











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
El maíz (Zea mays L.) es uno de los cultivos más importantes en el mundo desde el punto de vista socioeconómico y cultural; para el año 2020 se registraron 187 millones de hectáreas sembradas con el cultivo a nivel mundial (Cuevas-Mejía 2014, Karki et al. 2020). El maíz se cultiva bajo distintos sistemas de manejo agrícola desarrollados acorde a las características socio-ambientales de una región geográfica particular (Kato-Yamakake et al. 2009, Altieri & Nicholls 2013).
En México, uno de los sistemas de manejo más utilizados es la milpa: agroecosistema integrado por diferentes cultivos que se rota espacialmente con la vegetación silvestre en varios ciclos productivos. En ella, el maíz se combina con diferentes especies, como frijol (Phaseolus sp.) y calabaza (Cucurbita sp.), aunque esta composición varía por región, características de cada parcela, condiciones ambientales y preferencias del productor (Kato-Yamakake et al. 2009, Mariaca 2011). Por lo general este sistema depende de la lluvia como fuente de agua (agricultura de temporal) (Nigh & Diemont 2013).
La integración de diversas especies en la milpa provee beneficios a las plantas que lo integran, como reducción de temperatura, retención de humedad dentro del área de cultivo, control de arvenses, e incrementa la disponibilidad de nutrientes en el suelo (Sánchez-Morales & Hernández-Ortiz 2014, Pérez-Hernández et al. 2021). El conjunto de estos beneficios contribuye a una mayor resiliencia de los cultivos a perturbaciones externas, y al mismo tiempo, se puede llegar a superar a los monocultivos de maíz en términos productivos (Ebel et al. 2017). De Igual forma, el sistema milpa provee otras ventajas importantes como la conservación de la de biodiversidad, proveer de una diversidad de alimentos y nutrientes a los agricultores y sus familias (Novotny et al. 2021), además de que forma parte de la identidad y prácticas de algunas culturas en México, como la cultura Maya (García-Quintanilla & Reyes-García 2023).
En la Península de Yucatán, al sureste de México, la milpa (conocida como milpa Maya), se realiza mediante el sistema roza, tumba y quema para convertir un área de selva secundaria en un campo de cultivo. Para la roza se cortan las hierbas y arbustos, se cortan los árboles (tumba), y se quema la biomasa cortada para liberar nutrientes, después de lo cual, se realiza la siembra. Después de varios ciclos de cultivo, la fertilidad del suelo disminuye, por lo que se intercala con un periodo de descanso de la tierra (barbecho), que permite la regeneración de la vegetación secundaria que repone nutrientes del suelo, controla plantas arvenses y aumenta la disponibilidad de agua (Hernández 1992, Mariaca 2011, Terán-Contreras & Rasmussen 2009, Lara-Ponce et al. 2012, Nigh & Diemont 2013).
Si bien la milpa tradicional sigue en práctica, se ha incrementado el número de agricultores que optan por sembrar maíz en monocultivo con insumos externos, mientas que otros abandonan esta práctica en su totalidad derivado de la escasez de mano de obra y el bajo o nulo ingreso monetario que representa en la región (Salazar Barrientos et al. 2016, Ku-Pech et al. 2020). De igual forma, los intervalos de descanso de la tierra se han reducido, lo que implica que el rendimiento de las milpas decaiga por no tener el suelo los nutrientes necesarios (Bautista & Palacio 2012, Ebel et al. 2017, Ku-Pech 2019). Por ejemplo, se ha reportado que los rendimientos en el estado de Yucatán se han reducido aproximadamente 200 kg/ha en los últimos 25 años (Gómez-Rojas & Esquivel-Mota 2002, Rodríguez-Canto et al. 2016).
Aunado a lo anterior, el cambio climático afecta la productividad del cultivo, ya que el incremento de la temperatura y cambios en la distribución de lluvias inducen estrés hídrico en la planta por el aumento de la evapotranspiración (Jarma-Orozco et al. 2012). Esto conlleva a la deshidratación y disminución de la fotosíntesis, lo que altera el desarrollo, crecimiento y la productividad de las plantas (Squire 1990, Wahid et al. 2007). Los periodos de sequía afectan principalmente a los maíces que son cultivados en suelos con poca retención de agua, superficiales, con alta pedregosidad y que carecen de sistemas de riego (Moya-García et al. 2003, Terán-Contreras & Rasmussen 2009).
En este sentido, los sistemas de manejo agrícola pueden modificar el microclima dentro del área de cultivo. En cultivos establecidos al sur de México, se encontró que un monocultivo (M) de maíz presentaba una temperatura del suelo 4.4 °C mayor respecto a un policultivo Maíz-Frijol (MF); mientras que la retención de agua fue hasta 45 % mayor en un policultivo Maíz-Frijol-Calabaza (MFC, milpa) en comparación con el monocultivo (Pérez-Hernández et al. 2021). Además, las plantas de maíz en MF presentaron una tasa fotosintética y de transporte de electrones hasta 32 % mayor que las de M, mientras que la transpiración y la disipación no fotoquímica de energía fue mayor en plantas en MFC que en M. En MF y MFC las plantas fijaron mayor cantidad de CO2 que las de M, a pesar de que la eficiencia en el uso de agua fue similar en todos los casos. Dicho estudio se llevó a cabo en una zona de alta precipitación y suelos profundos, por lo que es necesario explorar el efecto del manejo en zonas donde la disponibilidad de agua es más limitada y con suelos menos profundos (superficiales).
Ante este panorama es recomendable desarrollar estudios fisiológicos que proporcionen información acerca de la adaptabilidad del maíz ante las condiciones ambientales de cada región (Ku-Pech 2019), lo cual contribuiría al entendimiento de la plasticidad del cultivo ante los efectos del cambio climático, como la reducción en la disponibilidad de agua en ambientes con poca capacidad de retención en suelo. Dado lo anterior, es necesario caracterizar prácticas que contribuyan a eficientizar recursos como el uso de nutrientes y agua en el suelo, a fin de identificar prácticas que contribuyan a reducir o mitigar los efectos del cambio climático, particularmente en regiones con baja precipitación y presencia de suelos superficiales y pedregosos, dada su alta vulnerabilidad ante este fenómeno.
Por lo anterior, el presente estudio tuvo como objetivo caracterizar el efecto que ejercen los sistemas de manejo monocultivo y milpa sobre el microclima dentro del área del cultivo, así como sobre la fisiología de la planta del maíz en dos sitios con diferentes condiciones de pedregosidad y profundidad del suelo. Esperábamos encontrar que la milpa de los dos sitios presentara un mayor contenido volumétrico de agua en el suelo, derivado de la mayor cobertura vegetal, y que las plantas de maíz en policultivo tuvieran un mejor desempeño fisiológico en comparación con el monocultivo, con una mayor tasa de fotosíntesis, mayor transpiración y conductancia estomática, una mayor eficiencia en el uso de agua, así como un mayor transporte de electrones, particularmente en el sitio con mayor pedregosidad y menor profundidad del suelo.
Materiales y métodos
Área de estudio. El estudio se realizó en el estado de Yucatán, al sureste de México, en donde se eligieron dos sitios caracterizados por contar con diferentes condiciones de pedregosidad y profundidad del suelo constantes. El primer sitio se ubica en el municipio de Peto (20º 017’ 57.40” N; 88º 58’ 10.08” O) a una altura de 35 m snm, con un clima cálido subhúmedo, temperatura media anual de 26.4 °C, y una precipitación total anual promedio de 1,100 mm (Ku-Pech et al. 2020, CLICOM 2022, INEGI 2010). El segundo sitio se localiza en el municipio de Mama (20º 28’ 16.49” N; 89º 22’ 02.97” O) a 24 m snm, un clima cálido subhúmedo, temperatura media anual de 26.1 °C y precipitación total anual promedio de 830 mm (CLICOM 2022, INEGI 2017). El suelo de Peto es un suelo Chak k’ankab, con una profundidad de 14.66 cm y con presencia de pedregosidad en el 60 % de la superficie utilizada en el experimento; el suelo del sitio de Mama es de tipo K’ankab, con una profundidad de 37.5 cm y con escasa pedregosidad (menor al 5 % de la superficie utilizada para el experimento).
Diseño experimental. En cada sitio se eligió una parcela de 600 m2, la cual fue dividida en bloques de 100 m2 (10 × 10 m). Por medio de un diseño de bloques al azar, se instalaron tratamientos para el cultivo del maíz bajo dos manejos contrastantes: A) monocultivo (M), y B) sistema milpa (combinación maíz-frijol-calabaza; MFC) con tres repeticiones (bloques de 100 m2) por tratamiento y sitio de trabajo. La siembra del maíz se realizó al inicio del periodo de lluvias en la región (junio 2022); la distancia de siembra de las semillas fue de 1 m entre filas y 60 cm entre plantas, se plantaron 45 g de maíz por bloque, y se tuvo una densidad de siembra de 200 plantas de maíz por bloque (equivalente a 20,000 plantas por ha). En el caso de la milpa las semillas del frijol y la calabaza fueron mezcladas con las del maíz previo a la siembra; los tres cultivos fueron sembrados al mismo tiempo, colocando las semillas del frijol o la calaza en el mismo hoyo donde se depositó el maíz. La distribución de las semillas de frijol y calabaza fueron azarosas dentro de cada bloque hasta agotar 7.5 g de frijol y 2.5 g de calabaza. Previo a la siembra del maíz, las arvenses se eliminaron manualmente; las semillas se sembraron a mano, sin aplicar labranza al suelo y colocando tres semillas de maíz (más una de frijol o calabaza cuando correspondió) en hoyos a 7 cm de profundidad.
Tanto las técnicas de sembrado como la elección de las variedades a sembrar se apegaron a las prácticas locales. Se empleó una variedad local de maíz conocida como Nal Xoy blanco, el cual presenta un ciclo corto para su cosecha (aprox. dos meses y medio), tiene tolerancia al estrés hídrico y un alto rendimiento en comparación con otras variedades (Chi Canul 2002, Gonzáles-Valdivia et al. 2017, Ku-Pech et al. 2020). El frijol (Phaseolus vulgaris L.) que se utilizó fue de la variedad Xcoli Bu’ul y la calabaza (Cucurbita moschata Duchesne ex Poir.) fue de la variedad Xnuk k’um.
Caracterización microclimática. Se caracterizó el microclima en cada bloque experimental (tres por tratamiento) mediante dos muestreos durante el desarrollo del maíz: a los 40 días después de la siembra (Material suplementario, Figura S1) cuando el maíz tenía aproximadamente 80 cm de altura y, a los 75 días después de la siembra, cuando las plantas de frijol y calabaza estaban bien desarrolladas. Los registros se tomaron durante tres días consecutivos, registrando datos cada 10 min durante las 24 h del día. Se utilizaron estaciones microclimáticas portátiles conformadas por un registrador de datos HOBO U30-NRC (Onset Computer Corporation, Massachusetts, USA), a la que se conectaron sensores para registrar: luz fotosintéticamente activa (PAR; S-LIA-M003), formación de rocío en las hojas (S-LWA-M003), temperatura y humedad relativa del aire (S-THB-M002), contenido volumétrico de agua en el suelo (CVA) (S-SMD-M005), y temperatura del suelo (S-TMB-M002). A partir de los datos de temperatura y humedad del aire, se calculó el déficit de presión de vapor de agua atmosférico (DPV) siguiendo a Jones (1992). A la par de los registros en cada bloque experimental, se colocó una estación microclimática en un sitio abierto ubicado a un costado de cada sitio experimental a manera de referencia; dicha estación contaba con el mismo equipamiento descrito anteriormente. Se midió la precipitación en cada uno de los sitios de manera ininterrumpida a lo largo del experimento (junio a noviembre), mediante un pluviómetro S-RGB-M002 conectado a un registrador H21, en donde se encontró que la precipitación fue de 721 mm para Peto y de 651.2 mm para Mama.
Caracterización fisiológica. Se realizaron muestreos fisiológicos al mismo tiempo que la caracterización microclimática. Se midieron cuatro plantas por bloque (12 plantas por tratamiento y sitio de trabajo), las cuales fueron seguidas a lo largo del experimento. Las mediciones realizadas fueron:
Intercambio de gases y fluorescencia de la clorofila.- Se caracterizó la asimilación de CO2 (A), transpiración (E) y conductancia estomática (gs) entre las 6:00 h y 18:00 h, tomando registros a intervalos de tres horas, mediante un analizador de gases infrarrojo LI-6800 (LI-COR Environmental, Lincoln, USA) equipado con una fuente de luz. El analizador de gases tenía integrado un fluorómetro que permitió caracterizar, al mismo tiempo de lo descrito anteriormente, la eficiencia cuántica del fotosistema II en plantas adaptadas a la luz (𝛷PSII) y adaptadas a la oscuridad (Fv/Fm), la tasa de transporte de electrones (TTE) y la disipación no-fotoquímica de energía (DNE). Se midió la cantidad de luz (medida como radiación fotosintéticamente activa; PAR por sus siglas en inglés) incidente en un espacio abierto de cada sitio de trabajo previo al inicio de cada medición, dicha cantidad de luz se fijó en la fuente de luz del LI-6800 para la medición correspondiente; la concentración de CO2 se fijó a 400 ppm, la temperatura de la cámara fue la ambiental, y la humedad relativa se fijó entre el 50 % y 60 %, acorde a las instrucciones del fabricante. Se presentan los datos de A y E registrados a lo largo del día, así como los totales diarios calculados como se indica a continuación (Pimienta-Barrios et al. 2003, LI-COR Technical Solutions 2015, Pérez-Hernández et al. 2021):
Dónde: Ai es la asimilación de CO2 instantánea (𝜇mol m-2s-1) de la planta i; E i es la transpiración instantánea (mmol m-2s-1) de la planta i; ∆T es el tiempo en segundos entre cada medición, calculado como:
Donde 180 min es el tiempo en minutos entre mediciones. A partir de la división de los totales diarios de A y E se calculó la eficiencia en el uso del agua total diaria (EUAtot; mmol CO2 m-2 d-1/ mol H2O m-2 d-1).
Contenido de clorofila.- Se caracterizó mediante un medidor portátil de concentración de clorofila MC-100 (Apogee Instruments, Inc., Utah, USA) en las mismas plantas mencionadas anteriormente; los datos se presentan en 𝜇mol m-2.
Estado hídrico.- Se colectaron muestras de hoja de las 12 plantas seleccionadas por tratamiento y sitio, previo al amanecer, las muestras se envolvieron en papel aluminio y estas se resguardaron dentro bolsas plásticas herméticamente selladas. Las muestras se almacenaron en una hielera para determinar: A) el potencial hídrico (Ψ) mediante un potenciómetro de punto de rocío WP4C (Decagon Devices, Inc., Washington, USA), este parámetro se midió en menos de 24 horas después de la colecta, y B) el contenido relativo de agua (CRA), para lo cual las muestras fueron pesadas (peso fresco), sumergidas en agua destilada hasta que el peso no varió (48 horas) y pesadas de nuevo (peso a saturación), y secadas en un secador artesanal a 60 ºC por 48 horas, después de lo cual se pesaron de nuevo (peso seco). El CRA se calculó siguiendo a González & González-Vilar (2001) como:
Crecimiento.- Durante cada muestreo se registró la altura de las 12 plantas por tratamiento con un flexómetro.
Producción de maíz.- Después de la cosecha, se seleccionaron cinco mazorcas de cada bloque, una vez secas se pesaron enteras, luego se desgranaron y se determinó el peso seco total de los granos obtenidos. En Peto no se pudo obtener cosecha ya que las mazorcas fueron consumidas por animales.
Análisis estadísticos. Para comparar los datos obtenidos del microclima (Temperatura del aire, temperatura del suelo, CVA y DPV), fisiología (E, gs, EUA, contenido de clorofila, CRA, Ψ, 𝛷PSII, TTE, DNF) y altura de las plantas, por sitio (Mama y Peto), día de medición (40 y 75) y tratamiento (M y MFC), se realizó un análisis de varianza factorial (ANOVA) con un α = 0.05 y una prueba post hoc de Tukey. Para el caso de A no hubo normalidad ni homogeneidad de varianzas, por lo que se utilizó un modelo lineal generalizado (GLM). Un análisis no métrico de escala multidimensional (NMDS por sus siglas en inglés) fue utilizado para identificar similitudes en las estrategias fisiológicas utilizadas por las plantas de maíz entre tratamientos y sitios de medición al día 75 (cuando las plantas de frijol y calabaza se encontraban bien desarrolladas). Para este análisis se utilizó el paquete vegan (Oksanen et al. 2019). Se calculó la covarianza de la salida de datos NMDS por tratamiento y sitio de medición para comparar los rangos de variación en dichos datos (este procedimiento no asume una relación lineal entre las variables). Se probó la correlación de las puntuaciones NMDS con las diferentes variables ambientales medidas, para analizar la relación entre los parámetros fisiológicos y microclimáticos. Los análisis se realizaron en R Software v. 4.3.0 (R Core Team 2023).
Resultados
Microclima. A los 40 días después de la siembra del maíz, no hubo diferencias entre tratamientos para las variables microclimáticas medidas en ninguno de los dos sitios (material suplementario Tabla S1). Sin embargo, al día 75 cuando las plantas de frijol y calabaza se encontraban bien desarrolladas, se encontró que en el tratamiento M recibía más luz respecto a MFC en ambos sitios de trabajo (Figura 1A y F, respectivamente). La temperatura mínima del aire fue hasta 1.8 ºC mayor en los tratamientos de Mama, respecto a los de Peto (F = 66.04; P < 0.05). La temperatura máxima en el tratamiento M en Peto fue 1.5 °C mayor que en MFC de Mama (F = 31.24; P < 0.05). La temperatura del aire en los tratamientos de Peto alcanzó su máximo a las 18:00 h, donde M presentó una temperatura promedio de 35.4 ± 0.4 °C y MFC de 34.5 ± 0.6 °C (Figura 1B). En Mama las temperaturas más altas se registraron entre las 13:00 h y 15:00 h, teniendo el monocultivo un máximo de 34.3 ± 0.3 °C y la milpa 33.9 ± 0.2 °C (Figura 1G).

Figura 1 Microclima en el área de cultivo de maíz bajo dos tipos de manejo establecidos en dos sitios al sureste de México (Peto: A-E y Mama: F-J), a los 75 días después de la siembra de los cultivos. M: monocultivo de maíz; MFC: combinación maíz-frijol-calabaza; PAR: Radiación fotosintéticamente activa; CVA: Contenido volumétrico de agua en el suelo; DPV: Déficit de presión de vapor de agua atmosférico. Los datos son promedios de tres días de medición.
La temperatura del suelo (Figura 1 C, H), no presentó diferencias significativas entre las temperaturas mínimas y máximas entre tipos de manejo y/o sitios de trabajo; sin embargo, la temperatura promedio del suelo en el monocultivo en Peto fue mayor a MFC (hasta 3.35 ºC mayor respecto a MFC; F = 15.29; P= 0.0001); en Mama no se registraron diferencias significativas en la temperatura mínima, máxima y promedio del suelo entre los tipos de manejo. El monocultivo presentó temperaturas hasta en 5 °C mayores respecto a MFC a las 15:00 h en Peto, y hasta 2 °C a las 13:00 h en Mama (Figura 1C, H).
El contenido volumétrico de agua en el suelo (CVA) promedio registró los valores más altos en el tratamiento MFC en Peto (0.3 ± 0.01 m3/m3), respecto al resto de tratamientos (F = 8.028; P = 0.0049). El curso diario del CVA (Figura 1 D, I) en el tratamiento MFC fue hasta 22 % mayor con respecto a M (Figura 1D) en Peto, mientras que en Mama (Figura 1I) el tratamiento M se mantuvo en el mismo valor (0.25 m3/m3) a lo largo del día, a diferencia de MFC que disminuyó a partir de las 11:00 h.
En el caso del DPV, en Mama se registraron los valores máximos más bajos, siendo hasta 0.82 kPa más bajos que en Peto (F = 22.44; P = 0.0002). A lo largo del día el DPV en Peto fue similar entre tratamientos (Figura 1E), mientras que en Mama el tratamiento M presentó un DPV 18 % mayor que MFC entre las 12:00 h y 16:00 h (Figura 1J).
Respuesta fisiológica del maíz. Durante el curso diario de A, se observó que las plantas bajo el tratamiento MFC en Peto registraron valores mayores respecto a las plantas del tratamiento M en ambos días de medición (Figura 2). La tasa de asimilación de CO2 total diaria (Atot) en las plantas de maíz en Peto y en Mama disminuyó a los 75 días en comparación con el día 40 del inicio de la siembra (F = 254.97; P < 0.05 y F = 189.28; P < 0.05, respectivamente); las plantas establecidas en ambos tratamientos en Mama presentaron mayor Atot que los tratamientos de Peto durante la primera y segunda medición (F = 100.75; P < 0.05 y F = 34.95; P < 0.5, respectivamente; Figura 2). En los cultivos de Peto, Atot fue igual entre M y MFC en ambas mediciones (Figura 2A, B). Para las plantas establecidas en Mama, Atot fue mayor en el día 40 respecto al día 75 (F = 189.28; P < 0.05), y las plantas bajo el tratamiento M presentaron mayor Atot que las plantas en el tratamiento MFC, en ambas mediciones (F = 5.32; P = 0.0258; Figura 2C, D).
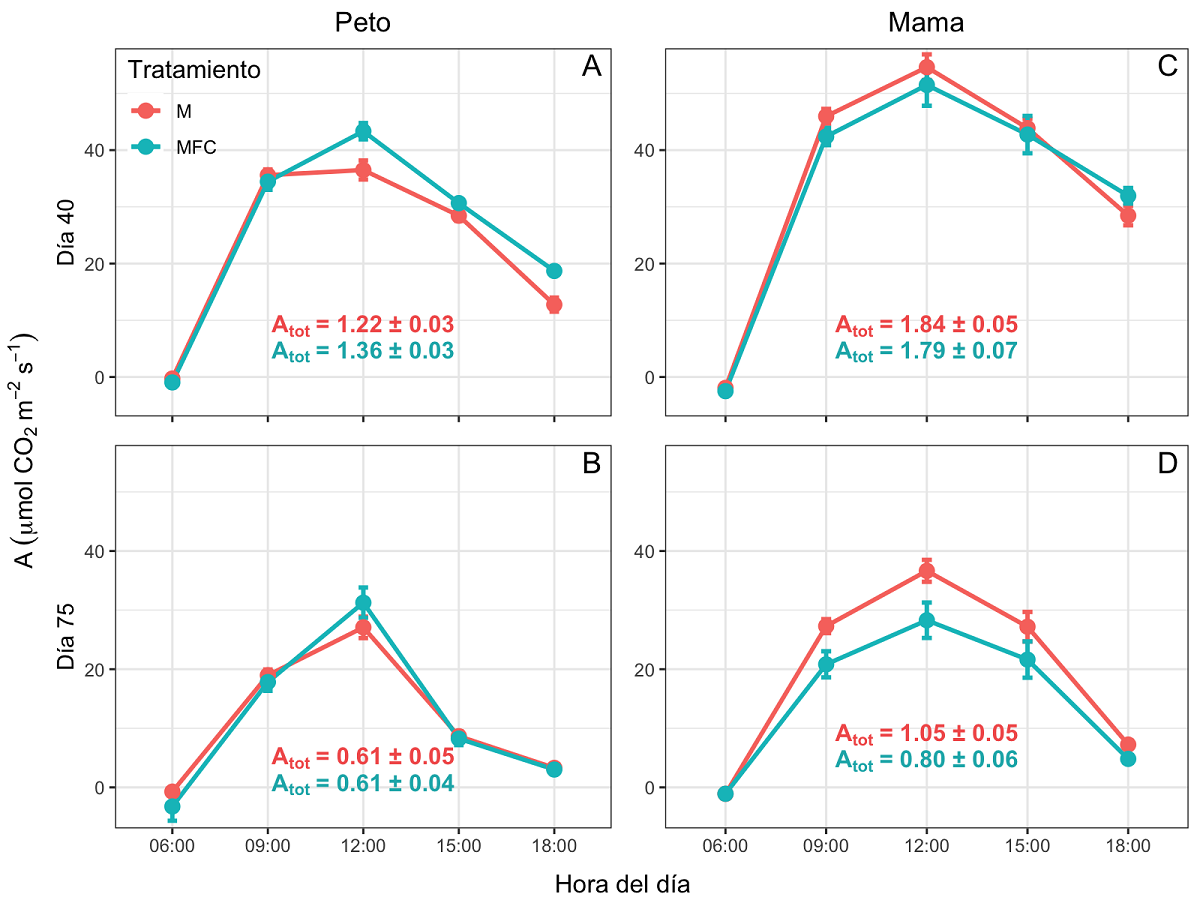
Figura 2 Curso diario de asimilación de CO2 (A; líneas y puntos) y promedio total diario (Atot; mol m-2d-1) en plantas de maíz bajo dos sistemas de manejo contrastantes y dos condiciones de precipitación al sur de México (Peto: A-B y Mama: C-D). M: monocultivo de maíz; MFC: combinación de maíz-frijol-calabaza. Los datos son promedios ± EE.
Los cursos diarios de transpiración (E), mostraron que las plantas establecidas en Peto presentaron mayor E a lo largo del día en el tratamiento MFC respecto al monocultivo, mientras que en Mama se observó lo contrario. Las tasas de transpiración totales diarias (Etot) en las plantas de ambos sitios y tratamientos disminuyeron entre la primera y segunda medición (F = 132.9; P < 0.05; Figura 3); las plantas establecidas en Mama presentaron mayor Etot en comparación con las establecidas en Peto (F = 116.72; P < 0.05), con la única excepción de las plantas en el tratamiento MFC al día 75. En Mama se registraron diferencias significativas en la segunda medición en donde las plantas bajo el tratamiento M presentaron una Etot 28.9 % superior a lo registrado en las plantas bajo el tratamiento MFC (F = 4.45; P = 0.0376; Figura 3D).

Figura 3 Curso diario de transpiración (E; líneas y puntos) y promedio total diario (Etot; mol m-2d-1) en plantas de maíz bajo dos sistemas de manejo contrastantes y dos condiciones de precipitación al sur de México (Peto: A-B y Mama: C-D). M: monocultivo de maíz; MFC: combinación de maíz-frijol-calabaza. Los datos son promedios ± EE.
Se observó que la eficiencia en el uso del agua (EUA) durante el día 40 fue mayor en las plantas bajo el tratamiento MFC hasta antes de las 15:00 h en Peto (Figura 4A), mientas que en Mama los registros fueron similares (Figura 4C); al día 75, las plantas en el monocultivo de Peto presentaron mayor EUA la mayor parte del día, mientras que en Mama se observó lo contrario (Figura 4B, D). La EUA total diaria (EUAtot) fue menor en ambos sitios en la segunda medición respecto a la primera (F = 56.81; P < 0.05; Fig. 4). La mayor EUAtot del experimento se registró en plantas del tratamiento M en Peto a los 40 días (6.15 ± 0.11 mmol CO2 / mol H2O m-2 d-1; F = 8.23; P = 0.0052). No se registraron diferencias entre tratamientos por día de medición en ninguno de los dos sitios de trabajo.

Figura 4 Curso diario de la eficiencia en el uso del agua (EUA, líneas y puntos) y promedio diario (EUAp; mmol CO2 m-2 d-1/ mol H2O m-2 d-1) en plantas de maíz bajo dos sistemas de manejo contrastantes y dos condiciones de precipitación al sur de México (Peto: A-B y Mama: C-D). M: monocultivo de maíz; MFC: combinación de maíz-frijol-calabaza. Los datos son promedios ± EE.
La conductancia estomática (Figura 5) fue mayor en las plantas del tratamiento MFC a lo largo del día, en comparación con las de M en ambos días de medición; en Mama se observó lo contario, con la excepción del día 40 entre las 15:00 y 18:00 h, cuando las plantas de MFC presentaron mayor conductancia que las de M.

Figura 5 Curso diario de la conductancia estomática (gs) en plantas de maíz bajo dos sistemas de manejo contrastantes y dos condiciones de precipitación al sur de México (Peto: A-B y Mama: C-D). M: monocultivo de maíz; MFC: combinación de maíz-frijol-calabaza. Los datos son promedios ± EE.
El Fv/Fm no presentó diferencias al día 40 de medición, mientras que al día 75 fue mayor en plantas bajo el tratamiento MFC respecto a las de M establecidas en Peto (F = 18.62; P < 0.05); en Mama no se encontraron diferencias entre tratamientos (Tabla 1). La eficiencia cuántica del fotosistema dos en plantas adaptadas a la luz (𝛷PSII) durante el periodo del día con mayor estrés lumínico no presentó diferencias significativas entre sitios, días de medición y tratamientos (Tabla 1), con la única excepción de las plantas bajo el tratamiento MFC de Mama en la segunda medición que fue 53.5 % menor que las plantas de M (F = 21.66; P < 0.05).
Tabla 1 Fluorescencia de la clorofila de las plantas de maíz registrados preamanecer (Fv/Fm) y entre las 12:00 h y 15:00 h (𝛷PSII, TTE y DNE) a los 40 y 75 días después de la siembra. M: monocultivo de maíz; MFC: combinación de maíz-frijol-calabaza; Fv/Fm: eficiencia cuática del fotosistema II en hojas adaptadas a la oscuridad; 𝛷PSII: eficiencia cuántica del fotosistema II en hojas adaptadas a la luz; TTE: tasa de transporte de electrones (µmol m-2 s-1); DNE: disipación no fotoquímica de energía. Los datos son promedios ± EE. Letras diferentes indican diferencias significativas entre sitios, día de medición y tratamientos (α = 0.05).
| Sitio | Día | Tratamiento | Fv/Fm | 𝛷PSII | TTE | DNE |
|---|---|---|---|---|---|---|
| Peto | 40 | M | 0.776 ±0.004b | 0.437 ± 0.02ab | 210.59 ± 10.49ª | 1.01 ± 0.14bc |
| MFC | 0.785 ± 0.001ab | 0.469 ± 0.02a | 235.87 ± 14.27ª | 0.83 ± 0.11c | ||
| 75 | M | 0.752 ± 0.004c | 0.396 ± 0.04ab | 121.92 ± 11.94b | 1.05 ± 0.17bc | |
| MFC | 0.773 ± 0.003b | 0.409 ± 0.04ab | 133.07 ± 14.98b | 0.80 ± 0.11c | ||
| Mama | 40 | M | 0.776 ± 0.004b | 0.363 ± 0.01ab | 291.11 ± 11.23ª | 1.36 ± 0.05ab |
| MFC | 0.792 ± 0.002a | 0.360 ± 0.01ab | 288.58 ± 14.19ª | 1.38 ± 0.06ab | ||
| 75 | M | 0.772 ± 0.003b | 0.302 ± 0.01b | 217.17 ± 10.98ª | 1.60 ± 0.05a | |
| MFC | 0.779 ± 0.003b | 0.219 ± 0.02c | 156.93 ± 15.38b | 1.53 ± 0.04a |
En el caso de la TTE (Tabla 1), los valores más altos observados entre las 12:00 h y 15:00 h se registraron en la primera medición respecto a la segunda (F = 93.86, P < 0.05); las plantas establecidas en Peto no presentaron diferencias entre tratamientos, mientras que en Mama las plantas del tratamiento M presentaron un valor hasta 46.4 % mayor que las de MFC en la medición del día 75 (F = 4.051, P = 0.04560; Tabla 2).
Tabla 2 Contenido de clorofila y estado hídrico foliar en plantas de maíz bajo dos manejos de cultivo establecidos en dos sitios con pedregosidad y profundidad del suelo contrastantes. M: monocultivo de maíz; MFC: combinación de maíz-frijol-calabaza; CC: Contenido de clorofoila foliar ; CRA: Contenido relativo de agua foliar; Ψ: Potencial hídrico. Los datos son promedios ± EE. Letras diferentes indican diferencias significativas entre sitios, día de medición y tratamientos (α = 0.05).
| Sitio | Día | Trat | CC (μmol m-2) | CRA (%) | Ψ (MPa) |
|---|---|---|---|---|---|
| Peto | 40 | M | 275.15 ± 12.47cd | 81.91 ± 2.23cd | -0.93 ± 0.08a |
| MFC | 310.71 ± 17.39c | 85.48 ± 1.06bcd | -1.14 ± 0.09ab | ||
| 75 | M | 250.53 ± 7.31d | 76.72 ± 4.74d | -1.60 ± 0.10c | |
| MFC | 347.88 ± 23.47bc | 90.86 ± 1.30abc | -1.39 ± 0.08bc | ||
| Mama | 40 | M | 433.46 ± 12.39a | 96.21 ± 0.62a | -0.95 ± 0.07a |
| MFC | 434.00 ± 14.67a | 91.04 ± 2.04ab | -1.06 ± 0.06ab | ||
| 75 | M | 418.52 ± 23.02ab | 73.12 ± 2.60d | -1.25 ± 0.10abc | |
| MFC | 449.79 ± 15.86a | 82.18 ± 1.88cd | -1.23 ± 0.03ab |
La DNE entre las 12:00 h y 15:00 h (Tabla 1) presentó diferencias significativas entre sitios, donde las plantas establecidas en ambos tratamientos en Mama presentaron valores más altos que los de Peto en ambas mediciones (F= 52.86; P < 0.05). No se registraron diferencias entre tratamientos y tiempo de medición en ninguno de los dos sitios.
Los cursos diarios mostraron que al día 75 en Peto la 𝛷PSII y la TTE fue similar en plantas de ambos tratamientos a lo largo del día, mientras que en Mama fueron superiores en las plantas de M respecto a las de MFC (Material Suplementario Figura S2 A, B, D, E). En las plantas en Mama no presentaron diferencias entre tratamientos, mientras que en Peto se registraron valores más altos en las plantas del monocultivo (Material Suplementario Figura S2 C, F).
El contenido de clorofila (CC) fue menor en las plantas bajo ambos tratamientos en Peto, respecto a Mama (F = 137.39; P < 0.05; Tabla 2). En Mama no se registraron diferencias entre tratamientos en ambas mediciones, mientras que en Peto, únicamente se registraron diferencias al día 75 cuando las plantas de MFC presentaron un CC 27.9 % mayor que las de M.
El contenido relativo de agua de las plantas (CRA, Tabla 2) se redujo a los 75 días de medición en Mama, respecto al día 40 en ambos tratamientos (F = 22.78; P < 0.05); no se registraron diferencias entre tratamientos por día de medición. En Peto se registraron diferencias en las plantas del tratamiento M que presentaron una reducción al día 75 respecto al día 40 (F = 7.21; P = 0.008; Tabla 2).
Se observó una interacción significativa entre tratamientos y día de medición (F = 5.621; P = 0.019) para el potencial hídrico foliar (Ψ; Tabla 2); los registros fueron más negativos al día 75, respecto al día 40 en los tratamientos de sitios. En Peto, no se registraron diferencias entre tratamientos por día de medición, sin embargo, las plantas en el tratamiento M presentaron valores más negativos al día 75. En Mama no se encontraron diferencias significativas entre tratamientos por día de medición.
Las plantas establecidas en ambos tratamientos en Mama presentaron mayor altura que las del sitio de Peto en ambas mediciones (F = 103.51; P < 0.05; Tabla 3). No se registraron diferencias significativas entre tratamientos en un mismo sitio y día de medición. Tampoco se registraron diferencias significativas en el peso de las mazorcas enteras (1,825 g vs 1,925 g para M y MFC, respectivamente), ni en el peso desgranado (1,506.7 g vs 1,553.38 g para M y MFC, respectivamente).
Tabla 3 Altura de plantas de maíz bajo dos sistemas de manejo establecidos en dos sitios con condiciones de pedregosidad y profundidad del suelo contrastantes. M: monocultivo de maíz; MFC: tratamiento de maíz-frijol-calabaza. Los datos son promedios ± EE. Letras diferentes indican diferencias significativas entre sitios, día de medición y tratamientos (α = 0.05).
| Sitio | Día | Tratamiento | Altura (cm) |
|---|---|---|---|
| Peto | 40 | M | 78.66 ± 3.57d |
| MFC | 93.66 ± 3.86d | ||
| 75 | M | 215.83 ± 2.96b | |
| MFC | 233.50 ± 4.59b | ||
| Mama | 40 | M | 113.16 ± 4.24c |
| MFC | 118.75 ± 5.67c | ||
| 75 | M | 252.58 ± 3.23a | |
| MFC | 256.50 ± 4.38a |
El análisis NMDS (Figura 6) sugiere que las plantas de maíz tuvieron comportamientos fisiológicos contrastantes entre sitios; dentro de cada sitio, el desempeño fisiológico entre plantas bajo ambos tratamientos fue más parecido en Mama (nuestro sitio con menor pedregosidad y suelos más profundos) en comparación con las plantas establecidas en Peto. En este último sitio, el comportamiento fisiológico de las plantas de maíz en el tratamiento MFC mostró una mayor amplitud de respuesta que las plantas en el tratamiento M, la cual se asoció más al contenido volumétrico de agua en suelo y al DPV mínimo, mientras que en las plantas del tratamiento M se asociaron en mayor medida a una mayor temperatura del suelo. El análisis también mostró que la respuesta fisiológica de las plantas de ambos tratamientos en Peto se asoció a una alta pedregosidad y baja profundidad del suelo, mientras que en Mamá se observó lo contrario.
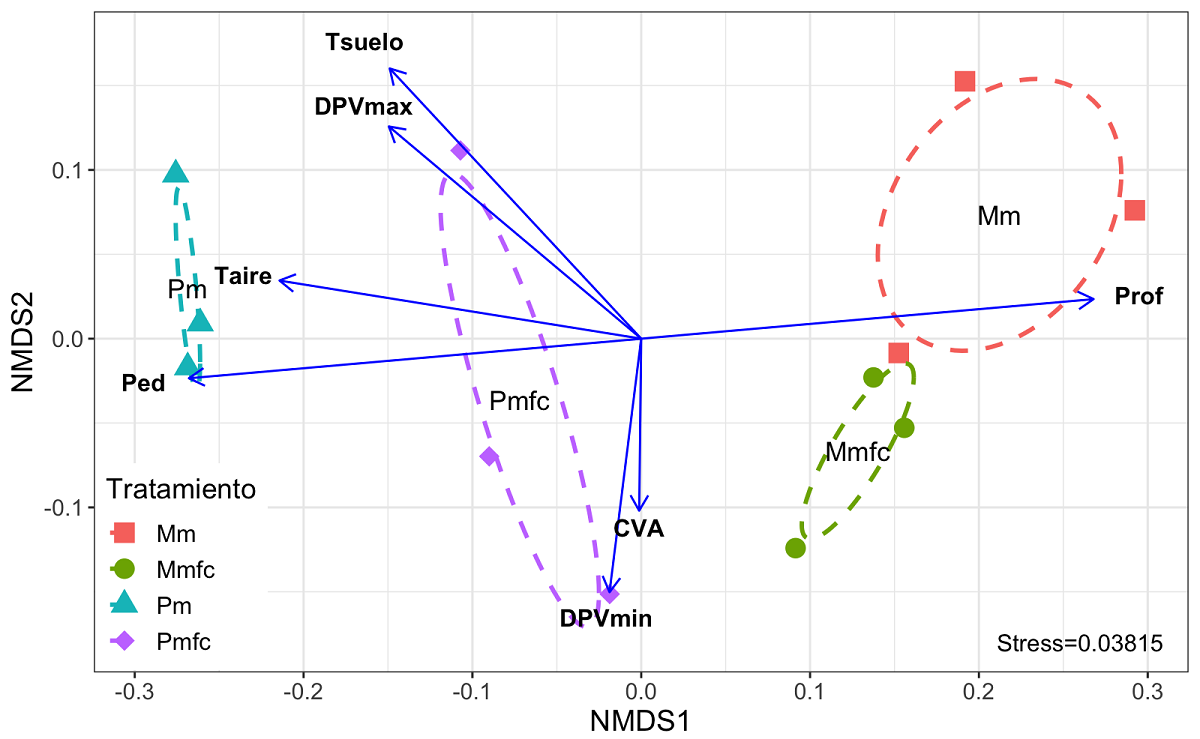
Figura 6 Análisis no métrico de escala multidimensional construido a partir de las variables fisiológicas medidas en plantas de maíz bajo dos tratamientos (monocultivo y combinación maíz-frijol-calabaza) establecidos en dos sitios con pedregosidad y profundidad del suelo (Peto y Mama, Yucatán), y su relación con las variables microclimáticas medidas en dichos sitios al día 75 contado a partir de la germinación del maíz. Mm: monocultivo en Mama; Mmfc: maíz-frijol-calabaza en Mama; Pm: monocultivo en Peto; Pmfc: maíz-frijol-calabaza en Peto. Tsuelo: temperatura del suelo; Taire: temperatura del aire; DPVmax y DPVmin; déficit de presión de vapor de agua máximo y mínimo, respectivamente; CVA: contenido volumétrico de agua en suelo. Las líneas punteadas indican el grado de dispersión de los datos para cada tratamiento y sitio de medición, construidos a partir de una matriz de covarianza de la salida de datos del NMDS.
Discusión
Nuestro estudio encontró que la relación entre el sistema de policultivo de la milpa (MFC) y el microambiente es complejo, ya que sólo en el caso del sitio de estudio con el suelo pedregoso y poco profundo (Peto), la milpa representó una ventaja al crecimiento del maíz. Siendo que, bajo condiciones de crecimiento más favorables, con suelos más profundos (Mama), el maíz registró mayores tasas de asimilación en monocultivo por lo que nuestras hipótesis solo se cumplieron parcialmente. Estudios ecológicos han encontrado que el fenómeno de la facilitación entre plantas complementarias se hace más frecuente en sitios más estresantes, mientras que en sitios más favorables se favorece la competencia (Callaway et al. 2002, Maestre et al. 2009). En este caso, el maíz, como planta C4 de crecimiento rápido, es la especie más competitiva y que más se beneficia de las condiciones favorables.
Las plantas de maíz en Mama presentaron mayores tasas de asimilación, mayor altura e incluso las mayores tasas de germinación comparado con Peto, lo cual pudo deberse a la mayor profundidad, menor pedregosidad y manejo del suelo en esta localidad. Lafitte (1993) menciona que los suelos poco profundos con poca capacidad de retención de agua son una de las principales causas de estrés hídrico sobre las plantas de maíz, limitando la germinación de las semillas y desarrollo óptimo de las plantas. Aunado a esto, Terán-Contreras & Rasmussen (2009), aseguran que en Yucatán los suelos delgados, permeables y pedregosos representan una de las limitantes ecológicas más importantes para el sistema milpa. Por otro lado, en Mama se registró el uso de Mucuna como abono verde durante los periodos inter-cultivos, lo que contribuye a la conservación del agua en el suelo y a la disponibilidad de nutrientes (Altieri & Nicholls 2013). Incluso, el sistema maíz-mucuna ha resultado más productivo que el sistema roza-tumba-quema a nivel de unidad familiar (Guevara et al. 2000). En su conjunto, estas características favorecieron en Mama mejores condiciones edáficas que fueron aprovechados por el maíz.
En el caso de Peto, la alta pedregosidad limitó la fotosíntesis y la transpiración del maíz, pero la disminución en estas variables fisiológicas fue menor en la milpa (MFC), lo cual fue acompañado por ventajas microclimáticas en el sistema de manejo MFC, en comparación con M en el mismo sitio. En Peto, MFC presentó una menor temperatura del suelo y mayor contenido volumétrico de agua, que M, lo que compensó la similitud en otros elementos climáticos como el DPV: al contar con mayor agua en el suelo, las plantas en el tratamiento MFC podrían compensar de mejor forma la pérdida de agua por transpiración en comparación con las de M. Gliessman (1985) explica que la cobertura vegetal que provee la diversidad de especies que componen los sistemas de manejo con cultivos múltiples permite una menor exposición a la radiación solar, lo que contribuye a la reducción de la temperatura del suelo y, por ende, a una menor evaporación del agua del suelo. Dado que la lluvia y la radiación solar son factores clave que afectan directamente la humedad del suelo (Vivoni et al. 2010), en suelos pedregosos y superficiales la humedad no puede ser almacenada en el suelo por tiempos prolongados, particularmente si la demanda evaporativa es grande, lo cual significó que los beneficios de dicha cobertura fueran más significativos en Peto.
Respecto a la reducción en las tasas de asimilación de CO2 y transpiración hacia el final del experimento, Geiger (2009) explica que una vez que cesa el crecimiento vegetativo, las plantas de maíz tienen una caída en las tasas fotosintéticas y de transpiración. De igual forma, las altas temperaturas registradas en Peto (superiores a los 32 ºC) pudieron contribuir a la reducción en la tasa de asimilación. Lo anterior coincide con lo reportado por Montoro & Ruíz (2017), quienes afirman que en algunos maíces, las tasas de fotosíntesis tienden a disminuir conforme aumenta la temperatura del aire, principalmente en un rango de 32.5 a 45 °C. En este sentido, los tratamientos de Mama tuvieron una temperatura promedio hasta 2 °C menos que los de Peto, lo que podría explicar por qué los tratamientos del primer sitio presentaron las tasas de asimilación de CO2 mayores a las del segundo.
Tomando en cuenta la relación entre fotosíntesis, transpiración y contenido de agua en suelo, el hecho de que los dos tratamientos en Peto tuvieron la misma asimilación de CO2 y tasa de transpiración hacia el final del experimento, podría explicarse por las altas temperaturas registradas en el sitio, lo que podría propiciar el cierre de estomas limitando el intercambio gaseoso. Si la cantidad de agua en el suelo es suficiente, las plantas no tienen necesidad de retener el agua, por ello los estomas se mantienen abiertos favoreciendo al intercambio gaseoso, aunque esto implique una mayor transpiración (Yu et al. 2016, Montoro & Ruiz 2017). Por ello, debido a que MFC contó con más agua en el suelo, la mayor conductancia estomática registrada permitió una mayor transpiración a diferencia de las plantas del tratamiento M. En este sentido nuestros resultados coinciden con los de Pérez-Hernández et al. (2021), las plantas de maíz en su tratamiento maíz-frijol y maíz-frijol-calabaza tuvieron una mayor tasa de asimilación (0.81 ± 0.05 y 0.76 ± 0.07 mol CO2 m-2 d-1 respectivamente) respecto al monocultivo, así como de transpiración, ya que estos tratamientos tenían un mayor contenido de agua en el suelo (maíz-frijol: 0.27 ± 0.001 m3/m3; maíz-frijol-calabaza: 0.33 ± 0.001 m3/m3) que los registrados en nuestro experimento.
Contrario a lo reportado por Pérez-Hernández et al. (2021), quienes no encontraron diferencias en la EUA entre plantas de maíz establecidas en un monocultivo respecto a otras establecidas bajo el sistema milpa, las diferencias registradas en nuestro caso concuerdan con lo reportado por Gliessman (1985), quien asegura que los cultivos múltiples usan de manera más eficiente el agua que los sistemas en monocultivo; sin embargo, en nuestro caso esto dependió de la pedregosidad y profundidad del suelo: el policultivo fue más eficiente en suelos pedregosos y superficiales, mientras que el monocultivo lo fue en sitios con suelos más profundos y menos pedregosos hacia el final del experimento. En este sentido, Conceição dos Santos et al. (2019), rescatan que las plantas adaptadas a condiciones de sequía suelen presentar mayores valores de EUA, lo cual resalta la importancia del uso de variedades nativas adaptadas a las condiciones locales, como Nal Xoy blanco en nuestro caso, el cual está adaptado a las condiciones del suelo descritas, además de tolerar el estrés hídrico.
Las mayores temperaturas del día 75, la baja precipitación (180 y 155 mm entre el día 40 y el 75 en Peto y Mama, respectivamente) y posiblemente la mayor competencia por agua al tener las plantas tallas más grandes, causaron una disminución del CRA y el potencial hídrico de las hojas de maíz con respecto a la primera medición. El potencial hídrico fue menor en Peto, aun cuando el CRA fue similar en ambos sitios. Esta diferencia podría explicarse por un ajuste osmótico (Blum 2017), o una acumulación de osmolitos que permite bajar el potencial hídrico para obtener agua del suelo, lo cual podría ser más difícil en los suelos pedregosos de Peto.
Adicionalmente, la condensación de rocío en las hojas resulta importante como una fuente de entrada de agua al sistema, en Mama documentamos este fenómeno con una duración de hasta cuatro horas más en las dos mediciones respecto a Peto (Material suplementario Figura S3). Según Paliwal et al. (2001), la cantidad de rocío que se condensa sobre las hojas y cae en la base de las plantas, puede ser una de las fuentes principales de agua en el cultivo del maíz, además de la lluvia, (Material suplementario Figura S5). Un ejemplo de lo anterior lo reportan Pérez-Hernández et al. (2021), donde la precipitación fue baja, pero gracias a una alta humedad relativa (80 %), hubo condensación de agua sobre las hojas de las plantas de maíz por 12 h, lo que contribuyó a humedecer el suelo.
Finalmente, el NMDS mostró un claro efecto del sistema de manejo sobre el desempeño fisiológico del maíz, donde se resalta la conservación de agua en suelo, la menor temperatura del suelo y aire en comparación con el monocultivo. La diferenciación en las estrategias fisiológicas seguidas por las plantas de maíz bajo ambos tratamientos es marcado bajo ambas condiciones de precipitación, a pesar de que algo similar fue reportado por Pérez-Hernández et al. (2021) bajo tratamientos similares, en su caso dicha diferenciación en el desempeño fisiológico del maíz no fue tan marcado como en nuestro caso, probablemente debido a que el experimento de estos autores se llevó a cabo en un sitio con una precipitación, humedad relativa del aire y profundidad del suelo mucho mayores que en nuestro estudio.
De acuerdo con los resultados obtenidos, podemos apoyar la premisa de que los diferentes sistemas de manejo de un cultivo pueden afectar las condiciones microclimáticas y comportamiento fisiológico de las plantas que lo componen. Si bien las ventajas fisiológias del policultivo respecto al monocultivo dependen de las condiciones del sitio de establecimiento (particularmente del suelo), consideramos una mejor alternativa el sistema milpa en todos los casos, ya que además de la obtención de maíz (foco de este trabajo), también se pueden obtener una diversidad de alimentos dentro del mismo espacio de terreno de cultivo.
La profundidad, baja pedregosidad y manejo previo del suelo en Mama, fueron factores que influyeron para que la asimilación de CO2 fuera mayor y que las plantas fueran más grandes respecto a las de Peto.
Por último, los resultados obtenidos de esta investigación demuestran la importancia del sistema de manejo como un mecanismo que contribuya a proveer mejores condiciones microclimáticas para el desempeño fisiológico del maíz, particularmente en sitios de alta pedregosidad, suelos delgados, y relativamente secos como en Peto.
Material suplementario
El material suplementario de este artículo puede encontrarse aquí: https://doi.org/10.17129/botsci.3365

