











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkEl abuso de los psicoestimulantes como la cocaína (COC), la anfetamina (ANF) y sus derivados causan múltiples trastornos neuropsiquiátricos, incluyendo la adicción a las drogas. Por lo tanto, es importante estudiar la neurobiología de las conductas relacionadas con los psicoestimulantes. La COC y la ANF son agonistas monoaminérgicos indirectos que tienen afinidad por los transportadores de dopamina (DA), norepinefrina (NE) y serotonina (5-HT) involucrados en los sistemas de la recaptura y almacenamiento vesicular (Rothman y Baumann, 2003). La COC inhibe la recaptura de DA, NE y 5-HT; por lo tanto, aumenta los niveles sinápticos de esos neurotransmisores. La ANF entra en las terminales presinápticas y, una vez dentro, bloquea la recaptura de DA, NE y 5-HT en las vesículas sinápticas, por lo que promueve un aumento en las concentraciones citoplasmáticas de esas monoaminas. Subsecuentemente, conforme los niveles citoplasmáticos de esas monoaminas aumentan, las monoaminas salen de la neurona, vía la inversión de la función de los transportadores de la recaptura de monoaminas, lo que conduce también a un aumento en los niveles sinápticos de DA, NE y 5-HT (Amara y Sonders, 1998; Elliot y Beveridge, 2005; Kahlig y Galli, 2003; Rothman y Baumann, 2003).
El sistema mesolímbico de la DA, en particular la proyección del área tegmental ventral (ATV) al núcleo accumbens (NAcc), es un locus importante en la producción de los efectos locomotores, reforzantes, gratificantes y discriminativos de los psicoestimulantes como la COC y la ANF (Di Chiara, 1995; Filip y Cunningham, 2002; Koob, 1992; Pontieri, Tanda y Di Chiara, 1995). La administración de COC y de ANF aumenta rápidamente la neurotransmisión dopaminérgica al interferir con la función del transportador de la DA, como se describió previamente, y como consecuencia se produce un aumento en la señalización dopaminérgica en áreas límbicas (Koob, 1992; Koob, Sanna y Bloom, 1998).
Actualmente hay evidencia que sugiere que la neurotransmisión de la 5-HT también juega un papel importante en algunos efectos conductuales relacionados con la adicción a los psicoestimulantes (Fillip, 2005; Walsh y Cunningham, 1997). Aunque hay 7 familias de receptores de 5-HT (5-HT1 - 5-HT7) con al menos 16 subtipos que se han identificado (Hannon y Hoyer, 2008), varios estudios han sugerido que los receptores 5-HT1A podrían estar involucrados en los efectos conductuales de las drogas de abuso, debido a que estos receptores están localizados pre y post sinápticamente en regiones límbicas, incluyendo el NAcc y el ATV (Chalmers y Watson, 1991; Müller, Carey, Huston y de Souza, 2007). Estudios previos realizados en nuestro laboratorio (Miranda, Sandoval-Sánchez, Cedillo, Jiménez y Velázquez-Martínez, 2010) mostraron que una dosis baja (0.03 mg/kg) del agonista a receptores 5-HT1A, 8-OH-DPAT, atenuó las propiedades discriminativas de la ANF. Este efecto podría deberse a varias causas. Primero, el control de la actividad de las neuronas dopaminérgicas del ATV está regulado por las interneuronas GABAérgicas del ATV a través de los receptores GABAA localizados en los cuerpos celulares de las neuronas dopaminérgicas del ATV (Creed, Ntamati y Tan, 2014). Segundo, las neuronas serotoninérgicas provenientes del núcleo del rafe inervan al sistema de la recompensa, particularmente al ATV y al NAcc, lo cual modula la actividad del sistema mesolímbico de la DA a través de la activación de varios receptores, entre ellos los receptores 5-HT1B, localizados en las terminales presinápticas de las interneuronas GABAérgicas del ATV (Bruinvels et al., 1994). La activación de estos receptores produce un efecto inhibitorio sobre la liberación de GABA, desinhibiendo a las neuronas dopaminérgicas del ATV (Yan, Zheng, Feng y Yan, 2005). De esta forma, la administración del agonista 8-OH-DPAT, que activa los receptores somatodendríticos del núcleo del rafe (Carey et al., 2005b), disminuye la actividad de las neuronas serotoninérgicas que inervan las interneuronas GABAérgicas del ATV (Hervé, Pickel, Joh y Beaudet, 1987; Van Bockstaele, Cestari y Pickel, 1994) y, como consecuencia, disminuye la actividad de los receptores 5-HT1B localizados en las terminales presinápticas de las interneuronas GABAérgicas (Yan, Zheng y Yan, 2004), incrementando los niveles de GABA en ATV y disminuyendo los niveles de DA en NAcc (O’Dell y Parsons, 2004; Yan y Yan, 2001). Si esta sugerencia es cierta, entonces el efecto observado en el estudio previo podría prevenirse si se bloquean los receptores GABAA localizados en el soma de las neuronas dopaminérgicas del ATV mediante la administración de un antagonista selectivo de los receptores GABAA. El presente estudio se diseñó para examinar los efectos producidos por la administración sistémica de un antagonista GABAA, la bicuculina, sobre los efectos del 8-OH-DPAT en las propiedades discriminativas de la ANF utilizando el condicionamiento de aversión a los sabores (CAS).
Los procedimientos de discriminación de drogas proporcionan información importante sobre los mecanismos neurofarmacológicos que subyacen a los efectos subjetivos de las drogas de abuso. En estos procedimientos se entrena a los animales de investigación a discriminar la droga de entrenamiento de un placebo, generalmente el vehículo de la droga de entrenamiento. Una vez que se aprendió la discriminación droga-vehículo, se valoran los efectos de distintos compuestos.
Método
Animales
Se utilizaron 10 ratas macho de la cepa Wistar de aproximadamente 120 días de edad y con un peso de 200 a 250 g provenientes del Bioterio General de la Facultad de Estudios Superiores Iztacala-UNAM. Las ratas se alojaron individualmente en cajas-hogar de acero inoxidable de 30 × 20 × 20 cm, con acceso libre a la comida (Teklad LM485 Rat Diet by Harlan) y bajo un ciclo luz-oscuridad controlado (luz: 8:00 am-8:00 pm) y a una temperatura ambiente de 21 ± 1 °C. Las ratas tuvieron acceso a soluciones líquidas a través de una o 2 botellas graduadas invertidas colocadas en el panel frontal de la caja. Los procedimientos de cuidado y manejo de animales se realizaron conforme a la Norma Oficial Mexicana (NOM-062-00-1999), titulada «Especificaciones técnicas para la producción, cuidado y uso de animales de laboratorio» y aprobada por el comité local de Bioética.
Drogas
Las drogas utilizadas en este estudio fueron sulfato de D-anfetamina (Sigma-Aldrich, St, Louis, MO, EE. UU.), cloruro de litio (LiCl; Sigma-Aldrich, St, Louis, MO, EE. UU.), hidrobromuro de 8-hidroxi-2-(di-n-propilamino)-tetralina (8-OH-DPAT) y bicuculina (Tocris, Ballwin, MO, EE. UU.). Todas las dosis de las drogas se calcularon de acuerdo al peso de la sal, disueltas en una solución salina (0.9%), y se administraron intraperitonealmente en un volumen de 1.0 ml/kg, a excepción del LiCl, que fue administrado en una dosis de 0.34 mEq (2 ml/kg de una solución de 0.177 M). La sacarina (Elly Lilly, México) fue disuelta en agua destilada a una concentración de 0.15% (p/v). Todas las drogas y la solución de sacarina fueron preparadas diariamente.
Procedimiento
El entrenamiento de discriminación con CAS y los procedimientos de prueba fueron similares a los descritos con anterioridad (Miranda et al., 2009). En resumen, las ratas fueron privadas por 23.5 h y entrenadas por 7 días a tomar agua diariamente durante un periodo de 20 min. Posteriormente, durante 2 días fueron entrenadas a tomar la solución de sacarina durante 10 min. Para la adquisición de la discriminación con el condicionamiento de aversión al sabor las ratas fueron sometidas a 2 tipos de ensayos: ensayo droga y ensayo salina (ver Tabla 1 para detalles del procedimiento).
Tabla 1 Diseño experimental del entrenamiento de discriminación de drogas
| Ensayo | Procedimiento | Pruebas de sustitución y combinación | |
|---|---|---|---|
| Compuestos | Prueba de elección | ||
| Droga | Anfetamina-sacarina-LiCL | Anfetamina (0.1, 0.3, 1.0) | Agua-sacarina |
| Salina | Salina sacarina-salina | 8-OH-DPAT (0.003, 0.01, 0.03) | Agua-sacarina |
| Bicuculina (0.5, 1.0, 3.0) | Agua-sacarina | ||
| 8-OH-DPAT (0.03) + ANF (1.0) | Agua-sacarina | ||
| Bicuculina (3.0) + 8-OH-DPAT (0.03) + ANF (1.0) | Agua-sacarina | ||
El grupo de ratas (n = 10) se sometió al entrenamiento de la discriminación ANF-salina. Posteriormente se sometieron a las pruebas de sustitución y combinación. Antes de cada prueba se llevaron a cabo ensayos de reentrenamiento y evaluación de generalización con la dosis de entrenamiento de la ANF y salina para comparar estos resultados con los obtenidos con los compuestos de prueba. Entre paréntesis las dosis, expresadas en mg/kg; intraperitonealmente.
Ensayos droga. A las ratas se les administró 1.0 mg/kg de ANF y 30 min después se les permitió el acceso a la sacarina durante 10 min. Después de finalizar este período, se les administró 2.0 ml/kg de 0.177 M de LiCl.
Ensayos salina. A las ratas se les administró salina, y 30 min después se les permitió el acceso a la sacarina durante 10 min. Después de finalizar este período, se les administró 1.0 ml/kg de salina.
Entre los ensayos droga y los ensayos salina hubo 2 días de descanso, donde se les permitió el acceso al agua simple durante 30 min en las cajas-hogar. El ciclo ensayo droga-ensayo salina se repitió 8 ocasiones en un orden aleatorio, con la única restricción de que no tuvieran lugar más de 2 ensayos droga consecutivos.
Pruebas de generalización y combinación con anfetamina y ligandos a receptores GABA A y 5-HT 1A
Estas pruebas comenzaron 2 días después de finalizar la adquisición de la discriminación ANF-salina y se hicieron sobre un ciclo de 4 días. En el día 1, a las ratas se les sometió a un procedimiento similar al que se sometieron en el ensayo droga. En el día 2, a los sujetos se les permitió consumir agua simple durante 30 min en sus cajas-hogar. El día 3 fue igual al ensayo salina. En el día 4, a las ratas se les administró una dosis particular de ANF, una dosis de una droga de prueba o una combinación de 2 o 3 de las drogas utilizadas en este experimento (Tabla 1). Después, las ratas se sometieron a una prueba de 2 botellas por 10 min; una botella tenía agua simple y la otra una solución de sacarina. Al final de la prueba no se administró ni LiCl ni salina. Las dosis y los tiempos entre la administración de cada droga y la prueba de 2 botellas se seleccionaron de acuerdo a la literatura. La secuencia de las drogas y sus dosis se eligió al azar, y el ciclo de 4 días se repitió hasta que todas las dosis de las drogas de prueba se evaluaron. Las drogas y dosis que se evaluaron en las pruebas de sustitución fueron ANF (0.1, 0.3 y 1.0 mg/kg, 30 min), 8-OH-DPAT (0.003, 0.01 y 0.03 mg/kg, 30 min) y bicuculina (0.5, 1.0 y 3.0 mg/kg, 30 min). En el caso de las pruebas de combinación se administró una dosis de 8-OH-DPAT (0.03 mg/kg), 8-OH-DPAT (0.03 mg/kg) + ANF (1.0 mg/kg) y bicuculina (3.0 mg/kg) + 8-OH-DPAT (0.03 mg/kg) más una dosis de ANF (1.0 mg/kg). En el caso de las pruebas de sustitución, también se evaluó la dosis de entrenamiento de la ANF (1.0 mg/kg) y la administración de salina antes de iniciar la evaluación de cada droga.
Análisis estadístico
En el curso de la adquisición de la discriminación ANF-salina se registró el consumo de la solución de sacarina en los ensayos droga y los ensayos salina y se analizó con un ANOVA de 2 factores, con los tipos de ensayos (ensayo droga - ensayo salina) como primer factor y el número de ensayo (los últimos 3 ensayos de cada condición) como segundo factor. Se registró también el consumo de líquidos en las pruebas de generalización y sustitución. A partir de estos datos se calculó un índice de aversión a la sacarina con la fórmula A/A + B, donde A es el consumo de sacarina y B es el consumo de agua. Con este índice, un valor de 1.0 indica preferencia por la sacarina y un valor de 0.0 aversión por la sacarina. Estos datos se analizaron usando un ANOVA de una vía; cuando los ANOVA fueron significativos, se llevó a cabo un análisis de comparaciones posteriores con la prueba de Tukey. En todas las pruebas, el nivel de rechazo del error tipo I fue de 0.05.
Resultados
Adquisición de la discriminación anfetamina-salina
Las ratas aprendieron la discriminación ANF-salina (Fig. 1). No se encontraron diferencias significativas (F [2,27] = 0.494, p > 0.05) entre el consumo de sacarina en las sesiones de la línea base, el primer ensayo droga y el primer ensayo salina. Durante los ensayos en los que se administró la ANF seguido por los apareamientos sacarina-LiCl se produjo una reducción del consumo de sacarina. Un ANOVA de 2 vías reveló diferencias significativas entre los últimos 3 ensayos droga y los últimos 3 ensayos salina (condición de tratamiento, F [1,9] = 49.63, p < 0.05). Los efectos del número de ensayo (F [2,18] = 0.174, p > 0.05) y la interacción entre los factores número de ensayo y tipo de ensayo (F [2,18] = 0.884, p > 0.05) no fueron significativos.

Pruebas de generalización con anfetamina
La administración de diferentes dosis de ANF que se evaluaron durante las pruebas de generalización con dos botellas mostró un control de estímulos dependiente de la dosis (panel izquierdo de la Figura 2). La evaluación de 1.0 mg/kg de ANF produjo una señal discriminativa similar a la producida con la dosis de entrenamiento de la ANF (1.0 mg/kg). Un ANOVA simple reveló diferencias significativas en la condición de tratamiento (F [4,45] = 21.214, p < 0.05). La prueba post hoc de Tukey reveló que el tratamiento con salina, las dosis de 0.1 y 0.3 mg/kg de ANF, produjo diferencias significativas con respecto a la dosis de entrenamiento de ANF.
Pruebas de generalización con el agonista a receptores 5-HT 1A , 8-OH-DPAT
Se evaluaron diferentes dosis del agonista a receptores 5-HT1A 8-OH-DPAT (0.003, 0.01 y 1.0 mg/kg). Los resultados de estas pruebas mostraron que ninguna dosis del 8-OH-DPAT sustituyó la señal discriminativa de la ANF (panel central de la Fig. 2). Un ANOVA de una vía reveló diferencias significativas en la condición de tratamiento (F [4,36] = 5.039, p < 0.05). Las comparaciones posteriores con la prueba de Tukey revelaron que la preferencia por la sacarina en todas las dosis del agonista 8-OH-DPAT fue significativamente diferente a la observada con la dosis de entrenamiento de ANF.

Figura 2 Resultados de las pruebas de sustitución con diferentes dosis de ANF (0.1, 0.3 y 1.0mg/kg), 8-OH-DPAT (0.003, 0.01 y 0.03mg/kg) y bicuculina (0.5, 1.0 y 3.0mg/kg). Se muestra la preferencia por la sacarina (0.0 indica una aversión a la sacarina y 1.0 indica una preferencia por la sacarina). * indica diferencias significativas (Tukey, p<0.05) con la dosis de entrenamiento de ANF. La gráfica del panel inferior indica el consumo total de líquidos.
Pruebas de generalización con el antagonista a receptores GABA A , bicuculina
Se evaluaron diferentes dosis del antagonista bicuculina (0.5, 1.0 y 3.0 mg/kg). Los resultados de estas pruebas mostraron que ninguna dosis de este compuesto sustituyó la señal discriminativa de la ANF (panel derecho de la Fig. 2). Una ANOVA de una vía reveló diferencias significativas en la condición de tratamiento (F [4,45] = 38.539, p < 0.05). Las pruebas posteriores de Tukey revelaron que la preferencia por la sacarina en todas las dosis de bicuculina fue significativamente diferente a la observada con la dosis de entrenamiento de ANF.
Pruebas de combinación con el agonista 8-OH-DPAT más anfetamina y el antagonista a receptores GABA A bicuculina más 8-OH-DPAT más anfetamina
Los resultados de las pruebas de combinación donde se evaluó la administración conjunta de bicuculina (3.0 mg/kg) más 8-OH-DPAT (0.03 mg/kg) más ANF (1.0 mg/kg) se muestran en la Figura 3. Como se puede observar, la administración de 8-OH-DPAT (0.03 mg/kg) + ANF (1.0 mg/kg) produjo una disminución en las propiedades discriminativas de la ANF, y la administración de bicuculina (3.0 mg/kg) previno el efecto del 8-OH-DPAT sobre la señal discriminativa de la ANF. Un ANOVA de una vía reveló diferencias significativas en el tratamiento (F [2,29] = 11.933, p < 0.05); el análisis post hoc con la prueba de Tukey reveló diferencias significativas entre la administración del 8-OH-DPAT (0.3 mg/kg) + ANF (1.0 mg/kg), con respecto a la dosis de entrenamiento de ANF.

Figura 3 Resultados de las pruebas de combinación con una dosis de 8-OH-DPAT (0.03mg/kg) más una dosis fija de ANF (1.0mg/kg) y de la administración de una dosis de bicuculina (3.0mg/kg) más una dosis de 8-OH-DPAT (0.03mg/kg) más una dosis de ANF (1.0mg/kg). Las barras indican la media±error estándar de la media. Se muestra la preferencia por la sacarina (0.0 indica una fuerte aversión a la sacarina y 1.0 indica una preferencia por la sacarina). * indica diferencias significativas (Tukey, p<0.05) con la dosis de entrenamiento de la ANF. La gráfica del panel inferior indica el consumo total de líquidos.
Consumo de líquidos en las pruebas de generalización y combinación
El consumo total de líquidos (Fig. 2, panel inferior) no se afectó por la administración de ANF (F [4,45] = 2.853, p > = 0.05), 8-OH-DPAT (F [4,45] = 1.269, p > 0.05) o bicuculina (F [4,45] = 0.980, p > 0.05). En el caso de las pruebas de combinación, el consumo total de líquidos (Fig. 3, panel inferior) tampoco se afectó por la administración 8-OH-DPAT (1.0 mg/kg), 8-OH-DPAT (0.03 mg/kg) + ANF (1.0 mg/kg) o bicuculina (3.0 mg/kg) + 8-OH-DPAT(0.03 mg/kg) + ANF(1.0 mg/kg) (F [2,29] = 0.644, p > 0.05).
Discusión
El propósito de la presente investigación fue examinar el efecto del bloqueo de los receptores GABAA y la activación de los receptores 5-HT1A en la señal discriminativa de la ANF utilizando el CAS como procedimiento de discriminación de drogas. Los resultados obtenidos muestran, en primer lugar, que las ratas fueron capaces de discriminar ANF de salina después de 8 ensayos droga y 8 ensayos salina. Estos resultados son consistentes con los obtenidos en estudios previos donde se utilizó la ANF como droga de entrenamiento en un procedimiento de CAS (Herrera y Velázquez-Martínez, 1997; Miranda et al., 2007; Miranda et al., 2009).
Adicionalmente, se encontró una sustitución dependiente de la dosis en las pruebas de generalización con ANF, mientras que la administración de bicuculina, antagonista de los receptores GABAA, así como el agonista de los receptores 5-HT1A, 8-OH-DPAT, fracasaron en sustituir la señal discriminativa de la ANF.
En segundo lugar, la administración de 8-OH-DPAT previo a una dosis de ANF produjo una atenuación de la señal discriminativa de la ANF. Estos resultados sugieren que los receptores 5-HT1A juegan un papel importante en la modulación de la señal discriminativa de la ANF y son compatibles con estudios donde se ha reportado que los efectos del 8-OH-DPAT, con dosis que estimulan preferencialmente a los autorreceptores somatodendríticos 5-HT1A, bloquean los efectos estimulantes de la COC en el aparato locomotor (Carey, DePalma y Damianopoulos, 2005a), las propiedades discriminativas de la metanfetamina (Sasaki, Tatham y Barrett, 1995) y también reducen la expresión de la sensibilización locomotora inducida por ANF (Przegalinski, Siwanawicz, Baran y Filip, 2000). En otros estudios se ha reportado que el 8-OH-DPAT inhibe la hiperactividad inducida por la COC (Przegalinski y Filip, 1997). Los resultados anteriores apoyan la hipótesis de que los efectos de las dosis bajas (en el rango de 0.003 a 0.03 mg/kg) del agonista de los receptores 5-HT1A 8-OH-DPAT modulan las conductas relacionadas con la adicción a los psicoestimulantes, y estos efectos parecen estar mediados por la estimulación de los autorreceptores 5-HT1A somatodendríticos del núcleo del rafe (Lê et al., 2005; Fletcher, Cliffe y Dourish, 1993; Fletcher, 1995).
En contraste con los resultados previos, otros estudios han reportado que la administración de 0.1 mg/kg 8-OH-DPAT incrementa la actividad locomotora (Tricklebank, Middlemiss y Neill, 1986; Dourish, Hutson y Curzon, 1985; Lucki, Ward y Frazer, 1989; Jackson, Wallsten, Jerning, Hu y Deveney, 1998; Müller et al., 2007). Young, Bondareva, Wesolowska y Glennon (2006) realizaron un estudio de discriminación de drogas donde los resultados mostraron que no hubo una generalización cuando se administró el 8-OH-DPAT (0.01-0.5 mg/kg) ni antagonizó los efectos de las propiedades discriminativas de la ANF. Estos resultados son consistentes con estudios donde la administración de 8-OH-DPAT (0.125-1.6 mg/kg), buspirona (2.0-16 mg/kg) o gepirona (2.5-10.0 mg/kg) no sustituyeron la señal discriminativa de la COC en ratas (Callahan y Cunningham, 1995, 1997; Przegalinski y Filip, 1997; Rapoza, 1993).
La razón de esta discrepancia podría involucrar las dosis empleadas en los estudios. Se ha reportado que las dosis bajas (en el rango de 0.003-0.03 mg/kg) dan lugar a la activación preferencial de los autorreceptores somatodendríticos 5-HT1A del núcleo del rafe (Casanovas, Lesourd y Artigas, 1997), mientras que las dosis altas (en el rango de 0.2-1.6 mg/kg) activan tanto a los autorreceptores como a receptores postsinápticos (Müller et al., 2007).
Estos datos son compatibles con estudios previos en donde la inhibición directa de neuronas serotoninérgicas del núcleo del rafe con la microinyección de 8-OH-DPAT (1.0 y 2.0 μg) produjo un decremento de aproximadamente el 25% de DA en el NAcc (Yoshimoto y McBride, 1992). En otros estudios de microdiálisis, la administración sistémica de 8-OH-DPAT (25, 50 y 100 μg/kg s.c.) previo a la administración de ANF (1.0 mg/kg s.c.) inhibió el incremento de DA extracelular en el NAcc, y este efecto fue prevenido por la administración del antagonista WAY100635 (Ichikawa, Kuroki, Kitchen y Meltzer, 1995; Ichikawa y Meltzer, 2000), mientras que una dosis alta de R(+)-8-OH-DPAT no afectó las concentraciones basales de DA en el NAcc (Ichikawa y Meltzer, 1999).
El mecanismo que podría explicar por qué en la presente investigación se observa una disminución de los efectos discriminativos de la ANF después de la administración del 8-OH-DPAT es el siguiente: debido a que el sistema de la recompensa del cerebro recibe proyecciones serotoninérgicas del núcleo del rafe, una disminución o un aumento en la liberación de 5-HT podría modificar la concentración de DA en el NAcc y alterar los efectos conductuales de las drogas de abuso. Las neuronas serotoninérgicas provenientes del núcleo del rafe inervan el ATV y el NAcc (Broderick y Phelix, 1997). El control de las neuronas dopaminérgicas del ATV está regulado por diferentes receptores, entre ellos los receptores 5-HT1B, que se encuentran localizados en las terminales de las interneuronas GABAérgicas del ATV. La estimulación de los receptores 5-HT1B tiene un efecto inhibitorio en la liberación de GABA, y por lo tanto en la desinhibición de las neuronas dopaminérgicas del ATV (Fig. 4). Los resultados de esta investigación apoyan la hipótesis de que los receptores 5-HT1A están involucrados en los efectos conductuales de las drogas de abuso, debido a que se ha reportado que la administración de dosis bajas (0.03 mg/kg) de 8-OH-DPAT disminuye la actividad de las neuronas serotoninérgicas que proyectan a las interneuronas GABAérgicas del ATV (Hervé et al., 1987; Van Bockstaele et al., 1994), disminuyendo la actividad de los receptores 5-HT1B de las interneuronas GABAérgicas (Yan et al., 2004), y como consecuencia se ha observado un incremento en los niveles extracelulares de GABA en ATV y una disminución en los niveles de DA en NAcc (O’Dell y Parsons, 2004; Yan et al., 2005; Yan et al., 2004).
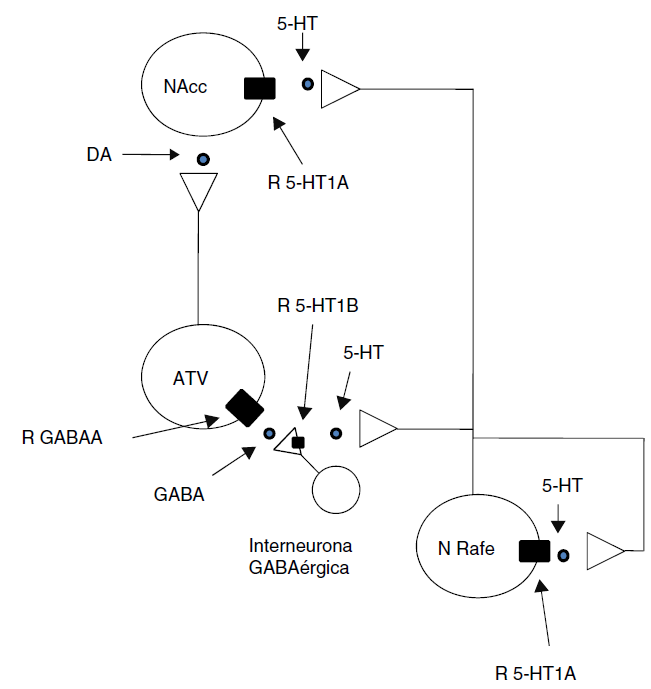
Figura 4 Se muestra cómo la bicuculina podría bloquear los efectos del agonista 5-HT1A 8-OH-DPAT sobre las propiedades discriminativas de la ANF (ver discusión para detalles)ATV: área tegmental ventral; DA: dopamina; 5-HT; serotonina; N: núcleo; NAcc: núcleo accumbens; R: receptor.
En tercer lugar, en la presente investigación también se observó que la administración del antagonista GABAA bicuculina, previa a la administración del agonista 8-OH-DPAT más una dosis de ANF, previno el efecto del 8-OH-DPAT sobre la señal discriminativa de la ANF. El mecanismo que pudiera explicar estos efectos es similar al señalado en el párrafo anterior. El apoyo de esta sugerencia proviene de estudios que reportan que la actividad de las neuronas dopaminérgicas del ATV es inhibida por las interneuronas GABAérgicas de ATV (Johnson y North, 1992), y como consecuencia de esta inhibición, los procesos de gratificación o recompensa son alterados (Van Zessen, Phillips, Budygin y Stuber, 2012). Los efectos inhibitorios de las interneuronas GABAérgicas sobre las neuronas dopaminérgicas de ATV son mediadas por los receptores GABAA localizados en los cuerpos celulares de esas neuronas (Westerink, Kwint y DeVries, 1996). Adicionalmente, se ha reportado que el bloqueo farmacológico con bicuculina de los receptores GABAA aumenta los estallidos de disparo de las neuronas dopaminérgicas (Paladini y Tepper, 1999) y aumenta los niveles extracelulares de DA en el NAcc (Ikemoto, Kohl y McBride, 1997), sugiriendo que las interneuronas GABAérgicas de ATV controlan la actividad de las neuronas dopaminérgicas de ATV a través de los receptores GABAA.
También se ha reportado que la neurotransmisión serotoninérgica regula la actividad de las interneuronas GABAérgicas de ATV (Johnson y North, 1992), e indirectamente también modula la actividad de las neuronas dopaminérgicas de ATV (Yan et al., 2004). Esta modulación involucra a varios tipos de receptores serotoninérgicos, entre ellos los receptores 5-HT1B (Yan et al., 2004). Estos receptores se encuentran localizados con moderada densidad como heterorreceptores en las terminales de las interneuronas GABAérgicas de ATV que hacen contacto sináptico con neuronas dopaminérgicas (Bruinvels et al., 1994). Debido a que los receptores 5-HT1B están acoplados negativamente a la adenilato ciclasa, el aumento o disminución de la neurotransmisión serotoninérgica disminuye o aumenta, respectivamente, la liberación de GABA en ATV (Yan y Yan, 2001). En este sentido, el bloqueo de los receptores GABAA en los cuerpos celulares de las neuronas dopaminérgicas podría prevenir la influencia indirecta de la 5-HT sobre el circuito de la gratificación (Ikemoto et al., 1997). En línea con lo anterior, en la presente investigación el bloqueo de los receptores GABAA por la bicuculina previno los efectos de la administración del 8-OH-DPAT sobre la señal discriminativa de la ANF.
En resumen, la presente investigación mostró que la administración del antagonista GABAA bicuculina previno los efectos del agonista 5-HT1A, 8-OH-DPAT sobre la señal discriminativa de la ANF. Estos datos proporcionan evidencia de que las conductas relacionadas con la adicción a los psicoestimulantes como la ANF involucran diferentes sistemas de neurotransmisión, como la DA, la 5-HT y el GABA.

