











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La belleza natural de las costas de la Península de Yucatán ha motivado un desarrollo turístico que día a día se incrementa y consecuentemente influye en la conservación de sus recursos. Entre los impactos ambientales originados por el turismo en el Caribe Mexicano y particularmente en Cancún, cabe mencionar la contaminación de sus playas y aguas costeras, la alteración o disminución de hábitat críticos, como son los manglares, sistemas arrecifales y la explotación de recursos pesqueros. Estas alteraciones reducen la complejidad ambiental y consecuentemente la biodiversidad.
Preocupada por esta problemática, la Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT), a través de la Comisión Nacional de Áreas Naturales Protegidas (CONANP) y el Centro de Educación y Capacitación para el Desarrollo Sustentable (CECADESU), en coordinación y colaboración con asociaciones civiles en pro de la protección ambiental y conservación de los recursos, han realizado esfuerzos para que áreas costeras de riqueza biológica sean establecidas como áreas naturales protegidas (INE, 1997). De esta manera, se resuelven conflictos entre los diversos usuarios y se contribuye a restablecer poblaciones sobreexplotadas o áreas degradadas (Agardy, 2000). Asimismo, se ha señalado como prioritario el proteger y restablecer las especies en peligro (por ejemplo: flamencos, manatís), evaluar los hábitat críticos (por ejemplo: humedales) y regular la presión acelerada que se está ejerciendo sobre los recursos de valor pesquero como la langosta, caracol y algunas especies de escama (Gaines y Costello, 2013). Dentro de estos esfuerzos, las áreas naturales protegidas se consideran una estrategia para la conservación y el mantenimiento de la biodiversidad de un país, así como para el aprovechamiento sustentable de sus recursos naturales (Andam y col., 2010).
Recientemente, el ecoturismo ha llegado a ser una importante derrama económica para los pobladores de Quintana Roo (Ramírez-Macías y col., 2012). En particular, la belleza y biodiversidad de Isla Contoy, y el hecho de constituir zonas de anidación y refugio para aves costeras, son factores de gran atracción para el turismo nacional e internacional, que al complementarse con información ecológica cumple su función, que es la de dar a conocer un recurso natural como un patrimonio nacional y apreciar su valor para conservarlo. En 1961 se decretó Isla Contoy (Isla Pájaros) como Parque Natural y Refugio de Fauna Nacional por constituir una zona de refugio, alimentación y anidación de diversas aves marinas, reptiles en peligro de extinción (tortugas), y gran número de especies de peces e invertebrados. Posteriormente, el 2 de febrero de 1998, se declaró en el Diario de la Federación como "Parque Nacional". Esta reserva se caracteriza por la presencia del buen estado de conservación de sus ecosistemas y en el caso de modificaciones, éstas han sido debidas a fenómenos naturales (huracanes, tormentas tropicales).
La investigación sistemática y biológica de las especies permite asegurar su conservación, debido a la amenaza constante de numerosas actividades humanas que están deteriorando los hábitat críticos e introduciendo especies exóticas que desplazan a las nativas (Greenwood, 1992). Como ejemplo reciente se tiene la invasión del pez león (Pterois milesy P. volitans) en las zonas arrecifales y hábitat asociados en el Atlántico tropical, siendo una de las principales amenazas para las especies nativas de esta región (Morris, 2012).
Entre las comunidades biológicas más estudiadas dentro del Parque Nacional Isla Contoy, se tiene la flora terrestre con dos tipos de vegetación: halófila o duna costera y manglar (Cabrera y col., 1982). De las faunísticas, se ha evaluado su valor para la crianza y alimentación de especies de aves permanentes y migratorias, como el pelícano gris, halcón peregrino, pájaro bobo, cormorán, fragata magnífica y garza gigante (Blankinship, 1986; Paul, 1991). Respecto a la ictiofauna, Schmitter-Soto y col. (2000), mencionan que la región caribeña está representada por 577 especies, siendo una de las provincias mexicanas con mayor diversidad debido a la presencia de arrecifes y por compartir más de 30 especies con el Atlántico oriental.
Se han realizado diversos estudios en la zona arrecifal de Veracruz (González-Gándara y col., 2012) y el Caribe Mexicano (Nuñez-Lara y Arias-González, 1998; Chitarro y col., 2005; Yeager y Arias-Gonzalez, 2008), pero la fauna íctica del Parque Nacional Isla Contoy es poco conocida. A este respecto, Villalobos (1979), realizó un inventario de la fauna isleña en general, en el que reporta a 12 especies de peces coralinos. Sin embargo, las zonas lagunares y costeras, cercanas a las zonas de anidación de las aves marinas, no han sido evaluadas desde un punto de visto ictiológico. Se carece de una descripción y evaluación cuantitativa de las poblaciones de peces que habitan estos ambientes. Este estudio tiene como objetivo contribuir al conocimiento taxonómico de las especies de peces y determinar la estructura de sus asociaciones en función de la variabilidad hidrológica de los sistemas lagunares y costeros del Parque Nacional.
MATERIALES Y MÉTODOS
Área de Estudio
El Parque Nacional de Isla Contoy se localiza en el Estado de Quintana Roo, México; (Figura 1a) forma parte del conjunto de Islas, Bancos y Arrecifes de la plataforma continental del Caribe junto con Isla Mujeres, Cayo Sucio, Isla Blanca y el Banco Arrowsmith. Se ubica al sureste de México, a 12.5 km de la costa oriental de la Península de Yucatán, a 30 km al norte de Isla Mujeres, y a 32.3 km de Cabo Catoche, con una superficie total de 238.2 ha, de las cuales, 96.6 % corresponde a tierra firme y pequeños islotes, y 3.4 % a lagunas interiores (INE, 1997). El clima es cálido subhúmedo con lluvias en verano, temperatura media de 27.4 °C con escasa oscilación mensual. La temperatura más alta se presenta en el mes de agosto, y la más baja en enero, con una diferencia de 3 °C a 4 °C. La precipitación anual es de 1 041 mm, es escasa de enero a abril, se incrementa de mayo a junio, disminuye durante julio y agosto y presenta un máximo en septiembre. De noviembre a febrero la presencia de huracanes o ciclones pueden acarrear intensas precipitaciones.
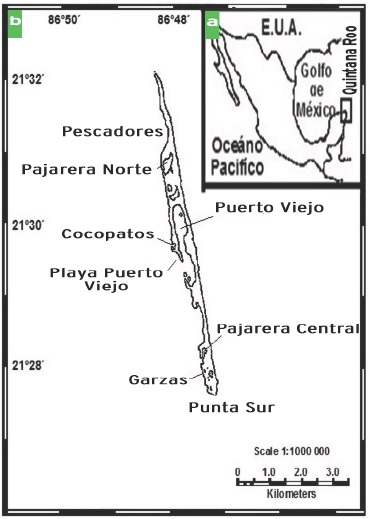
El Parque Nacional Isla Contoy tiene longitud de 8.8 km, y anchura desde 20 m (norte) hasta 700 m (centro). La costa oriental está dispuesta a barlovento, constituida en gran parte por zonas rocosas erosionadas por el oleaje y vientos dominantes del sureste. Por el contrario, la costa Este dispuesta a sotavento, presenta una alternancia de zonas rocosas y arenosas entre las que destacan de norte a sur: Playa de Cocos, Playa Pájaros, Playa Pescadores, Playa Ixmapoit, Playa Tortugas y Playa Palmas. Sobre la misma costa, numerosas playas están cortadas por la boca de varios sistemas lagunares; dentro de los principales en dirección norte a sur se encuentran: a) Laguna Norte, que se comunica con el mar por una boca (8 m de ancho) y con Laguna Pajarera Norte; b) Laguna Pajarera Norte (Figura 2), con aguas poco profundas en proceso de eutroficación, debido al guano de las aves que anidan en las riveras y de la materia orgánica en descomposición proveniente de los manglares circundantes; c) Laguna Muerta, actualmente en proceso de desecación y muerte por la hipersalinidad de sus suelos; d) Laguna Puerto Viejo, la de mayor extensión, presentando una amplia boca y tres islotes en su interior; e) Laguna Pajarera Central (Figura 3), con alta transparencia cerca del canal y turbidez por eutroficación en la parte este; f) Laguna Garzas y g) Pajarera Sur, bordeadas por manglar con aguas someras y turbias.


Isla Contoy carece de cuerpos de agua dulce y corrientes superficiales. Sin embargo, presenta infiltración del agua de lluvia al subsuelo (Vega-Cendejas y Hernández, 2002). La vegetación insular, principalmente sobre la línea de costa y bordes de las lagunas, se encuentra representada por especies de manglar rojo (Rizophora mangle) y negro (Avicennia germinans), y en menor cantidad por el blanco (Laguncularia racemosa) y mangle botoncillo (Conocarpus erecta). Estas especies están asociadas con vegetación de tipo matorral (Suriana maritima, Tournefortia gnaphalodes, Cordia sebestana) y cocoteros introducidos. La vegetación marina del lado oriental está representada por Sargassum, y de la costa poniente por grandes extensiones de ceibadal (Thalassia testudinum) asociada con Syríngodium filiforme (Souza y Cabrera, 1983).
Trabajo de campo y muestreo
El área de muestreo comprendió únicamente el poniente de la Isla debido al fuerte oleaje y erosión que impidieron la realización del trabajo de campo en la zona oriental. Durante aproximadamente un año (abril de 1999 a enero de 2000) se realizaron recolectas bimensuales en ocho sitios (Figura 1b), localizados en los sistemas lagunares de Puerto Viejo, Pajarera Central, Pajarera Norte y Garzas, así como en las localidades de la zona costera en Playa Puerto Viejo, Cocopatos, Pescadores y Punta Sur. Para ello se tomó siempre en cuenta el no perturbar las zonas de reproducción y anidación de las aves (Permiso de Pesca de Fomento No. 290799-213-03, expedido por la Dirección de Vida Silvestre de la SAGARPA). En el caso de Puerto Viejo, se ubicaron tres localidades que se consideraron réplicas (boca, zona de anidación y sardinera) y dos en Pajarera Central (boca y zona interna).
En campo, previo a la toma de muestras biológicas, se registraron en cada sitio de colecta la temperatura, oxígeno disuelto y salinidad, utilizando un equipo Yellow Spring Instrument (YSI) modelo 51 (Ohio, EUA), el pH con un check mate marca TM HI 9126 (EUA) y las coordenadas con un GPS marca Garmin (PPS12XL, EUA). Adicionalmente, se tomaron muestras de agua que fueron etiquetadas y puestas en hielo, para posteriormente en el laboratorio de química marina del Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional (CINVESTAV Unidad Mérida) realizar la determinación de nutrientes (nitritos, nitratos, fosfatos y silicatos), mediante los métodos reportados en Strickland y Parson (1972).
Para la recolecta de los peces se usó una red de arrastre tipo camaronera (4lm abertura vertical y 2.5 cm malla) en los canales de navegación (2 m a 3-m profundidad), y un chinchorro (30 m x 2 m, 2.5 cm de malla), en las zonas de menor profundidad (60 cm a 80cm). En cada sitio se realizaron por lo menos dos lances con un tiempo promedio de 45min cada uno, siguiendo los lineamientos sugeridos por Moyle y Cech (2004). Los organismos recolectados se conservaron en formaldehído (10 %), y se etiquetaron con los datos de recolecta (recolector, método, fecha, hora, número de estación), en formatos específicos que permitieron su identificación y seguimiento en su traslado al laboratorio.
Adicionalmente, se tomaron fotografías de los ejemplares cuya coloración en fresco se considera esencial para la identificación taxonómica.
Trabajo de Laboratorio y análisis de datos
Los ejemplares recolectados, después de haber sido sometidos a un proceso de lavado, se identificaron utilizando bibliografía especializada (Fisher, 1978; Dickson y Moore, 1998; McEachran y Fechhelm, 2005). El orden sistemático se basó en el criterio de Greenwood y col. (1967), modificado por Nelson (2006). Se registró individualmente el peso y la longitud estándar (LE) de cada espécimen a fin de elaborar las bases de datos correspondientes. Los peces se dividieron en cuatro categorías ecológicas de acuerdo a su origen y tolerancia a la salinidad como eurihalino, estenohalino, estuarino y especies de agua dulces (Day y col., 1989; Castro-Aguirre y col., 1999).
La similitud de los parámetros ambientales (temperatura, O2, pH, conductividad, NH4 NO2, NO3, K, SiO4) entre sitios, se analizó utilizando el índice de similitud de distancias Euclideanas con la matriz de los parámetros transformados (raíz cuarta) (Clarke y Warwick, 2001). Las diferencias se evaluaron utilizando las pruebas de análisis de similitud (ANOSIM). La composición y abundancia de las especies se estimó agrupando los especímenes recolectados con ambos artes de pesca. Se utilizó una combinación de análisis multivariados para evaluar las diferencias entre los parámetros registrados y la composición de los ensamblajes en cada sitio de muestreo, así como su relación con las variables ambientales. La estructura de la comunidad se analizó espacialmente por medio de sus principales descriptores (riqueza, dominancia, diversidad y equidad). La riqueza (S), se estimó como el número total de especies por localidad. La diversidad de especies (H'), que combina el número y distribución de la abundancia entre ellas, se determinó a partir del índice de Shannon-Wiener (bits/individuo), cuyo valor se incrementa con el número de especies y una mayor distribución de los individuos. La equidad (J'), se refiere a la distribución de la abundancia entre las especies y se obtiene a partir de la relación entre la diversidad observada H' y la máxima H' max (Ludwig y Reynolds, 1988; Krebs, 2009). Los valores de J' pueden tener un intervalo de prácticamente cero (no equidad), a uno (completamente iguales).
La dominancia, que implica la jerarquización y evaluación de la importancia relativa de las especies en la estructura de la comunidad, se determinó en una base anual a partir del índice de valor de importancia (IVI), en donde se incorpora información acerca de la densidad (DR), frecuencia (FR) y biomasa relativas (BR), de cada especie registrada (IVI = DR + FR + BR) (Brower y col., 1989). El IVI puede tomar valores de 0 a 3 (o 300 %), dividiendo el IVI entre 3 (100 %) indica el porcentaje de importancia de las especies en una comunidad. Para este estudio se consideraron como dominantes las especies que en conjunto constituyeron el 75 % del IVI.
La matriz de la densidad por sitio de colecta se transformó a raíz cuarta para cumplir con los supuestos de normalidad y homocedasticidad. La contribución de cada especie por sitio se realizó utilizando la prueba estadística SIMPER (porcentaje de similitudes), la cual determina la contribución porcentual de cada especie (densidad) para tipificar un grupo (similitud), o bien discriminar especies entre grupos (disimilitud). El grado de similitud entre sitios se evaluó a través de un análisis de escalamiento no métrico multidimensional (NMDS), con las especies que representaron más del 95 % de la densidad total. Para ello, se utilizó el índice de disimilitud de Bray Curtis, como medida de distancia para realizar la matriz triangular, por el método aglomerativo de unión promedio. La bondad de ajuste se verificó teniendo en cuenta el valor del stress. Este valor hace referencia a la calidad de la representación: bajos valores (< 0.15), indican un buen ajuste (Clarke y Warwick, 2001). El NMDS es un método robusto, que construye un diagrama de ordenación de los n sitios en un número específico de dimensiones, intentando satisfacer todas las condiciones impuestas por el rango de la matriz de similitud, es decir, que las distancias entre las estaciones en el espacio k dimensional se parezca lo más posible a las distancias observadas entre las estaciones de la matriz, de tal manera, que las estaciones afines en composición y abundancia se localizarán más cerca una de la otra. Para determinar las diferencias entre la composición y abundancia de las especies se usuaron las pruebas estadísticas de ANOSIM. Los análisis anteriores se obtuvieron con el programa estadístico PRIMER 6 Plymouth Routines in Multivariate Ecological Research (Clarke y Gorley, 2006). Se realizaron análisis de varianza (ANOVA) y pruebas de T para evaluar las diferencias en la riqueza y diversidad de especies entre los sitios de muestreo. La relación de las especies con los parámetros ambientales se estableció a través del coeficiente de correlación de Pearson (Ludwing y Reynolds, 1988).
RESULTADOS
Hidrología
No se registraron diferencias significativas entre los parámetros ambientales de los sistemas costeros (ANOSIM, R = 0.09, P = 0.16). Respecto a la temperatura, el valor medio registrado (27.8 °C), muestra las condiciones tropicales de los cuerpos de agua estudiados con el valor máximo promedio de 30.5 °C, el cual se registró en Playa Puerto Viejo, y el mínimo (23.4 °C) en Pajarera Central. En forma similar, la salinidad presentó poca variación entre sitios con condiciones prácticamente marinas. Sin embargo, se registraron valores cercanos a 40 en las Lagunas Puerto Viejo y Pajarera Central y Playa Puerto Viejo (Tabla 1). En Pajarera Central también se registraron valores bajos de oxígeno disuelto (4.7 mg/L), altos de fosfatos (3.1 µM), amonio (5.3 µM). Respecto a los nitratos y nitritos, se determinaron valores mínimos en todos los sitios de estudio (Tabla 1). Tanto la Laguna como la Playa en Puerto Viejo, se caracterizaron por presentar valores máximos de silicatos y una alta asociación con la salinidad, mientras que Cocopatos y Pajarera Norte con los nitritos y fosfatos (Figura 4). Por el contrario, Pajarera Central y Garzas presentaron niveles bajos de oxígeno disuelto (Tabla 1). Por otra parte, Punta Sur no presentó ninguna relación con los nutrientes; se caracterizó por presentar aguas transparentes.
Tabla 1 Parámetros hidrológicos promedio ± (desviación estándar) registrados en las estaciones de muestreo durante el período de estudio (abril 1999 a enero 2000). Temp = temperatura, Salinidad, Profundidad, O2= oxígeno disuelto, NH4= amonio, NO2= nitritos, NO3= nitratos, PO4= fosfatos, SiO4= silicatos.

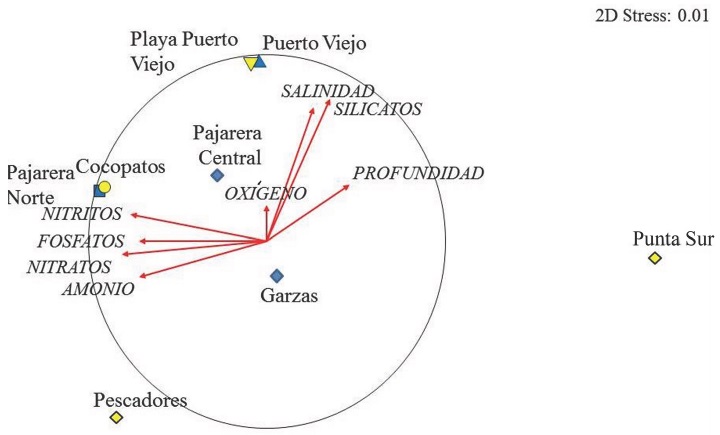
Figura 4. Análisis no métrico multidimensional (NMDS) derivado de una matriz de similitud Bray Curtis construida a partir de los datos de la abundancia de las especies (transformación raíz cuarta) en los diferentes sitios en relación con las variables ambientales registradas en el Parque Nacional de Isla Contoy, Quintana Roo, México.
Figure 4. Non-metric multidimensional scaling ordination plot (NMDS) derived from Bray-Curtis similarity matrix constructed from the fish abundance data (fourth root transformation) in different sites in relation with environmental variables in the National Park of Isla Contoy, Quintana Roo, Mexico.
Contribución de las especies a los ensamblajes
De los 64 órdenes de peces propuestos para el mundo por Nelson (2006), en la Reserva de Isla Contoy se han registrado 15 (23 %), los cuales agrupan a 40 familias, que corresponden al 7.2 % del total a nivel mundial en base a Eschmeyer (2014), y a un registro de 62 géneros y 92 especies (Tabla 2). Estos datos hablan por sí mismos de la alta diversidad íctica de la Reserva, que se refleja en los registros de especies de los componentes eurihalino, estenohalino, estuarino y dulceacuícola (Tabla 3). Dentro de los órdenes registrados, el de los Perciformes fue el que alcanzó el mayor número de familias (17). Las familias más representativas por comprender el mayor número de especies pertenecen a este orden: Scaridae con 10 especies incluidas en cuatro géneros y Gerreidae con un total de seis especies; y al orden Gasterosteiformes: Syngnathidae con cinco especies. El patrón observado de ocurrencia de especies de las Familias Cichlidae y Poeciliidae, sugiere la presencia de descargas puntuales de agua dulce (afloramientos), en la zona lagunar y costera de isla. Cabe resaltar que estas especies son de un origen dulceacuícola secundario, es decir que han evolucionado en el agua dulce, pero se han originado de grupos marinos y son capaces de cruzar pequeñas regiones marinas (Lowe-McConnell, 1987; Vega-Cendejas y col., 2013).


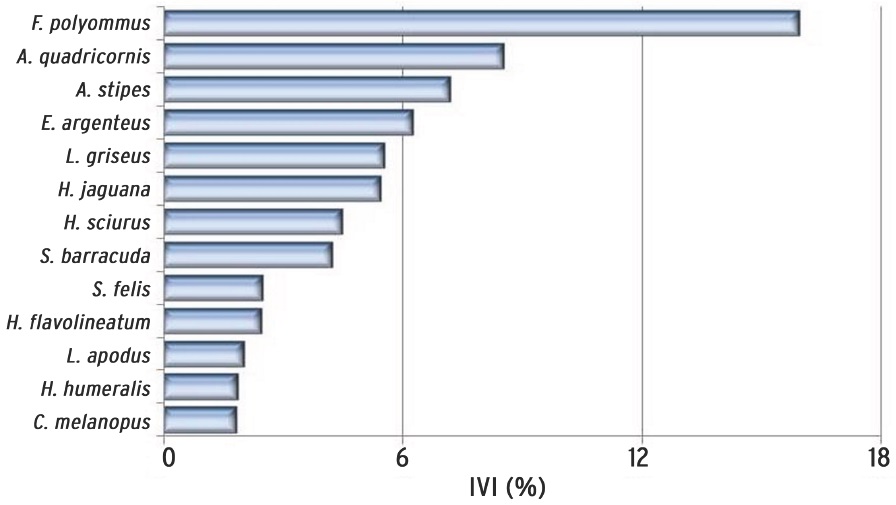
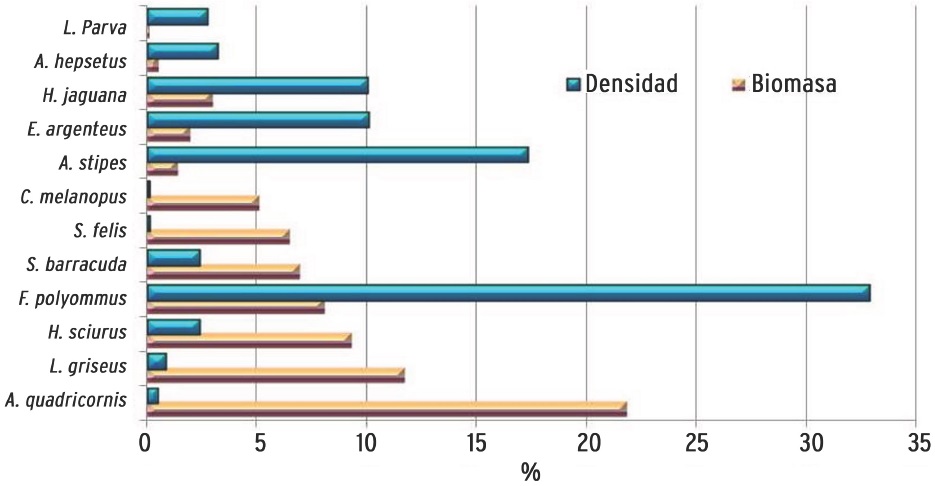
De acuerdo a la densidad, biomasa y frecuencia del total de las especies registradas (Tabla 3), 13 de ellas comprendieron el 67.91 % del IVI (Figura 5). Dentro de éstas sobresale el "bolín" Floridichthys polyommus por su abundancia y amplia distribución en los sistemas lagunares y norte de la zona costera. Las especies con mayor contribución por su biomasa, tienen poca representación numérica (A. quadricornis, Lutjanus griseusy Haemulon sciurus) y las especies abundantes numéricamente están representadas por individuos de talla pequeña (F. polyommus, Atherinomorus stipes, Lucania parva) y ejemplares en estadio juvenil, cuyas tallas (E argenteus. 1.5 cm a 7.5 cm de LE, H. jaguana. 2.5 cm a 10 cm de LE) y bajos valores de biomasa lo confirman (Figura 6).
Patrón de distribución espacial
Se registraron diferencias significativas de los ensamblajes de peces entre sitios (ANOSIM, R = 0.54, P < 0.001). La comparación de los parámetros ecológicos muestra los valores más altos de densidad y biomasa promedio en la zona costera (Cocopatos y Pescadores, respectivamente) (Tabla 4). En Cocopatos, la mayor densidad está dada por especies de engraulidos y gérridos, mientras que en Pescadores, las rayas (Urobatis jamaicencis), junto con los bagres (S. felis y C. melanopus), contribuyeron significativamente en los valores altos registrados para la biomasa. Los bagres, aunque presentaron baja abundancia númerica en este ecosistema, se consideran especies tipificantes por su aportación en biomasa (Figura 6). Por el contrario, el decremento de ambos parámetros ecológicos que se obtuvieron en Punta Sur, fue debido a la influencia de las corrientes que imperan en esa zona (observación in situ). La mayor riqueza y diversidad se obtuvieron en Pajarera Central y Garzas, con el registro de especímenes característicos de las cuatro categorías ecológicas. Los valores bajos de diversidad registrados en Puerto Viejo (Zona Lagunar) fue debido a la dominancia de A. stipes (26.8 %) y especies estuarinas como Fpolyommus (18.7 %) y Lucania parva (11.8 %), las cuales constituyeron especies tipificantes de este sitio lagunar (Tabla 4). Las especies de ciprinodontidos (Cyprinodon artifrons, F. polyommus), y gérridos (E. argenteus), caracterizaron tres o más sitios, siendo mayor o menor su contribución dependiendo de su ocurrencia. En este sentido, los ejemplares de F. polyommus presentaron alta incidencia en Puerto Viejo (Laguna y Playa), así como Cocopatos y Pajarera Norte. Por el contrario, Punta Sur constituyó un hábitat muy característico con especies tipificantes del ambiente arrecifal (100 %), como son las especies de la familia Scaridae reconocidas como pez loro o pez perico, debido a la cercanía de este complejo insular con el Corredor Arrecifal Mesoamericano.
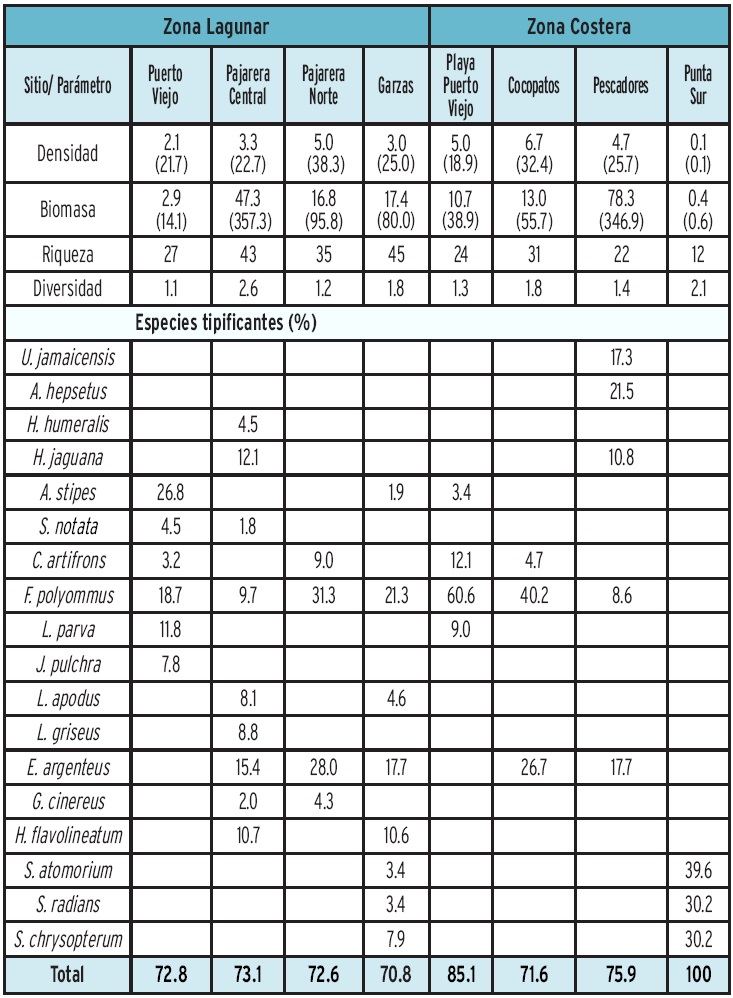
El dendrograma de afinidad (Figura 7), muestra la similitud de sitios en base a la composición y abundancia de las especies representativas (95 % de la abundancia total). Se generaron tres agrupamientos a un nivel de 40 %. El primer grupo correspondió a la localidad de Punta Sur, ubicada como su nombre lo dice en el sur de la isla, y la cual está influenciada por las corrientes del Golfo de México y el Caribe. En este grupo, además de que la densidad y riqueza específica es menor (Tabla 4), se registraron especies arrecifales de las familias Scaridae. En el grupo II se ubica la localidad Pescadores (Figura 7), con influencia marina y oleaje, caracterizado por especies marinas eurihalinas (U. jamaicensis, E. argenteus), estuarinas (F. polyommus), y la presencia de especies de movimiento rápido como las sardinas (H. jaguana), o por las que son transportadas por las corrientes (A. hepsetus) (Tabla 4). Por otra parte, en el grupo III se incluyen las zonas protegidas con poca influencia de corrientes. A su vez, dentro de este agrupamiento se diferencian tres subgrupos por la contribución en su abundancia de especies marinas eurihalinas y estuarinas: a) Cocopatos y Pajarera Norte, donde F. polyommus y E. argenteus son muy abundantes; b) Puerto Viejo y Playa Puerto Viejo, donde especies de la familia Cyprinodontidae (C. artifrons, F. polyommus) y Fundulidae (L. parva) son tipificantes y; c) Pajarera Central y Garzas, caracterizadas por una mayor riqueza de especies (Tabla 4).

Correlación de abundancia de especies con variables ambientales
Como resultado del análisis de correlación de las especies con los parámetros a través del coeficiente de Pearson (Ludwing y Reynolds, 1988), se determinaron 4 grupos de especies (Tabla 5): 1) Especies presentes en zonas de influencia marina con fuerte oleaje que habitan en aguas marinas, niveles altos de oxígeno disuelto y bajas concentraciones de nutrientes (correlaciones positivas con oxígeno). Dentro de este grupo se tienen Syngnathus floridae, Syngnathus louisianae 2) Especies que habitan en sistemas costeros y cuyo ambiente se caracteriza por presentar altas salinidades, bajas concentraciones de oxígeno disuelto y altas concentraciones de nutrientes (Cichlasoma urophthalmus, F. polyommus, Jordanella pulchra, Eucinostomus argenteus, Diapterus rhombeus, Megalops atlanticus, entre otras especies); 3) Aquellas con amplia distribución dentro de la reserva, encontrándose tanto en las lagunas, así como en zonas costeras con influencia de oleaje. Estas especies presentaron correlaciones positivas con la temperatura (C. artifrons) y amonio (Caranx latus, Achirus lineatus) y; 4) Especies que habitan el ambiente arrecifal y que se registraron en aguas con bajas concentraciones de nutrientes, principalmente con nitratos. Las correlaciones negativas con estos factores lo reflejan (Acanthostracion polygonius, Sparisoma radians, S. rubripinne, S. chrysopterum, S. viride).

DISCUSIÓN
Los sistemas lagunares costeros, manglares y humedales, son ampliamente reconocidos por sus valores de productividad primaria, llegando a ser 20 veces superior a la del mar y 5 veces a la de las zonas de surgencia (Flores-Verdugo y col., 2007). Estos ambientes proveen protección a los estadios juveniles de peces y crustáceos contra los depredadores, así como alimento que sostiene una alta abundancia y diversidad de peces y de otros invertebrados. Debido a su productividad y a constituir zonas de resguardo y protección, estos ecosistemas costeros son considerados "hábitat de crianza"; en parte porque exportan nutrientes esenciales -carbón, nitrógeno y fósforo-hacia las tramas alimenticias costeras. Esta exportación puede ocurrir a través de los procesos migratorios de los organismos hacia el ambiente marino, o por relaciones predador-presa, así como por procesos de mezcla de las aguas estuarinas, ricas en materia orgánica disuelta y particulada, con la zona costera (Beck y col., 2003).
La elevada producción heterotrófica de estos ecosistemas costeros también ha permitido que las regiones donde existen, sean un importante punto de alimentación y descanso de varios cientos de miles de aves acuáticas (Paul, 1991; Flores-Verdugo y col., 2007). Entre ellas existen muchas especies migratorias y otras que sin serlo, realizan desplazamientos oportunistas en búsqueda de humedales con abundancia de alimento y sitios para anidar (Weller, 1999). La anidación de estas aves está relacionada principalmente con la hidrología y la disponibilidad de alimento, pues de ello depende el éxito de forrajeo que determina la posibilidad de obtener los componentes alimentarios que satisfagan los requerimientos calóricos y nutricionales tanto de padres como de crías (Frederick y col., 2009). En este sentido, Isla Contoy es reconocida como Isla Pájaros, al ser una región con características particulares de productividad e idóneas para la anidación y alimentación de un alto número de especies de aves.
La importancia de este estudio radica en ser el "primero y único" que se ha realizado sobre la comunidad de peces y su distribución espacial a lo largo de la zona costera y sistemas lagunares del Parque Nacional Isla Contoy. La distribución de la ictiofauna en los sistemas lagunares está determinada por las respuestas físico-biológicas a su ambiente, lo que permite a los individuos seleccionar los hábitat que les ofrezcan la mejor combinación de alto crecimiento y reproducción y bajo riesgo de mortalidad. La distribución espacial de las especies refleja la respuesta de las especies o estadios de los ciclos de vida a distintos factores como la velocidad de la corriente, temperatura, salinidad, niveles de oxígeno disuelto y estructura de hábitat (Caballero-Vázquez y Vega-Cendejas, 2012). Esto es debido a los límites de tolerancia de las especies, dado que al ser la mayoría dependientes marino-eurihalinas (51 % de las especies) y marino-estenohalinas (41 % de las especie), su distribución va a estar restringida por los gradientes ambientales (Maci y Basset, 2009; Selleslagh y col., 2009). Dentro de éstas, la salinidad es la principal barrera física para la ocurrencia de los organismos (Vega-Cendejas y Hernández, 2004, Barletta y col., 2005; Whitfieldy col., 2006), ya sea entre hábitat, en un ciclo diario o a lo largo de un año. Otro factor importante que afecta la distribución de los peces es la preferencia del hábitat (Allen y col., 2007; Tse y col., 2008). Al respecto, muchos organismos, principalmente juveniles de varias especies, se encuentran en praderas de pastos sumergidos, planicies lodosas o entre las raíces de los manglares, sitios que les brindan refugio y protección contra predadores, además de proveer una alta abundancia de recursos alimenticios (por ejemplo: detritus, micro y macro-invertebrados).
En términos generales, en la mayoría de los sitios costeros (grupo III, Figura 7), se observó abundancia de medusas, lo que indica un exceso de materia orgánica. En particular, una de las especies que es un sifozoario (Cassiopeia xamachana), ha sido documentado como persistente en sistemas contaminados, y se le considera un bioindicador efectivo para estos ambientes costeros (Radwan y col., 2005; Templeton y Kingsford, 2009).
Individualmente, todas las variables hidrológicas tuvieron alguna relación significativa con la ocurrencia de algunas especies en particular. Los resultados obtenidos a través de la correlación de Pearson mostraron valores positivos significativos de la salinidad con la abundancia de especies marino eurihalinas (Syngnathus louisianae), estuarinas (J pulchra) y dulceacuícolas (C.urophthalmus) (Tabla 5). Estos valores indican los altos niveles de tolerancia de estas especies a la salinidad. Estas especies se registraron en las lagunas de Puerto Viejo y Pajarera Central, donde la salinidad alcanzó valores cercanos a 40 (Tabla 1). Al respecto, la salinidad ha sido reportada como un factor importante en la estructuración de ensamblajes de las especies de peces en sistemas lagunares-estuarinos (Selleslagh y Amara, 2008; Maci y Basset, 2009; Selleslagh y col., 2009).
Los bajos valores de nitrógeno registrados en todos los sistemas costeros de la Isla (Tabla 1), pueden deberse a su reducción debido a la salinidad y a su captación por las algas, como ha sido señalado por Herrera-Silveira y Ramírez (1998) para las lagunas de la Península de Yucatán. En los sistemas costeros, el contenido de fósforo en el agua está influenciado por la presencia del guano, debido a la cercanía de los sitios de anidación y preferencias tróficas de las aves. En la Bahía de Florida se ha determinado que el aporte de fósforo a la columna de agua es a través de las excretas de aves, las cuales actúan como fertilizante (Blaber, 1997), y son un indicador de procesos de eutrofización (Blais y col., 2005; Ratcha y col., 2010). Por el contrario, las estaciones ubicadas al sur de la isla (Punta Sur y Garzas), son aguas más frías y pobres de nutrientes debido a las corrientes que imperan en esa zona (Tabla 1).
La concentración de silicatos fue otra de las variables que explicaron significativamente (P < 0.05), la distribución y abundancia de las especies estuarinas y dulceacuícolas en las zonas costeras protegidas (Tabla 5). Diversos estudios han confirmado la estrecha relación entre aportes de agua subterránea y silicatos, y como un indicador de la vulnerabilidad de los ecosistemas al proceso de enriquecimiento de nutrimentos (Smith y col., 1999). En el Parque Nacional Isla Contoy se registraron afloramientos y descargas puntuales de agua dulce, principalmente en Playa Puerto Viejo y Lagunas Puerto Viejo y Pajarera Central. Esto se evidencia por las mayores concentraciones de silicatos en estos sitios y por el registro de especies dulceacuícolas y estuarinas (Tabla 4), las cuales muy probablemente son traídas a través de estas descargas, lo que revela una comunicación hidrológica y biológica entre la Península de Yucatán e Isla Contoy a través de 15 km aproximadamente, siendo éste el primer registro de descargas puntuales en el Parque Nacional. En las zonas costeras y lagunares del Parque Nacional Isla Contoy se registraron concentraciones medias de silicatos menores de < 10 μM (Tabla 1). Valor muy por debajo al reportado como referencia en las lagunas de tipo euhalino de Yucatán (20 μM) (Herrera-Silveira y col., 2004; Herrera-Silveira, 2006).
Respecto al oxígeno disuelto, se ha reportado que puede actuar como una barrera física en la distribución de las especies al combinarse con altas temperaturas (Marshall y Elliott, 1998). En el presente trabajo, el oxígeno disuelto en el agua se correlacionó de manera negativa con la abundancia de Megalops atlanticus, D. rhombeus y C. urophthalmus (Tabla 5), lo cual se debe a los hábitos bentónicos y altos niveles de tolerancia de estas especies a condiciones anóxicas. Las especies representativas de los sistemas costeros del Parque Nacional Isla Contoy son de origen marino, con capacidades eurihalinas y con dependencia estuarina a lo largo de su ciclo de vida, como es el caso de los pargos Lutjanusspp. y sábalo M. atlanticus (Figura 8). Estas especies también han sido reportadas como dominantes en estudios realizados en sistemas lagunares de la Península de Yucatán, como la Reserva Bocas de Dzilam (Arceo-Carranza y Vega-Cendejas, 2009), la Reserva de la Biósfera de Celestún (Vega-Cendejas y col., 1997; Vega-Cendejas, 2004; Poot-Salazar y col., 2009; Arceo-Carranza y col., 2010; Burgos-León y col., 2013) y de Ría Lagartos (Vega-Cendejas y Hernández, 2004; Peralta-Meixuero y Vega-Cendejas, 2011), y la laguna Chacmochuch (Caballero-Vásquez y Vega-Cendejas, 2012).

Floridichthys polyommus y C. artifrons son especies representativas de la Reserva de la Biósfera de Ría Lagartos. Sistema hiperhalino con valores de abundancia de 30l°/o para ambas especies (Vega-Cendejas y Hernández, 2004; Peralta-Mieixuero y Vega-Cendejas, 2011). Resultados similares han sido reportados en la Reserva de Bocas de Dzilam (Arceo-Carranza y Vega-Cendejas, 2009) y laguna de Chelem (Vega-Cendejas y Hernández, 2008). La capacidad fisiológica de estas especies para tolerar niveles de salinidad superiores a 100 y niveles de oxígeno < 3 mg / L, les confieren una amplia distribución y alta abundancia en hábitat hiperhalinos y eutróficos, al disminuir la competencia y disponibilidad de recursos alimenticios abundantes; condiciones estresantes para otros organismos (Vega-Cendejas y Hernández, 2004). Las otras especies registradas como dominantes en el Parque Nacional Isla Contoy, considerando el IVI, fueron Acanthostracion quadricornis (8.5 %), Atherinomorus stipes (7.2 %), la mojarra Eucinostomus argenteus (6.23 %) y la sardina Harengula jaguana (5.4 %) (Tabla 3, Figura 5). Estas especies marino eurihalinas y estenohalinas penetran a los sistemas costeros en estadio juvenil para alimentarse y protegerse, regresando posteriormente como adultos al ambiente marino (Hoese y Moore, 1998; Caballero-Vazquez y Vega-Cendejas, 2012).
CONCLUSIONES
El presente estudio contribuye al conocimiento de las poblaciones de peces que habitan los sistemas insulares mexicanos, al registrarse 92 especies de peces de origen marino eurihalino, marino estenohalino, estuarino y dulceacuícola, siendo éstos los primeros y únicos registros para la zona costera del Parque Nacional de Isla Contoy. Esta información es sustancial para la conservación de la biodiversidad de las áreas marinas protegidas y su valoración como hábitat críticos de diversos componentes bióticos; constituye un punto de partida para la comprensión ecológica y funcional dentro de las tramas tróficas, al ser los peces la principal fuente de alimento de las aves que se reproducen y anidan en estos sitios, y cuyas excretas son fuente de fósforo y nitrógeno, lo que incrementa la productividad a su vez de los sistemas costeros. A través del registro de silicatos y de la composición íctica con especies estuarinas residentes y dulceacuícolas, se confirma la presencia puntual de descargas subterráneas en las lagunas de Puerto Viejo, Pajarera Norte y Pajarera Central. Es imprescindible realizar otro estudio en el Parque Nacional, a fin de evaluar los cambios temporales en la estructura de los ensamblajes y valorar su función en la protección y conservación de la biodiversidad.

