfoque IMM fueron los modelos con solo tres parámetros (Tabla 2).
Es importante indicar que en este estudio se utilizó la varianza residual, obtenida por el cociente de la suma de las diferencias cuadráticas (SSQ), entre el número de datos, en la aplicación de los criterios de información CIAc y CIB. Al utlizar este criterio [uso de la varianza residual en sustitución de la técnica de máxima verosimilitud (MV)] a cada uno de los modelos, además β 0 y β 1, se le sumó otro parámetro, por considerar a la varianza residual (
Índice de Sustentabilidad Pesquera
En el periodo de análisis (1998 a 2012) la producción pesquera de la JA en la LMT, fluctuó entre 1 104 T y 3 462 T, con un promedio anual de 2 310 T (Figura 2). En la serie de tiempo revisada, y de acuerdo al ISP, se identificaron seis periodos de 1998 a 1999, 2000 a 2002, 2003 a 2005, 2006 a 2007, 2008 a 2011, y el año 2012; y los promedios de cada uno de ellos fueron: 2 400 T, 1 285 T, 3 044 T, 2 259 T, 2 954 T y 1 927 T, respectivamente. Las tasas de cambio entre periodos fueron de la forma siguiente: 1° y 2° = - 46 %; 2° y 3° = 137 %; 3° y 4° = - 26 %; 4° y 5° = 31 %; y, 5° y 6° = - 35 %. Se observa que el cambio positivo más grande ocurre entre los periodos 2° y 3°, y el negativo se dio entre los periodos 1° y 2°, y que los cambios que se registran, tanto positivos como negativos, revelan una disminución en su magnitud en la medida que avanza la temporalidad. El ISP es relativamente reciente y permite revisar una panorámica clara y rápida sobre la evolución de la pesquería que se trate.

En términos de evaluación de un recurso pesquero, el análisis de la tendencia de una serie de tiempo, tradicionalmente se recurre a la biomasa de la pesquería, entre otros parámetros, cuando los datos se disponen, y se llegan a expresar puntos de referencia pesqueros en función de los niveles de la biomasa (Cadima, 2003). Sin embargo, también la captura de algunas pesquerías en el mundo ha sido objeto de análisis bajo el enfoque de la tendencia y la identificación de las fases de las pesquerías (Grainger y García, 1996; Froese y Kesner-Reyes, 2002; 2009; FAO, 2010; Froese y Pauly, 2003; Worm y col., 2006; 2007; Pauly y col., 2008; Zeller y col., 2009; Kleisner y Pauly, 2011; Garibaldi, 2012). En particular, estos autores han analizado los datos globales de captura para obtener información sobre el estado de las pesquerías mundiales, revelando por ejemplo, un aumento en los stocks (volumen de biomasa de las poblaciones pesqueras) colapsados y una disminución de nuevos stocks.
Fases de la pesquería
Al aplicar la propuesta de caracterización de la vida de una pesquería de Csirke (1984), a la serie de tiempo de la captura de la JA en la LMT, durante el periodo 1998 a 2012, pero omitiendo la tendencia de la abundancia, se identificaron con sus respectivos periodos, las fases siguientes: fase de colapso: 1998 a 2000; fase de recuperación: 2000 a 2002; fase de crecimiento: 2002 a 2003; fase de explotación plena: 2003 a 2004; fase de sobreexplotación: 2004 a 2005; fase de colapso: 2005 a 2007; fase de recuperación: 2007 a 2009; fase de crecimiento: 2009 a 2010; fase de explotación plena: 2010 a 2011 y fase de colapso: 2011 a 2012 (Figura 3).

Fuente: Modificado a partir de datos de la Subsecretaría de Pesca y Acuacultura del Gobierno del Estado de Tamaulipas.
Figura 3 Fases de desarrollo de la pesquería de la jaiba azul (Callinectes sapidus) en la Laguna Madre, Tamaulipas, durante el periodo 1998 a 2012, de acuerdo al modelo de Csirke (1984)).
Figure 3. Stages of development of the fishery for the blue crab (Callinectes sapidus) in the Laguna Madre, Tamaulipas, México, during the period from 1998 to 2012, according to the model of Csirke (1984).
De igual forma, y de acuerdo al modelo de las fases de una pesquería, propuesto por Caddy (1984), en la pesquería de la JA en la LMT, durante el periodo 1998 a 2012, se identifican las fases en los periodos siguientes: fase IV decadencia: periodo 1998 a 2000; fase I sin desarrollo: periodo 2000 a 2002; fase II en desarrollo: periodo 2002 a 2003; fase III de madurez: periodo 2003 a 2005; fase IV de decadencia: periodo 2005 a 2007; fase I sin desarrollo: periodo 2007 a 2009; fase II en desarrollo: periodo 2009 a 2011; y fase IV de decadencia: periodo 2011 a 2012 (Figura 4).

Fuente: Modificado a partir de datos de la Subsecretaría de Pesca y Acuacultura del Gobierno del Estado de Tamaulipas.
Figura 4 Fases de desarrollo de la pesquería de la jaiba azul (Callinectes sapidus) en la Laguna Madre, Tamaulipas, durante el periodo 1998 a 2012, de acuerdo al modelo de Caddy (1984).
Figure 4. Stages of development of the fishery for the blue crab (Callinectes sapidus) in the Laguna Madre, Tamaulipas, Mexico, during the period from 1998 to 2012, according to the model of Caddy (1984).
La amplitud de la serie de datos y la misma tendencia, permiten registrar dos ciclos de vida de la pesquería de acuerdo a estos modelos, que incluyen las fases de recuperación, crecimiento, explotación plena, sobre-explotación y colapso, en el caso del modelo de Csirke (1984) y las fases de sin desarrollo, en desarrollo, madurez y decadencia, incluidas en el modelo de Caddy (1984).
Un diagnóstico de una pesquería, basado solamente en la captura total, dispone de mayor representatividad a mayor longitud de la serie de datos. Según han señalado Grainger y García (1996) y la FAO (1999), cuando existe una serie histórica suficientemente larga y han ocurrido cambios significativos en la pesquería, el modelo generalizado discutido con anterioridad permite un diagnóstico del estado actual de la pesquería, a partir de la observación de la fase de desarrollo alcanzada. Sin embargo, la serie de datos aquí utilizada, contabiliza más de una década de años, por lo que representa un claro indicio de la posible evolución de la pesquería en cuestión en el periodo de estudio, e incluye la totalidad de las fases de desarrollo que caracterizan a una pesquería.
Una de las características comunes a muchas series de tiempo es la presencia simultánea de una tendencia de largo plazo y una componente cíclica estacional con un periodo fijo (Velásquez y Franco, 2012). La serie de tiempo analizada en este estudio incluyó dos ciclos de "vida" de una pesquería, de acuerdo a los modelos de Csirke (1984) y de Caddy (1984), donde el primero incluyó cinco fases y el segundo cuatro en un periodo de 14 años. Cabe señalar que el concepto de ciclo no tiene una periodicidad fija, y además es un término inapropiado matemáticamente, pero es utilizado porque es el que mejor correspondencia tiene con el uso popular de la palabra "ciclo" (Caddy, 1984).
CONCLUSIONES
Los modelos compuesto, crecimiento, exponencial y logístico, presentaron el mejor ajuste a los datos analizados, de acuerdo a los Criterios de Información Akaike y Bayesiano, los cuales fueron utilizados para estimar el modélo promedio, según el procedimiento de la inferencia multimodelo. Bajo el enfoque de selección de modelos, los mejores modelos para representar la tendencia de la captura de la jaiba azul en la Laguna Madre de Tamaulipas fueron los modelos compuesto y logístico. El índice de sustentabilidad pesquera reveló seis periodos de la serie de tiempo de captura de la jaiba azul de la Laguna Madre Tamaulipas, durante el periodo 1998 a 2012; y una tendencia a la disminución de la magnitud de los cambios de la captura. Durante el periodo 1998 a 2012 se identificaron dos ciclos de vida para esta pesquería. Es importante destacar, que los modelos utilizados señalaron, que en el último año del periodo de estudio (2012), la pesquería de la jaiba se encontraba en una etapa de colapso y en decadencia, por lo que se requiere mantener en estudio la evolución de la pesquería para mantener la sustentabilidad de la actividad económica.

