











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El gasterópodo carnívoro-carroñero P. pansa(Gould, 1853) y el poliplacóforo herbívoro C. (Chiton) articulatus Sowerby in Broderip and Sowerby, 1832, son especies que habitan en la zona intermareal rocosa e interactúan bajo una relación depredador-presa de dicho sistema. Diversos estudios han demostrado, que en la costa de Guerrero, ambas especies son abundantes, dominantes y que presentan altas densidades; en lo que respecta a las longitudes, se ha evidenciado que C. articulatus presenta mayores tallas con respecto a otras especies de moluscos poliplacóforos, mientras que P pansa registra las menores tallas en comparación a distintos trabajos de la misma especie efectuados en otras entidades federativas (Holguín y Michel-Morfín, 2002; Valdés-González y col., 2004; Flores-Campaña y col., 2007; Flores-Rodríguez y col., 2007; García-Ibáñez y col., 2007; Flores-Garza y col., 2012a; Flores-Garza y col., 2012b).
Por otra parte, se destaca el hecho de que ambas especies actualmente se utilizan para el consumo humano (Holguín-Quiñones, 2006; Flores-Garza y col., 2012c; García-Ibáñez y col., 2012; García-Ibáñez y col., 2013; Olea-de la Cruz y col., 2013). Sin embargo, en el caso de P.pansa dicha actividad puede considerarse como furtiva, dado que es una especie protegida por la norma NOM-059-ECOL-2010 (Anónimo, 2010), con un estatus de protección especial.
Diversos estudios enfatizan el hecho de que las características asociadas al sustrato y exposición al oleaje, entre otros factores ambientales, influyen en la diversidad funcional y dinámica de los sistemas ecológicos (Garrity y Leavings, 1981; Rojas, 1988; Barbosa y col., 2009; Aguilera y Navarrete, 2011; Aguilera y Navarrete, 2012; Flores-Campaña y col., 2012; Ávila-Poveda, 2013; Ávila-Poveda y Abadia-Chanona, 2013).
La presente investigación analizó los patrones de respuesta de las especies P pansa y C. articulatus a los factores ambientales de cuatro localidades rocosas, tales como la complejidad y estructura del sustrato, intensidad de oleaje, temperatura promedio del mar, salinidad y clorofila-α, bajo el supuesto de que dada la relativa cercanía de los sitios, las densidades y longitudes totales de las especies son similares.
MATERIALES Y MÉTODOS
Área de estudio
El Estado de Guerrero, se encuentra situado en el sur de la República Mexicana y se localiza entre los 16°18' y 18°48' de latitud norte y los 98°03' y 102°12' de longitud oeste. Carranza-Edwards y col. (1975), mencionan que la costa guerrerense tiene una longitud aproximada de 470lkm y la clasifican como costa de colisión continental. El presente estudio se llevó a cabo en cuatro sitios rocosos costeros del municipio de Acapulco de Juárez, el cual se ubica entre los paralelos 16°41' y 17 13' de latitud norte y los 99°32' y 99°58'de longitud oeste (Figura 1).

La descripción de lugares es como se señala a continuación: 1) Los Pilares: extensión rocosa ubicada entre los 16°49'19.28''N y 99°54'05.01"O, de alta intensidad de oleaje y de alta comple jidad (diaclasas, rugosidades y fracturas); 2) Jaramillo: sitio rocoso de acumulación ubicado entre los 16°52'24.14''N y 99°56'24.94''O, de alta intensidad de oleaje y presencia de bloques de tamaño variable con baja complejidad; 3) Majahua: sitio rocoso de acumulación que se ubica entre los 16°47'41.61''N y 99°50'32.0''O, con baja intensidad de oleaje y bloques de tamaño variable, con baja complejidad; 4) Palmitas: extensión rocosa ubicada entre los 16°49'25.58''N y 99°54'41.51''O, de baja intensidad de oleaje y de alta complejidad (Figura 2).

Materiales
Durante los meses de octubre y diciembre de 2010, así como de febrero y abril de 2011, se realizaron muestreos en la zona intermareal rocosa, en horas de marea baja y en fases de luna nueva. Para delimitar el área de trabajo, en cada sitio se colocó un transecto paralelo a la línea de costa, con anchura de un metro. Se utilizó un cuadrante de 1 m de lado como unidad de muestreo. Se aplicó un diseño sistemático, seleccionando al azar el punto de inicio dentro de tres previamente definidos. Cada cuadrante fue examinado, y con ayuda de una espátula cuidadosamente se retiraron todos los ejemplares de P pansa y C. articulatus, depositándolos en bolsas de plástico separadas y etiquetadas. Al concluir cada unidad muestral o réplica se dejaba un espacio de dos metros y se continuaba con el muestreo, hasta sumar un total de 20 m2 por sitio y fecha. Se registró la longitud total (Lt) de ambas especies con un calibrador digital marca Surtek, (modelo 122204 con precisión 0.001 mm; Shanghai, China).
Para P. pansa, L se determinó partiendo desde la punta del ápice hasta la abertura del canal sifonal, y para C. articulatus, Lt se consideró como el punto más distante entre la valva anal y valva cefálica, tomando en cuenta el cinturón. La determinación de ambas especies se apoyó con literatura especializada (Keen, 1971; Reyes- Gómez, 2004), y la sistemática de ambas especies se actualizó de acuerdo a Skoglund (2002) y Sirenko (2006).
Semanalmente, se hizo el registro de parámetros ambientales, para lo cual se utilizó una sonda multiparámetros YSI (modelo 6920V2; Yellow Springs, Ohio, Estados Unidos), anotando los valores de la temperatura promedio del mar (°C), salinidad (mg/L) y clorofila-α µg/L).
Métodos estadísticos
Para ambas especies, por sitio de estudio se determinó la densidad relativa (org/m2) y su desviación estándar (DE); en el caso de la (Lt) se calcularon el promedio (
Por fecha de estudio se calcularon estadísticos descriptivos como el
Para determinar la relación de las variables ambientales con la frecuencia de aparición y longitudes de ambas especies, se aplicó un análisis multivariado de datos con la técnica de análisis de componentes principales (ACP); dicho análisis se efectúa con una matriz de correlaciones con la finalidad de obtener n componentes ortogonales que permitan la mejor interpretación de la relación entre variables (Tabla 1). Las rutinas estadísticas del ACP consistieron en: I) obtención de una matriz de correlaciones bivariadas y su determinante; II) revisión de la adecuación de la muestra y comprobación de que la matriz de correlaciones no es una matriz identidad, aplicando los estadísticos Kaiser-Meyer-Olkin (KMO) y la prueba de esfericidad de Bartlett, respectivamente; III) determinación de la correlación parcial de variables, a través del cálculo de la matriz anti-imagen y verificación de los valores de la diagonal principal; IV) varianza total explicada (por el modelo), varianza explicada por variable (comunalidades), número de componentes extraídos, y saturación de las variables en los componentes; V) rotación por el método de máxima varianza.

RESULTADOS
Se recolectaron 2 925 ejemplares de ambas especies, de los cuales, 745 correspondieron a la especie P pansa, y 2 180 a C. articulatus (Figura 3a y 3b). Las densidades relativas de cada especie presentaron gran variación entre los sitios de estudio y en todos los casos siempre fueron mayores a 1.0 org/m2. De esta manera, P.pansa registró sus mayores densidades en los sitios Los Pilares y Palmitas (K W, X2 = 43.60, P = 0.0), mientras que C. articulatus las presentó en Jaramillo, Los Pilares y Majahua (K-W, X2 = 27.52, P = 0.0). Con respecto a Lt, en el caso de P.pansase observó que el mayor valor se presentó en el sitio Jaramillo y el menor en el sitio Palmitas (K-W, X2 = 100.30, P = 0.0); para C. articulatus, las mayores longitudes se determinaron en Majahua, Jaramillo y Los Pilares (K-W, X2 = 60.80, P = 0.0) (Tabla 2).



En cuanto a las variables ambientales, se observó que a través del tiempo, tanto la temperatura promedio del mar (TemProm) como salinidad (Salini), presentaron valores promedio mensuales muy cercanos, así como desviaciones estándar pequeñas; caso contrario fue la variable Clorofila-α (Cloro), cuyos valores promedio mensuales tuvieron mayor fluctuación y valores más grandes de la desviación estándar. Dicho comportamiento se corroboró con el coeficiente de variación, donde se observa que el mayor valor lo presentó Cloro, y en el caso de TemProm y Salini fueron pequeños (Tabla 3).

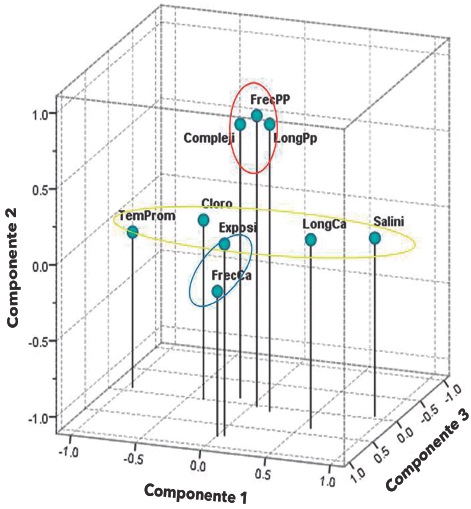
A partir del ACP realizado con nueve variables (Cloro, TempProm, Salini, Exposi, Compleji, FrecPp, FrecCa, LongPp y LongCa), se observó que de 36 valores de correlación bivariada calculados, un total de 19 fueron significativos (P < 0.05), presentándose en una escala de -0.90 a 0.52, donde la relación TempProm-Salini presentó la mayor asociación negativa y LongPp-FrecPp la mayor asociación positiva. El valor del determinante fue de 0.049, sugiriendo linealidad entre variables. El estadístico KMO presentó un valor de 0.54, por lo que el tamaño de muestra se consideró aceptable. La prueba de Esfericidad de Bartlett resultó significativa (X2 =779.700, g.l.=36; P = 0.0), confirmando la relación lineal entre variables. Los valores de la diagonal principal de la matriz antiimagen fluctuaron entre 0.256 (Cloro) y 0.853 (LongCa), lo que evidenció correlación parcial de variables. En función de los autovalores (eigen valores) y la saturación de las variables, se extrajeron tres componentes con una varianza total explicada de 60.55 %, donde el primer componente aportó un 27.92 %, el segundo 18.94 % y el tercero 13.69 %. En lo que respecta a las comunalidades, el menor valor lo presentó Cloro (0.146), y el mayor TempProm (0.917).
El primer componente recibió el nombre de "Ambiente y longitud del herbívoro", en el cual saturaron las variables TempProm, Salini, Cloro y LongCa; al segundo componente se le denominó "Microhábitat, longitud y abundancia del carnívoro" y saturaron Compleji, FrecPp y LongPp; el tercer componente se denominó "Intensidad de oleaje y abundancia del herbívoro" y saturaron Exposi y FrecCa (Tabla 4, Figura 4).

DISCUSIÓN
Tanto el caracol P. pansa como el poliplacóforo C. articulatus presentaron altas densidades en todos los sitios y fechas de estudio, lo que por una parte permitió constatar la dominancia de las especies, así como también observar y comparar los cambios en sus abundancias (temporo-espaciales) como respuesta entre otros aspectos, a las variaciones locales del hábitat en conjunción con el efecto de condiciones del ambiente. A pesar de la relativa corta distancia entre los sitios de estudio, y del aparente efecto similar de las mismas condiciones ambientales, se determinaron diferencias sustanciales en los indicadores biológicos.
En el estado de Guerrero, diversos trabajos han manifestado que tanto el carnívoro-carroñero como el herbívoro, son especies dominantes y abundantes (Tabla 5). Es posible manifestar que los resultados de la presente investigación, coinciden en el hecho de que ambas especies son abundantes y tienen densidades relativas altas. Lo anterior se puede considerar una fortaleza para estudios futuros, ya que la importancia de trabajar con especies dominantes y frecuentes es que se puede observar de manera más detallada su respuesta a las condiciones del hábitat y las variaciones ambientales, de tal forma que cambios en dichos parámetros pueden verse reflejados en variaciones de sus abundancias en una escala temporal y espacial.
Tabla 5 Densidades relativas y longitudes promedio de P. pansa observadas en la costa rocosa de Guerrero, México.
Table 5. Relative densities and average lengths of P. pansa observed on the rocky coast of Guerrero, Mexico.
| Autores | Sitio | Densidad (org/m2) y/o Frecuencia (%) | Longitud promedio (mm) | Observaciones |
|---|---|---|---|---|
| Flores-Rodríguez y col. 2003 | Isla La Roqueta | 100 | No reporta | Especie dominante. |
| García-Ibáñez y col. 2004 | Punta Maldonado | 1.88 | 28.3 | Consideran que la especie presenta la mayor densidad y menor longitud promedio del Pacífico Mexicano. |
| Las Peñitas | 6.94 | 20.1 | ||
| Playa Ventura | 3.42 | 19.6 | ||
| Zoológico | 7.56 | 20.1 | ||
| Palmitas | 9.0 | 17.9 | ||
| Tlacoyunque | 5.0 | 26.7 | ||
| B. de Potosí Expuesto | 3.13 | 26.4 | ||
| B. de Potosí Protegido | 1.63 | 28.0 | ||
| Troncones | 3.75 | 22.2 | ||
| Total | 4.70 | 21.7 | ||
| Valdés-González y col. 2004 | Sitio Expuesto | 9.3 - 12.85 | No reporta | 100 % frecuente en ambos sitios. |
| Sitio Protegido | 6.60 - 9.55 | |||
| Flores-Rodríguez y col. 2007 | Playa Troncones | 100 | No reporta | Especie representativa de la comunidad por su abundancia y frecuencia de aparición. |
| Flores-Garza y col. 2007 | Playa Ventura | 5.19 ± 4.63 | 20.10 ± 6.01 | La población de caracoles del sitio se caracterizó por presentar tallas pequeñas y densidades altas. |
| García-Ibáñez y col. 2007 | Punta Maldonado | 3.59 | No reporta | No hay comentarios al respecto. |
| Las Peñitas | 5.71 | |||
| Playa Ventura | 5.19 | |||
| Zoológico | 10.26 | |||
| Palmitas | 8.34 | |||
| Tlacoyunque | 5.59 | |||
| B. de Potosí Expuesto | 4.14 | |||
| B. de Potosí Protegido | 0.89 | |||
| Troncones | 7.93 | |||
| Torreblanca-Ramírez y col. 2012 | Parque de la Reina | 0.10 | 17.80 | Especie común. |
| Flores-Garza y col. 2012a | Costa Chica | 4.83 | 20.90 | Indican que la población de caracoles se caracteriza por presentar tallas pequeñas y altas densidades. |
| Presente Estudio | 1.23 - 4.65 | 19.84 - 25.86 | ||
Por otra parte, distintas investigaciones han planteado el hecho de que en la costa rocosa del estado de Guerrero, se han registrado las longitudes promedio más pequeñas de P pansa (Tabla 5), situación que actualmente se observa, ya que el promedio determinado en el presente trabajo es similar a los valores reportados. En el caso de la especie C. articulatus, estudios realizados en la costa del estado de Guerrero, mencionan que sus ejemplares alcanzan las mayores tallas en comparación con otros moluscos poliplacóforos, reportando longitudes promedio que pueden ser consideradas como similares a las obtenidas por la presente investigación; sin embargo, también se destaca que las longitudes obtenidas resultaron ser más pequeñas al compararlas con otras registradas a lo largo del Pacífico Mexicano (Tabla 6). De esta manera, puede decirse que la costa del Estado de Guerrero, se caracteriza por presentar altas densidades y longitudes promedio pequeñas de ambas especies.
Tabla 6. Densidades relativas y longitudes promedio de C. articulatus observadas en la costa rocosa del Pacífico Mexicano.
Table 6. Relative densities and average lengths of C. articulatus observed on the rocky coast of the Mexican Pacific.
| Autores y entidad federativa | Sitio | Densidad (org/m2) y/o Frecuencia (%) | Longitud promedio (mm) | Observaciones |
|---|---|---|---|---|
| Holguín y Michel-Morfín 2002 (Colima) | Bahía Binners | 3.72 | 67.9 | Molusco abundante con tallas más grandes que en Acapulco. |
| Bahía Braithwaite | 8.48 | 66.5 | ||
| Bahía Grayson | 3.86 | 75.6 | ||
| Bahía Academia | 4.38 | 71.9 | ||
| Total | 5.3 | 69.6 | ||
| Valdés-González y col., 2004 (Guerrero) | Isla La Roqueta | 0.50 - 1.85 | No reporta | Especie común y dominante en el sitio expuesto. |
| Flores-Campaña y col., 2007 (Sinaloa) | Isla Venados ZP | No reporta | 39.4 | Tendencia de mayores tallas en zonas expuestas. |
| Isla Venados ZE | 42.9 | |||
| Isla Pájaros ZP | 42.6 | |||
| Isla Pájaros ZE | 51.7 | |||
| Total | 44.1 | |||
| Flores-Rodríguez y col., 2007 (Guerrero) | Playa Troncones | 80 | No reporta | Especie dominante. |
| Flores-Garza y col., 2007 (Guerrero) | Playa Ventura | 1.71 | No reporta | Especie dominante. |
| Flores-Garza y col., 2011 (Guerrero) | Playa Majahua | 0.95 | 25.19 | Especie dominante y poliplacóforo con la mayor talla. |
| Flores-Rodríguez y col., 2012 (Guerrero) | Playa Carrizalillo | 2.38 | No reporta | Poliplacóforo de mayor densidad. |
| Playa Coral | ||||
| Playa Cometa | ||||
| Galeana-Rebolledo y col., 2012 (Guerrero) | Playa Tlacopanocha | 0.05 | 25.34 | Especie de poliplacóforo ocasional y de mayor talla. |
| Torreblanca-Ramírez y col., 2012 (Guerrero) | Parque de la Reina | 1.00 | 21.66 ± 1.07 | Especie dominante y poliplacóforo de mayor longitud. |
| Flores-Garza y col., 2012b (Guerrero) | MPR No. 32 | 1.89 | 22.83 | Especie dominante de mayor longitud y amplia distribución. |
| Presente Estudio | 4.54 - 14.55 | 18.78 - 24.91 |
Para coadyuvar en la explicación del hecho anterior, García-Ibáñez y col. (2004), mencionan que en el caso de P. pansa existe un fenómeno de denso-dependencia ligado al hábitat disponible y densidad relativa de la especie, de tal manera que a mayor densidad se observan menores tallas. Por otra parte, algunas investigaciones han manifestado que ambas especies son capturadas para el consumo humano (Flores-Garza y col., 2012c; García-Ibáñez y col., 2012), y que en el caso de C. articulatus existe un efecto de la pesca que se expresa en cambios en la distribución de frecuencias de tallas (Holguín y Michel-Morfín, 2002; Flores-Campaña y col., 2007), lo que también se refleja en la reducción progresiva de las longitudes de captura (García-Ibáñez y col., 2013), situación que incluso se ha reportado en otras entidades federativas (Holguín-Quiñones, 2006). En el caso de las regiones costeras de Acapulco y Costa Grande del Estado de Guerrero, Olea-de-la-Cruz y col. (2013), manifiestan que la especie C. articulatus es ampliamente demandada, ya que su oferta en distintos puntos de venta es considerada como frecuente; y en lo que corresponde a P. pansa, se destaca que la especie se encuentra en protección especial bajo la norma oficial mexicana NOM-059-ECOL-2010 (Anónimo, 2010), sin embargo, los autores de este trabajo han observado su consumo tanto en restaurantes como en expendios de Acapulco.
En el presente estudio fue posible establecer que los factores Ajos, como la complejidad del sustrato rocoso, así como la intensidad de oleaje, influyen en la densidad relativa y longitudes de ambas especies, no obstante, a simple vista no resulta fácil reconocer el gradiente o jerarquía de participación de los factores. De esta manera, en el caso del gasterópodo se observó que sus mayores densidades se presentaron en dos sitios que difieren en cuanto a la intensidad de oleaje, pero que se caracterizan por tener un sustrato rocoso de alta complejidad; aunque en lo que respecta a sus tallas la situación no fue tan precisa.
En cuanto a C. articulatus, los mayores valores tanto de densidad relativa como de la longitud total se presentaron en tres sitios; sin embargo, no existió una correspondencia directa entre las características del sitio con ambas variables, es decir, que las mayores tallas del molusco poliplacóforo no necesariamente se relacionaron con la mayor densidad relativa. Por lo tanto, la comparación exclusiva de promedios de la densidad y tallas de ambas especies en función de características del sustrato y de oleaje, no permitieron una definición de grupos precisa.
Sin embargo, al incorporar otras variables como las ambientales y realizar un análisis multivariado, se observó con mayor precisión su efecto sobre los parámetros biológicos de las especies, lo cual se considera que fue debido a que pudo ser incorporada no sólo la correlación lineal entre variables, sino también su correlación parcial.
De esta manera, en lo que respecta al componente "Ambiente y longitud del herbívoro", se destaca que a un decremento de la temperatura superficial del mar y del estimador de la productividad primaria (clorofila-α), se observó un incremento en la salinidad, así como en la longitud del C. articulatus. Lo anterior exhibe dos situaciones: 1) por las fechas de realización del estudio, la conclusión del período de lluvias y huracanes y el inicio del período de sequía o estiaje, y 2) bajo la óptica de valores máximos y mínimos, un evento previo de altos niveles de productividad primaria, fue base para la alimentación de C. articulatus en dicha temporada, lo que repercutió en el paulatino incremento en longitud del herbívoro.
En términos reproductivos, Rojas (1988), mencionó que a partir de una longitud total de 40 mm, es muy frecuente encontrar ejemplares sexualmente maduros; Ávila-Poveda y Abadia-Chanona (2013), comentan que los machos alcanzan su madurez sexual en tallas más pequeñas con respecto a las hembras, y para propósitos prácticos, proponen una clasificación donde puede considerarse que en ambos sexos los estadios subjuveniles, juveniles y subadultos se observan en tallas menores a 40 mm, y los adultos se registran en tallas iguales o mayores a 40 mm. Por lo anterior, es posible no sólo relacionar los cambios de las variables ambientales con el crecimiento del molusco, sino además, se podrían predecir los cambios en las fases de maduración sexual. Por lo tanto, el presente estudio coincide con trabajos como los de Ishiyama (1995), Barbosa y col. (2009) y Ávila-Poveda (2013), los cuales resaltan el efecto de la temperatura del mar en especies de moluscos poliplacóforos, particularmente en sus épocas reproductivas.
En el componente "Microhábitat, longitud y abundancia del carnívoro", se observó la relevancia de la estructura y complejidad de los sitios rocosos, ya que a mayor discontinuidad del sustrato se encontró una mayor cantidad de ejemplares del carnívoro y donde a su vez, se registraron las mayores longitudes. A este respecto, Garrity y Levings (1981), remarcaron la importancia de grietas húmedas y microhabitats sombreados en la abundancia de P. pansa. Valdés-González y col. (2004), registraron altas densidades del gasterópodo en dos sitios García-Ibañez y col. (2014). Respuesta de Plicopurpurapansay Chiton articulatusa factores ambientales de alta y baja intensidad de oleaje, respectivamente, pero que tienen en común sustratos complejos. Otros trabajos también han destacado la influencia de la complejidad topográfica del hábitat en las abundancias de especies de moluscos intermareales y su distribución, depredación y diversidad biológica, como los de Beck (2000), Chapman (2000), Giraldo y col. (2002) y Fernández y Jiménez (2006). De esta manera, el presente trabajo coincide con los estudios anteriormente citados y sugiere que una característica estrechamente relacionada con la abundancia de P. pansa, corresponde a la estructura y complejidad del hábitat de cada sitio rocoso, lo cual en el caso de los sitios estudiados en Acapulco, Guerrero, presenta una jerarquía mayor a la intensidad de oleaje.
En lo que respecta al componente "Intensidad de oleaje y abundancia del herbívoro", se destaca que una mayor energía del oleaje de cada sitio rocoso se correspondió con una mayor abundancia del C. articulatus. Dos explicaciones pueden sugerirse al respecto: 1) en la zona de estudio, un sitio expuesto al oleaje intenso puede presentar (en función de su pendiente o inclinación), una mayor área húmeda debido al proceso constante de inmersión y emersión de la superficie, así como del rocío que baña la zona supralitoral, de esta manera, y aunado a la cantidad de luz solar, será posible un área mayor de forrajeo, descanso y protección, que repercutirá en una mayor colonización y abundancia de organismos, entre ellos los productores; 2) otra suposición no necesariamente excluyente de la anterior, es el hecho de la dificultad de la pesca del molusco poliplacóforo en zonas de alta energía de oleaje. En este sentido (Holguín y Michel-Morfín, 2002; Flores-Campaña y col., 2007; Olea-de-la-Cruz y col., 2013), han señalado el efecto que generan en la pesca del poliplacóforo, el oleaje y topografía de la zona intermareal rocosa, así como factores climáticos como viento y lluvia. Estudios como el de Fernández y Jiménez (2006), Nishida y col. (2006) y Aguilera y Navarrete (2011), también remarcan el efecto de la intensidad de oleaje con la obtención de alimento por parte de invertebrados herbívoros. De esta manera, el caso de la especie C. articulatus puede abordarse a partir de dos situaciones: I) A nivel temporal, las longitudes del molusco poliplacóforo dependen de la productividad primaria y condiciones físico-químicas (temperatura y salinidad) del mar; II) A nivel espacial, las abundancias dependen en mayor grado de las condiciones del sitio rocoso asociadas con una alta energía de oleaje.
Flores-Campaña y col. (2012), manifiestan que es necesaria más información sobre la influencia de factores ambientales para generalizar patrones sobre la forma y tamaño del molusco poliplacóforo C. (Chiton) albolineatus Broderip and G. B. Sowerby I, 1829, así como en su distribución y abundancia. Olabarria y col. (2001), comentan que la estructura de la comunidad de moluscos en sustratos rocosos se determina por interacciones complejas entre factores bióticos y abióticos.
En términos de biología evolutiva, tanto P. pansa como C. articulatus presentan adaptaciones morfológicas y fisiológicas (tamaño del pie, poca o nula presencia de nódulos o espinas, forma ovalada y dorsoventralmente aplanada, entre otras) para coexistir en las zonas intermareales rocosas, siendo por lo tanto organismos que pueden demarcar de manera efectiva la zona mesolitoral, debido a la dominancia, abundancia y densidad relativa que presentan.
La importancia del estudio de las relaciones funcionales en el espacio y tiempo, es debido a que dichas relaciones se modifican en función de cambios en la sucesión de la comunidad (Aguilera y Navarrete, 2012).
En base a lo anterior, es posible comentar que tanto el gasterópodo como el poliplacóforo objeto de estudio, son especies que pueden coadyuvar en el entendimiento de la estructura funcional del sistema intermareal rocoso en Acapulco, Guerrero, y por ende pueden ser especies indicadoras de cambios en el ambiente.
CONCLUSIONES
El caracol P. pansa, así como el poliplacóforo C. articulatus, son moluscos que en la zona intermareal rocosa de Acapulco, Guerrero, presentan densidades y longitudes que varían en función de características ligadas a la complejidad del sustrato y exposición de la costa rocosa. Al relacionar los indicadores biológicos de las especies con características locales del sitio, así como con parámetros ambientales del área de estudio, se observó una mejor definición de la relación funcional de las especies con su medio, donde se resalta que la longitud del poliplacóforo se encuentra estrechamente relacionada con la temperatura del mar, salinidad y productividad primaria (expresada en términos de la clorofila-α), así también que tanto la abundancia y longitud del gasterópodo se relacionan con una mayor complejidad del sustrato rocoso; y por último, que las abundancias del poliplacóforo están mayormente ligadas con las condiciones del sustrato rocoso asociadas con una alta intensidad de oleaje.

