










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa de investigación psicológica
versión On-line ISSN 2007-4719versión impresa ISSN 2007-4832
Acta de investigación psicol vol.2 no.1 Ciudad de México abr. 2012
Efecto de la portación de alelos de riesgo de MAO-A y DRD4 sobre mediciones de agresión
Effect of Risk Alleles MAO-A and DRD4 on Aggression Measures
César Romero, Feggy Ostrosky1 & Beatriz Camarena2
Universidad Nacional Autónoma de México.
2Instituto Nacional de Psiquiatría "Ramón de la Fuente Muñiz".
1 Correspondencia:
Facultad de Psicología,
Universidad Nacional Autónoma de México,
Rivera de Cupia 11071, Lomas de Reforma,
México D.F. 11900. Fax: (+525) 5251-76-56.
Correo: feggy@servidor.unam.mx
Original recibido / Original received: 16/12/2011
Aceptado / Accepted : 31/03/2012
Resumen
Entre los diversos factores asociados con la predisposición a la conducta violenta se encuentra la portación de alelos de riesgo. Para investigar el efecto de dos alelos de riesgo (baja actividad de MAO-A y 7 repeticiones DRD4) sobre mediciones de agresión, se evaluaron mediante escalas psicológicas a 60 hombres sanos. Los resultados indicaron que tanto el efecto principal de cada uno de los alelos de riesgo, como su interacción impactan sobre mediciones de hostilidad, enojo, impulsividad, empatía y rasgos antisociales de psicopatía. Se concluye que es necesario investigar el efecto de estos alelos sobre la estructura y función cerebrales, además estos alelos aparentemente confieren riesgo para el desarrollo de conductas violentas.
Palabras clave: Agresión, DRD4, Epistasis, Impulsividad, MAO-A, Violencia.
Abstract
Among the factors associated with the predisposition to violent behavior is the bearing of risk alleles. To investigate the effect of two risk alleles (low activity of MAO-A and 7 repeat DRD4) 60 healthy men were assessed on psychological scales of aggression. The results indicated that both the main effect of each of the risk alleles, and their interaction impact on measures of hostility, anger, impulsivity, empathy and antisocial traits of psychopathy. We conclude that it is necessary to investigate the effect of these alleles on brain structure and function; these alleles apparently also confer risk for the development of violent behavior.
Keywords: Aggression, DRD4, Epistasis, Impulsivity, MAO-A, Violence.
Actualmente ha habido avances importantes en el campo de la genética de la agresión y la violencia, se sabe que la contribución genética a la conducta es importante, se ha propuesto que los factores genéticos explican entre el 40% y el 50% de la varianza de los rasgos asociados al trastorno antisocial incluyendo la violencia (Rhee & Waldman, 2002).
Epidemiológicamente se han asociado polimorfismos de riesgo para el desarrollo de conductas violentas en el gen de la monoamin oxidasa-A (MAO-A) y en el gen de los receptores de dopamina D4 (DRD4).
La MAO es una enzima que cataliza monoaminas en el cerebro y en tejidos periféricos. Se conocen dos tipos de MAO que se han denominado MAO-A y MAO-B. La forma A tiene afinidad por la serotonina (5HT), norepinefrina (NE), dopamina (DA) y la clorgilina, la forma B tiene mayor afinidad por la feniletilamina, benzilamina y el deprenil, ambas formas están localizadas en el cerebro en la parte externa de la membrana de la mitocondria. Los genes de ambas formas de MAO se cree que provienen del mismo gen ancestral, ambos están localizados en el cromosoma X (Xp11.23) y cuentan con 15 exones con idéntica organización intrón-exón. En humanos y roedores MAO-A está presente antes que MAO-B, los niveles de MAO-A son mayores en la etapa prenatal y posnatal que en la adultez, mientras que los niveles de MAO-B incrementan con la edad, este incremento de MAO-B se debe a la proliferación de células gliales donde se localiza predominantemente (Shih, Chen & Ridd, 1999). La preferencia que tiene MAO-A por 5HT y NE se descubrió en ratones knockout para MAO-A, y no para MAO-B, los cuales presentaron mayores niveles de serotonina y norepinefrina en el cerebro comparados con ratones control, además presentaron mayores niveles de conducta agresiva (Cases et al., 1995).
En humanos se ha descrito una mutación en el gen de la MAO-A (Xp11.23-11.4) que introduce un codón de terminación e identificado en una familia con predisposición a la violencia de tipo impulsivo (Brunner, Nelen, Breakefield, Ropers & van Oost, 1993). Dicha mutación es rara, sin embargo se ha localizado otro polimorfismo funcional del tipo VNTR o número variable de repeticiones en tándem, localizado en la región promotora del gen de la MAO-A (MAO-A-uVNTR) y caracterizado por una repetición de la secuencia de 30 pares de bases que impacta en la eficiencia de la transcripción del gen in Vitro; la actividad transcripcional mostró ser mayor en los portadores de los alelos de 3.5 o 4 repeticiones (alelos MAOA-H) y menor en los portadores de 2, 3 o 5 repeticiones (alelos MAOA-L) (Sabol, Hu & Hamer, 1998).
Los estudios llevados a cabo en humanos acerca del efecto de la variación genética de MAO-A sobre mediciones de agresión impulsiva han arrojado resultados contradictorios. Los resultados que destacan son los que se refieren al papel moderador del genotipo MAO-A en la relación abuso infantil-violencia. Caspi et al. (2002) reportaron una interacción significativa genXambiente que mostró que los sujetos que recibieron maltrato infantil y eran portadores de la variante MAOA-L tenían una mayor probabilidad de desarrollar conducta antisocial en la edad adulta, siendo más fuerte el efecto en los hombres. En este mismo sentido, en otro estudio se reportó una interacción significativa géneroXgenotipoXmaltrato sobre el puntaje de agresión física, esto es, los hombres portadores del alelo de baja actividad de MAO-A con eventos traumáticos en la niñez presentaron mayores puntajes de agresión física (Frazzetto et al., 2007). Por otro lado, en un estudio cuyo objetivo fue replicar lo propuesto por Caspi y colaboradores, se reportó que el maltrato en la adolescencia fue un factor de riesgo para el desarrollo de problemas conductuales como rasgos antisociales y violencia, sin embargo, se obsevó que las variantes de la MAO-A no mostraron asociación ni interacción con el maltrato como un factor de riesgo para el desarrollo de conducta antisocial (Huizinga et al., 2006). En otro estudio se encontró que el genotipo MAO-A fue capaz de distinguir los hombres, tanto la pertenencia a pandillas como el uso de armas. El análisis de frecuencias de alelos reveló que entre los hombres miembros de pandillas y los usuarios de armas había mayor número de portadores del alelo de baja actividad de MAO-A (Beaver, DeLisi, Vaughn & Barnes, 2010).
En resumen, estos estudios demuestran consistentemente que factores como el género masculino y ser portador del alelo MAOA-L, en interacción con maltrato infantil, incrementan la susceptibilidad para el desarrollo de rasgos antisociales y de agresión física y violencia. Lo anterior va en la misma línea con lo reportado en un metanálisis en el cual se concluyó que la variación del genotipo MAO-A modera la relación entre maltrato en la niñez y la presencia de problemas mentales, p. e. trastorno antisocial, esto es, los hombres MAOA-L son los más vulnerables a los efectos de los eventos estresantes y son más proclives al desarrollo de rasgos antisociales en la adultez (Kim-Cohen et al., 2006).
La dopamina es un neurotransmisor del grupo de las monoaminas que regula funciones cerebrales como la actividad locomotora, la recompensa y la cognición. En algunos estudios se ha señalado un papel del sistema dopaminérgico en algunos rasgos característicos de la conducta agresiva como el enojo y la impulsividad. Se ha sugerido que la dopamina está relacionada al enojo y la delincuencia (en las formas agresivas y no agresivas), el metilfenidato reduce los ataques de ira, conductas antisociales como los robos, y episodios de agresión. El aripiprazol que es un agonista de la dopamina se ha relacionado con la reducción del enojo en el trastorno limítrofe de la personalidad (Dmitrieva, Chen, Greenberger, Ogunseitan & Ding, 2010).
El receptor de dopamina D4 (DRD4) es una proteína G acoplada al receptor que se expresa principalmente en la corteza prefrontal, región que modula funciones como la orientación de la conducta, la categorización y el filtro de claves ambientales (Avale et al., 2004); además, se ha asociado a la reactividad del estriado ventral en relación al sistema de la recompensa (Dmitrieva et al., 2010).
Se ha identificado un polimorfismo VNTR en el gen DRD4, localizado en el tercer exón y caracterizado por una secuencia de 48 pares de bases que se pueden repetir de 2 a 11 veces. Se ha sugerido que los alelos cortos, constituídos por la variantes de 2 a 5 repeticiones presenta una función diferencial comparado con los alelos largos, que incluye a las variantes de 6 a 11 repeticiones (Lusher, Chandler & Ball, 2004; Vieyra, Moraga, Henríquez, Aboitiz & Rothhammer, 2003).
En particular, las diferencias funcionales del alelo de 7 repeticiones del DRD4 consisten en la reducción en la inhibición postsináptica, lo cual resulta en una mayor reactividad del estriado ventral en relación a la recompensa. Algunos estudios han demostrado que existe una relación entre el alelo de 7 repeticiones del DRD4 y trastornos neuropsiquiátricos como el TDAH, el rasgo de búsqueda de la novedad y el abuso de sustancias (Vieyra et al., 2003), con delincuencia en la adolescencia, temperamento explosivo y búsqueda del peligro en hombres comparados con mujeres (Dmitrieva et al., 2010), y con agresión física en pacientes con esquizofrenia (Fresán et al., 2007).
Un método interesante en el estudio de la contribución genética a la conducta, es el análisis del efecto de las interacciones entre distintos genes-epistasis- sobre una característica en particular, este modelo pretende probar el efecto de un conjunto de genes en el desarrollo de un fenotipo. A la fecha no se han reportado investigaciones sobre epistasis y conducta violenta, así como el efecto de estas interacciones sobre constructos asociados a la conducta violenta como la hostilidad, impulsividad, reactividad interpersonal y rasgos psicopáticos.
Se ha propuesto que el incremento en la impulsividad se relaciona con conductas agresivas, sin embargo, el incremento en niveles de hostilidad, enojo, la presencia de rasgos psicopáticos, sobre todo las tendencias antisociales, y la falta de empatía se consideran factores de riesgo para la violencia (Hare & Neumman, 2008; Kennealy, Skeem & Walters, 2010;Lovett & Sheffield, 2007; Oquendo et al., 2001; Romero & Ostrosky, 2010;Suter et al., 2002; Yang et al., 2010;).
El objetivo del presente estudio fue investigar la presencia de epistasis entre variantes genéticas de la MAO-A y DRD4 sobre mediciones de agresión, hostilidad, impulsividad, reactividad interpersonal, rasgos de psicopatía y enojo en una población sana.
Método
Participantes
Participaron 60 hombres adultos sanos, con edades entre 21 y 59 años (M=29.8, DE=7.8), con una escolaridad entre 12 y 18 años (M=15.4, DE=1.7).
Los participantes fueron reclutados de diversos lugares de la Cd. de México, firmaron un consentimiento informado y fueron entrevistados para descartar antecedentes psiquiátricos, neurológicos y de abuso de sustancias, posteriormente se aplicaron los cuestionarios y se obtuvieron las muestras bucales para obtener el ADN. Todas las evaluaciones se llevaron a cabo en la Facultad de Psicología de la Universidad Nacional Autónoma de México. Se les mencionó que todos los datos eran confidenciales y serían utilizados con fines de investigación.
Extracción de ADN y genotipificación
A todos los participantes se les tomó una muestra de células bucales utilizando el Kit Gentra Puregen Buccal Cell (Qiagen), para la obtención del ADN. El análisis del polimorfismo MAO-A-uVNTR fue realizado por el método de reacción en cadena de la polimerasa (PCR). Las secuencias de los oligonucleótidos utilizados en este trabajo fueron, orientación sense: 5'-ACA GCC TGA CCG TGG AGA AG-3', orientación antisense: 5'-GAA CGG ACG ACG CTC CAT TCG GA-3'. La reacción de PCR se realizó en un volumen final de 12.5 µl conteniendo 1.5 mM de MgCl2, 200 µM de cada oligonucleótido, 0.2 µM de dNTPs (dATP, dCTP, dGTP, dTTP), 0.25 U de Go Taq Flexi de promega y 50 ng de ADN genómico. Después de 4 min. de desnaturalización a 95 °C, se realizaron 35 ciclos con las siguientes condiciones: 1 min. a 95°C, 1 min. a 62°C y 1 min. a 72°C. Se finalizó con un paso de 4 min. a 72°C. . Los productos de PCR fueron analizados mediante electroforesis en geles de agarosa/Metaphor al 2.5% y visualizados con luz UV después de teñirlos con bromuro de etidio.
El análisis del polimorfismo DRD4 fue realizado por el método PCR. Las secuencias de los oligonucleótidos utilizados en este trabajo fueron, D4-3 (5'-GCG ACT ACG TGG TCT ACT CG-3') y D4-42 (5'AGG ACC CTC ATG GCC TTG-3'). La reacción de PCR se realizó en un volumen final de 12.5 µl conteniendo 2 mM de MgCl2, 200 µM de cada oligonucleótido, 0.2 µM de dNTPs (dATP, dCTP, dTTP) con 0.1 µM de dGTP y 0.1 µM de 7-deaza-GTP, 10% de DMSO, 0.5 U de DNA polimerasa y 120 ng de ADN genómico. Después de 3 min. de desnaturalización a 95°C, se realizaron 40 ciclos con las siguientes condiciones: 30 seg. min. a 95°C, 30 seg. a 54°C y 1 min. a 72°C. Se finalizó con un paso de 4 min. a 72°C. Los productos de PCR fueron analizados mediante electroforesis en geles de agarosa/ Metaphor al 2.2% y visualizados con luz UV después de teñirlos con bromuro de etidio. El tamaño de los alelos se determino al comparar las bandas con una escalera de ADN de 50 pb.
Los sujetos fueron divididos de acuerdo a los genotipos de MAO-A (alta actividad vs. baja actividad) y DRD4 (no alelo 7 -no7R- vs. portadores de al menos un alelo de 7 repeticiones -7R-).
Instrumentos
Cuestionario de agresión reactiva y proactiva RPQ (Raine et al., 2006) en su versión en español (Andreu, Peña & Ramírez, 2009). Es un instrumento utilizado para medir la agresión reactiva y proactiva está compuesto por 23 ítems distribuidos en 2 factores, agresión reactiva y agresión proactiva, con una consistencia interna global de .91. Los ítems son puntuados en una escala de frecuencia (0=nunca, 1=a veces, y 2=a menudo).
Inventario de hostilidad de Buss-Durkee BDHI (Buss & Durkee, 1957), en su versión en español (Oquendo et al., 2001). Es un instrumento formado por 75 ítems Verdadero/Falso, está compuesto por una puntuación total y 8 subescalas: violencia, hostilidad indirecta, irritabilidad, negativismo, resentimiento, recelos, hostilidad verbal y culpabilidad. Tiene una consistencia interna global de .86.
Escala de impulsividad de Plutchik IP (Plutchik & Van Pragg, 1989) en su versión en español (Páez et al., 1996). Mide la tendencia para involucrarse en conductas impulsivas, consta de 15 ítems contestados en una escala de frecuencia de 3 puntos. Se compone de 4 subescalas: autocontrol, planeación de las acciones, conductas fisiológicas y actuación espontánea. Tiene una consistencia interna global de .66.
Escala de enojo de Novaco (Novaco, 1994). Mide la inclinación general a tener reacciones de enojo, consiste de 2 partes. La parte A consiste de 48 ítems agrupados en 3 subescalas: cognitiva, activación y conductual. La parte B consiste en 25 ítems que describen situaciones que podrían inducir enojo, se agrupa en 5 subescalas de acuerdo a la naturaleza de la situación que desencadena el enojo: trato irrespetuoso, injusticia, frustración, rasgos molestos de los demás e irritación. Tiene una consistencia internal global de .97.
Índice de reactividad interpersonal IRI (Davis, 1980) en su versión en español (Pérez, de Paúl, Etxeberría, Montes & Torres, 2003). Es un instrumento que evalúa la empatía disposicional. Se compone de 28 ítems distribuidos en 4 factores: perspective taking, fantasy, empathic concerní y personal distress. La consistencia interna de los factores oscila entre .70 y .78.
Escala de psicopatía de Levenson EPL (Levenson, Kiehl & Fitzpatrick, 1995). Mide los rasgos de psicopatía en población no institucionalizada, consta de 26 ítems distribuidos en 2 factores: psicopatía primaria y psicopatía secundaria con una consistencia interna de .82 y .63 respectivamente.
Análisis estadístico
Para investigar los efectos principales y las interacciones se generaron modelos de ANOVA factorial mediante el modelo lineal general, se incluyeron los puntajes de las escalas como variables dependientes y los genotipos MAO-A (alta vs. baja) y DRD4 (no7R vs. 7R) como factores entre sujetos, se corrigió el nivel de significancia mediante el método de Bonferroni para comparaciones múltiples. Los datos se analizaron usando el paquete estadístico SPSS 17.
Resultados
Distribución de alelos y genotipos
Los alelos de MAO-A se distribuyeron en 68.3% de alta actividad y 31.7% de baja actividad, mientras que los genotipos de DRD4 se distribuyeron en 2/3 (1.7%), 2/4 (3.3%), 2/7 (1.7%), 3/4 (1.7%), 3/7 (1.7%), 4/4 (40%), 4/6 (3.3%), 4/7 (33.3%), 4/8 (1.7%), 6/6 (3.3%), 6/7 (1.7%), 7/7 (6.7%). En el análisis del polimorfismo del gen DRD4 se comparó entre los no7R (2/3, 2/4, 3/4, 4/4, 4/6, 4/8 y 6/6; 55%) contra los 7R (2/7, 3/7, 4/7, 6/7 y 7/7; 52.9%).
Efecto principal de MAO-A
La variación genética de MAO-A tuvo efecto sobre mediciones de hostilidad, impulsividad, empatía y enojo, en el que los portadores del alelo de baja actividad presentaron un incremento en el puntaje de la subescala de resentimiento del BDHI respecto a los portadores del alelo de alta actividad F (1, 53)=5.19, p=.027; en el puntaje total de la escala de impulsividad de Plutchik F (1, 54)=4.29, p=.043 y en la subescala de actuación espontánea de la misma escala F (1, 54)=4.71, p=.035. En la subescala de angustia personal del IRI F (1, 49)=5.59, p=.022 y por último en el puntaje de duración del enojo del factor activación de la parte A del NAS F (1, 55)=4.22, p=.045.
Efecto principal de DRD4
Los sujetos portadores del alelo de 7 repeticiones tuvieron incrementos en los puntajes de mediciones de hostilidad, impulsividad, rasgos antisociales de la psicopatía y enojo. Específicamente estos incrementos se presentaron en la subescala de hostilidad verbal del BDHI F(1, 52)=4.85, p=.032, en la subescala de planeación de la escala de impulsividad de Plutchik F (1, 54)=4.22, p=.045, en los rasgos antisociales de psicopatía de la escala de Levenson F(1, 51)=6.22, p=.016, en el puntaje de expresión indirecta del enojo del factor conductual de la parte A del NAS F (1, 55)=8.98, p=.004 y en el puntaje de frustración de la parte B del NAS F (1, 55)=4.69, p=.035.
El alelo de 7 repeticiones se expresó como un decremento en el puntaje de la subescala de preocupación empática del IRI F (1, 49)=5.76, p=.02. Epistasis
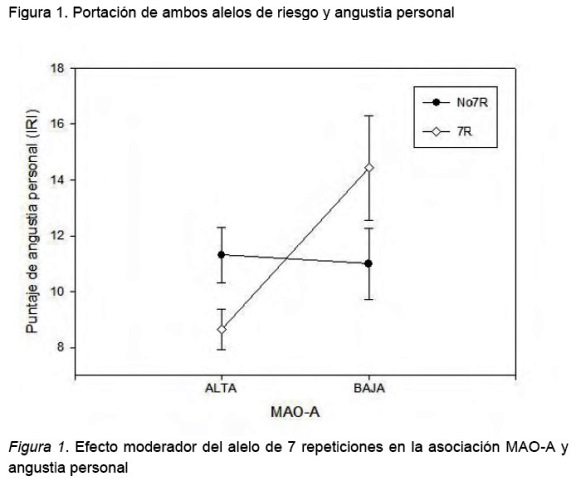
Se encontró una interacción significativa entre ambos genes en el puntaje de angustia personal del IRI F (1, 49)=6.94, p=.011, esta interacción se expresó como un incremento significativo en este puntaje en los sujetos portadores de ambos alelos de riesgo el de 7 repeticiones de DRD4 y el de baja actividad de MAO-A (tabla 1 y figura 1).

Discusión
Este estudio fue diseñado para investigar los efectos principales y las posibles interacciones entre 2 genes considerados de riesgo para el desarrollo de conductas violentas y de rasgos asociados a estas como hostilidad, impulsividad, empatía, psicopatía y enojo, el alelo de baja actividad de MAO-A y el alelo de 7 repeticiones del DRD4.
La variante de baja actividad transcripcional del gen de MAO-A se asocia a un incremento de rasgos hostiles, en particular el resentimiento, se considera que altos niveles de hostilidad preceden al desarrollo de conductas agresivas impulsivas (Oquendo et al., 2001), el resentimiento medido mediante el BDHI se refiere a la expresión de celos y odio hacia los demás, lo cual puede indicar que estos sujetos tienden a sentir estos afectos negativos. Esta variante de igual forma impactó sobre las mediciones de impulsividad, particularmente sobre la conducta reflexiva y descontrolada (Páez et al., 1996), Beaver et al. (2010) reportaron que los hombres con baja actividad de MAO-A presentan conductas antisociales con un componente impulsivo importante como las agresiones con armas. La baja actividad de MAO-A también tuvo un efecto sobre la duración del enojo fisiológicamente hablando, esta hiperactivación fisiológica que de acuerdo a nuestros resultados presentaron los sujetos con baja actividad de MAO-A se asocia a la interpretación cognitiva de estímulos ambiguos que subjetivamente se etiquetan como estímulos que provocan enojo, a pesar de que el enojo no necesariamente resulta en conductas antisociales, se ha identificado que la intervención enfocada al manejo del enojo resulta en una disminución de episodios violentos (Suter et al., 2002).
Otro de los resultados indica que la baja actividad de MAO-A se expresó como un incremento en el puntaje de angustia personal del índice de reactividad interpersonal, esta subescala mide la tendencia de un sujeto a experimentar sentimientos de incomodidad al ser testigo de experiencias negativas en otras personas tales como situaciones emocionalmente tensas o estresantes como el rechazo social (Pérez et al., 2003), este resultado concuerda con los hallazgos más consistentes acerca de la vulnerabilidad que presentan los portadores del alelo de baja actividad (MAOA-L) que se expresa como una hipersensibilidad ante eventos adversos como el maltrato infantil que, de acuerdo a estos estudios, los pone más en riesgo de cometer conductas violentas de tipo impulsivo a manera de defensa, sin embargo, estas conductas son reacciones exageradas (Caspi et al., 2002; Frazzetto et al., 2007; Kim-Cohen et al., 2006).
Debido a que la MAO-A tiene un efecto en el cerebro resulta interesante tomar una perspectiva neurobiológica, en este sentido, la hiper-reactividad ante situaciones emocionalmente adversas se ha descrito en los portadores del alelo de baja actividad. En un estudio de resonancia magnética estructural (MRI) y funcional (fMRI) al realizar una tarea de activación emocional (caras de enojo y miedo) los sujetos con baja actividad de MAO-A presentaron una actividad incrementada en la amígdala y un decremento en el subgenual, supragenual -estas estructuras participan en la interpretación de situaciones emocionales negativas-, corteza cingulada ventral, corteza orbital lateral izquierda y corteza insular izquierda, además en una tarea de memoria emocional (codificación y evocación de información aversiva) estos sujetos mostraron mayor actividad de la amígdala izquierda y la formación hipocampal en la evocación del material por lo tanto se considera que un sistema compuesto por la corteza prefrontal (orbital), cíngulo, amígdala e hipocampo participa en la regulación emocional que sugiere las bases de los mecanismos neurales de la tendencia genética a la violencia impulsiva (Meyer-Lindenberg et al., 2006).
Se ha hipotetizado que el genotipo de baja actividad de MAO-A confiere hipersensibilidad al rechazo social lo cual podría resultar en una predisposición genética al desarrollo de rasgos agresivos. En un estudio de fMRI los hombres MAOA-L mostraron mayor nivel de agresión y de hipersensibilidad interpersonal así como un incremento en la actividad de la corteza cingulada anterior ante una condición de exclusión social, lo cual sugiere que la MAO-A y la agresión se relacionan mediante la hipersensibilidad socioemocional (Eisenberger, Way, Taylor, Welch & Liebermann, 2007), además se considera que la sensibilidad emocional es fundamental para la interacción social (Spinella, 2005).
El alelo de 7 repeticiones se asoció con mayores puntajes de hostilidad verbal, falta de planeación, psicopatía secundaria, expresión indirecta del enojo, mayor disposición a la frustración; y con un menor puntaje de preocupación empática. Los resultados del incremento en el puntaje de dichas mediciones concuerdan con lo propuesto anteriormente acerca del papel de la dopamina en la regulación de conductas antisociales (Dmitrieva et al., 2010).
Los estudios cognitivos y de neuroimagen han demostrado que el alelo de 7 repeticiones se asocia con un pobre desempeño en tareas de inhibición conductual (Langley et al., 2004), con un efecto sobre el volumen de la corteza prefrontal la cual participa en el control inhibitorio (Durston et al., 2005), y con una mayor reactividad del estriado ventral que se expresa en un fenotipo impulsivo (Forbes et al., 2009) nuestros resultados evidencian un incremento en los niveles de impulsividad, lo cual sugiere que aún en población sana este alelo produce un fenotipo impulsivo lo cual se ha reportado previamente en sujetos con TDAH y con abuso de sustancias (Vieyra et al., 2003). El incremento en la tendencia a la frustración, impulsividad y hostilidad presentado por los sujetos 7R sugiere que este alelo confiere riesgo al desarrollo de conductas agresivas impulsivas, estas conductas ocurren en presencia de provocaciones o la percepción subjetiva de amenazas, por definición son conductas no planeadas y defensivas en naturaleza y se asocian a enojo y frustración. Este tipo de agresión tiene su origen en las teorías de "frustración-agresión" y se define operacionalmente como la "agresión en respuesta a la provocación" (Nouvion, Cherek, Lane, Tcheremissine & Lieving, 2007). En este sentido se ha propuesto que los rasgos antisociales de la psicopatía predicen tanto la conducta violenta como la comisión de crímenes violentos (Kennealy et al., 2010; Romero & Ostrosky, 2010; Yang et al., 2010), los sujetos 7R presentaron mayores niveles de psicopatía secundaria que se caracteriza por altos niveles de impulsividad, lo cual los pone en riesgo de desarrollar conductas violentas.
En cuanto al decremento en el puntaje de preocupación empática que se refiere a la tendencia a experimentar sentimientos de compasión y preocupación hacia otros, se han reportado disfunciones prefrontales que correlacionan negativamente con mediciones de empatía en sujetos sanos (Spinella, 2005), lo anterior aunado al fenotipo impulsivo de los portadores del alelo de 7 repeticiones sugiere que este alelo confiere riesgo al desarrollo de conductas agresivo-impulsivas.
Un resultado interesante fue el de la interacción significativa de ambos genes sobre el puntaje de angustia personal lo cual sugiere que la variación genética de DRD4 modera la relación entre la baja actividad de MAO-A y la hipersensibilidad ante situaciones emocionalmente negativas, esto es, la baja actividad de MAO-A confiere hiperreactividad ante situaciones estresantes la cual se incrementa si un sujeto es portador de al menos un alelo de 7 repeticiones de DRD4.
Nuestros resultados sugieren que ambos genes producen un fenotipo caracterizado por el enojo, la hostilidad y la impulsividad que podrían conferir riesgo para el desarrollo de conductas violentas impulsivas (Hare & Neumman, 2008; Kennealy et al., 2010; Lovett & Sheffield, 2007; Oquendo et al., 2001; Romero & Ostrosky, 2010; Suter et al., 2002; Yang et al., 2010;). Es importante señalar que estos alelos de riesgo, considerando nuestros resultados, no necesariamente se relacionan con episodios violentos, en futuras investigaciones es necesaria la medición de estos episodios para poder establecer una relación entre el incremento en el enojo, hostilidad e impulsividad y la comisión de conductas violentas.
Estos resultados son interesantes, ya que, abren la posibilidad de investigar endofenotipos que sean más cercanos a los efectos fisiológicos de la variación genética, tales como el desempeño neuropsicológico o el volumen y activación cerebrales, y de esta manera explorar si existe relación entre las mediciones conductuales medidas mediante escalas y los endofenotipos. Consideramos que la investigación de endofenotipos permitiría aclarar el efecto diferencial de ambos genes, esto es, ambos genes tuvieron efecto sobre distintos componentes de los constructos y esto tal vez se deba a la expresión que cada gen tiene sobre el volumen o la función cerebrales.
Un aspecto importante de esta investigación es el estudio de las interacciones entre genes, ya que, se ha propuesto que la función de un gen puede modificar la actividad fenotípica de otro gen, en futuras investigaciones, sería interesante estudiar si existe epistasis entre estos genes y otros que se han asociado a la conducta violenta, así como con otros que aunque no se asocien con violencia tengan algún efecto sobre la actividad de MAO-A y dopamina.
En conclusión podemos sugerir que variaciones genéticas de MAO-A y DRD4 tiene un efecto sobre mediciones relacionadas a la conducta violenta, expresadas como un efecto principal y como la interacción entre los alelos de riesgo que incrementan la reactividad ante situaciones estresantes lo cual se considera un factor de riesgo para el desarrollo de conductas violentas.
Referencias
Andreu, J., Peña, E. y Ramírez, J. (2009). Cuestionario de agresión reactiva y proactiva: un instrumento de medida de la agresión en adolescentes. Revista de Psicopatología y Psicología Clínica, 14, 1, 37-49. [ Links ]
Avale, M., Falzone, T., Gelman, D., Low, M., Grandy, D. & Rubinstein, M. (2004). The dopamine D4 receptoris essential for hyperactivity and impaired behavioral inhibition in a mouse model of attention deficit/hyperactivity disorder. Molecular Psychiatry, 1, 718-726. [ Links ]
Beaver, K., DeLisi, M., Vaughn, M. & Barnes, J. (2010). Monoamine oxidase A genotype is associated with gang membership and weapon use. Comprehensive Psychiatry, 51, 130-134. [ Links ]
Brunner, H., Nelen, M., Breakefield, X., Ropers, H. & van Oost, B. (1993). Abnormal behavior associated with a point mutation in the structural gene for monoamine oxidase A. Science, 262, 5133, 578-580. [ Links ]
Buss, A. & Durkee, A. (1957). An inventory for assessing different kin, DS of hostility. Journal of Consulting and Clinical Psychology, 21, 343-349. [ Links ]
Cases, O., Seif, I., Grimsby, J., Gaspar, P., Chen, K., Pournin, S., ... De Maeyer, E. (1995). Aggressive behavior and altered amounts of brain serotonin and norepinephrine in mice lacking MAOA. Science, 268, 5218, 1763-1766. [ Links ]
Caspi, A., McClay, J., Moffitt, T., Mill, J., Martin, J., Craig, I., ... Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children. Science, 297, 5582, 851-854. [ Links ]
Davis, M. H. (1980). A multidimensional approach to individual differences in empathy. Catalog of Selected Documents in Psychology, 10, (85), 1-17. [ Links ]
Dmitrieva, J., Chen, C., Greenberger, E., Ogunseitan, O. & Ding, Y. (2010). Gender-specific expression of the DRD4 gene on adolescent delinquency, anger and thrill seeking. Social Cognitive and Affective Neuroscience, 6, (1), 82-89. [ Links ]
Durston, S., Fossella, J., Casey, B., Hulshoff, H., Galvan, A., Schnack, H., ... van Engeland H. (2005). Differential effects of DRD4 and DAT1 genotype on fronto-striatal gray matter volumes in a sample of subjects with attention deficit hyperactivity disorder, their unaffected siblings, and controls. Molecular Psychiatry, 10, 678-685. [ Links ]
Eisenberger, N., Way, B., Taylor, S., Welch, W. & Lieberman, M. (2007). Understanding genetic risk for aggression: Clues from the brain's response to social exclusion. Biological Psychiatry, 61, 1100-1108. [ Links ]
Forbes, E., Brown, S., Kimak, M., Ferrell, R., Manuck, S. & Hariri, A. (2009). Genetic variation in components of dopamine neurotransmission impacts ventral striatal reactivity associated with impulsivity. Molecular Psychiatry, 14 (1), 60-70. [ Links ]
Frazzetto, G., Di Lorenzo, G., Carola, V., Proietti, L., Sokolowska, E., Siracusano, A., . Troisi, A. (2007). Early trauma and increased risk for physical aggression during adulthood: the moderating role of MAOA genotype. PLoSONE, 2 (5). 1-6. [ Links ]
Fresán, A., Camarena, B., Apiquian, R., Aguilar, A., Urraca, N. & Nicolini, H. (2007). Association study of MAOA and DRD4 genes in schizophrenic patients with aggressive behavior. Neuropsychobiology, 55, 171-175. [ Links ]
Hare, R. & Neumman, C. (2008). Psychopathy as a clinical and empirical construct. Annual Review of Clinical Psychology, 4, 217-246. [ Links ]
Huizinga, D., Haberstick, B., Smolen, A., Menard, S., Young, S., Corley, R., ... Hewitt, J. (2006). Childhood maltreatment, subsequent antisocial behavior, and the role of monoamine oxidase A genotype. Biological Psychiatry, 60, 677-683. [ Links ]
Kennealy, P., Skeem, J. & Walters, G. (2010). Do core interpersonal and affective traits of PCL-R psychopathy interact with antisocial behavior and disinhibition to predict violence?. Psychological Assessment, 22 (3), 569-580. [ Links ]
Kim-Cohen, J., Caspi, A., Taylor, A., Williams, B., Newcombe, R., Craig, I. & Moffitt, T. (2006). MAOA, maltreatment, and gene-environment interaction predicting children's mental health: new evidence and a meta-analysis. Molecular Psychiatry, 11, 903-913. [ Links ]
Langley, K., Marshall, L., van den Bree, M., Thomas, H., Owen, M., O'Donovan, M. & Thapar, A. (2004). Association of the dopamine D4 receptor gene 7-repeat allele with neuropsychological test performance of children with ADHD. American Journal of Psychiatry, 161, (1), 133-138. [ Links ]
Levenson, M., Kiehl, K. & Fitzpatrick, C. (1995). Assessing psychopathic attributes in a noninstitutionalized population. Journal of Personality and Social Psychology, 68, (1), 151-158. [ Links ]
Lovett, B. & Sheffield, R. (2007). Affective empathy deficits in aggressive children and adolescents: a critical review. Clinical Psychology Review, 27, 1 -13. [ Links ]
Lusher, J., Chandler, C. & Ball, D. (2004). Alcohol dependence and the alcohol Stroop paradigm: Evidence and issues. Drug and Alcohol Dependence, 75, (3), 225-231. [ Links ]
Meyer-Lindenberg, A., Buckholtz, J., Kolachana, B., Hariri, A., Pezawas, L., Blasi, G., ... Weinberger, D. (2006). Neural mechanisms of genetic risk for impulsivity and violence in humans. PNAS, 103 (16), 6269-6274. [ Links ]
Nouvion, S., Cherek, D., Lane, S., Tcheremissine, O. & Lieving, L. (2007). Human proactive aggression: association with personality disorders and psychopathy. Aggressive Behavior, 33, 552-562. [ Links ]
Novaco, R. (1994). Anger as a risk factor for violence among the mentally disordered. En J. Monohan & H. Steadman (Eds.), Violence and mental disorder (pp. 21-59). Chicago: University of Chicago Press. [ Links ]
Oquendo, M., Graver, R., Baca-García, E., Morales, M., Montalván V. & Mann, J. (2001). Spanish adaptation of the Buss-Durkee Hostility Inventory (BDHI). The European Journal of Psychiatry, 15 (2), 101 -112. [ Links ]
Páez, F., Jiménez, A., López, A., Ariza, R., Paulo, J., Ortega, H. & Nicolini, H. (1996). Estudio de validez de la traducción al castellano de la Escala de Impulsividad de Plutchik. Salud Mental, 19 (3), 10-12. [ Links ]
Pérez, A., de Paúl, J., Etxeberría, J., Montes, M. y Torres, E. (2003). Adaptación de Interpersonal Reactivity Index (IRI) al español. Psicothema, 15 (2), 267-272. [ Links ]
Plutchik, R & Van Pragg, H. (1989). The measurement of suicidality, aggressivity and impulsivity. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 13, 23-34. [ Links ]
Raine, A., Dodge, K., Loeber, R., Gatzke-Kopp, L., Lynam. D., Reynolds, C., . Liu, J. (2006). The reactive.proactive aggression questionnaire: differential correlates of reactive and proactive aggression in adolescent boys. Aggressive Behavior, 32, 159-171. [ Links ]
Rhee, S. H. & Waldman, I. D. (2002). Genetic and environmental influences on antisocial behaviour: Meta-analysis of twin and adoption studies. Psychology Bulletin, 128, 633-645. [ Links ]
Romero, C. y Ostrosky, F. (2010). Efecto de las cogniciones implícitas y la psicopatía sobre la comisión de crímenes violentos. Manuscrito no publicado. Laboratorio de Neuropsicología y Psicofisiología, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Sabol, S., Hu, S. & Hamer, D. (1998). A functional polymorphism in the monoamine oxidase A gene promoter. Human Genetics, 103 (3), 273-279. [ Links ]
Shih, J., Chen, K. & Ridd, M. (1999). Monoamine oxidase: From Genes to Behavior. Annual Review of Neuroscience, 22, 197-217. [ Links ]
Spinella, M. (2005). Prefrontal substrates of empathy: psychometric evidence in a community sample. Biological Psychology, 70, 175-181. [ Links ]
Suter, J., Byrne, M., Byrne, S., Howells, K. & Day, A. (2002). Anger in prisoners; women are different from men. Personality and Individual Dfferences, 32, 1087-1100. [ Links ]
Vieyra, G., Moraga, M., Henríquez, H., Aboitiz, F. & Rothhammer, F. (2003). Distribution of DRD4 and DAT1 alleles from dopaminergic system in a mixed Chilean population. Revista Médica de Chile, 131 , 135-143. [ Links ]
Yang, M., Wong, S. & Coid, J. (2010). The efficacy of violence prediction: a metaanalytic comparison of nine risk assessment tools. Psychological Bulletin,136 (5), 740-767. [ Links ]
Nota
Agradecimiento. Proyecto financiado por el Instituto de Ciencia y Tecnología del Gobierno del Distrito Federal 422.01 PICDS08-19.

