











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Tropical montane cloud forests (TMCF) are ecosystems where tree crowns condense atmospheric water because they are regularly in contact with dense fog, generating a phenomenon known as cloud stripping or horizontal precipitation that supports the development of local vegetation (Bruijnzeel & Proctor 1995). These forests distribute across tropical Africa, America and Asia and, despite their reduced cover (less than 1 % of the continental Earth’s surface), preserve them is a worldwide priority because they concentrate several endemic species and provide a number of ecosystem services, including regulation of hydrological cycles, carbon sequestration and prevention of soil erosion (Doumenge et al. 1995, Bruijnzeel et al. 2011). Notwithstanding the ecological value of these ecosystems, TMCF have been historically affected by extensive land use changes and their functional integrity is strongly threatened (Karger et al. 2021).
In Mexico, TMCF distribute over the major mountain ranges of this country and harbor 6,163 native vascular plants and a highly diverse fauna-551 birds, 257 mammals, 249 reptiles and 138 amphibians (Gual-Díaz 2014). These forests originally covered about 20,000 km2 (Rzedowski 1978), but more than 50 % of their surface have been replaced by grasslands and agricultural fields during the past century (Challenger & Soberón 2008). These activities, however, in some places have progressively ceased over the last three decades due to the increasing emigration of rural population towards urbanized areas (Sanchez & Pacheco 2012) and the Mexican government has implemented public policies addressed to restore TMCF (Ramírez-Soto et al. 2018, Trujillo-Miranda et al. 2018, Toledo-Aceves et al. 2021). These policies promote the transplanting of saplings of mid- and late-successional native trees in open areas or fallow because they are aimed in recovering ecosystem services as soon as possible, but this strategy usually fails because saplings do not tolerate the elevated solar exposure, high temperatures and water shortage conditions that prevail in these sites (Camacho-Cruz et al. 2000, Ramírez-Bamonde et al. 2005, Ramírez-Marcial et al. 2006). To cope with these limitations, it was proposed that the early introduction of native pioneer trees in degraded areas can ameliorate these harsh environmental conditions and, thereby, they can act as nurse plants that facilitate the establishment of species belonging to advanced successional stages (Padilla & Pugnaire 2006, Avendaño-Yáñez et al. 2014).
Successional inter-specific interactions have been described with three mechanisms: facilitation, inhibition and tolerance (Connell & Slatyer 1977). The facilitation mechanism seeks to explain processes by which particular (pioneer) species generate more favorable microenvironments for the establishment of other species, mostly of advanced successional stages. The tolerance mechanism stands for simultaneous establishment of both early and late species, while the latter presents higher recruitment rates in advanced successional stages. Finally, inhibition is when species from different successional stages establish but one species inhibits the germination and/or establishment of other species. In this case, the replacement of species occurs only when the initial species die. In tropical forests facilitation and tolerance mechanisms occur simultaneously (Saldaña-Acosta 2001, Pineda-López et al. 2000).
These positive plant-plant interactions are commonly due to the provisioning of shaded microhabitats beneath nurse canopies, where temperature is lower and soil moisture is higher than in areas without vegetation cover (Callaway 1992, Suzán-Azpiri & Sosa 2006, Padilla & Pugnaire 2009, Badano et al. 2016). Nevertheless, these microenvironmental conditions are often distributed heterogeneously beneath nurses because the amount of solar radiation intercepted by their canopies varies among cardinal directions - e.g., in the Northern Hemisphere, south-facing sides of plant canopies receive more solar radiation than north-facing ones, while the converse occurs in a desertified alpine meadow in the Southern Hemisphere (Liou 2002) and in Mediterranean pastures in North Hemisphere (López-Pintor et al. 2006). As these microenvironmental variations affect the performance of beneficiary plants (Franco & Nobel 1989, Drezner & Garrity 2003). The using positive plant-plant interactions to restore TMCF requires determining the cardinal direction on which these species must be transplanted to increase their establishment rates beneath nurses.
Additionally, in order to stimulate successional processes in ecosystems, ecological interactions must be considered. For example, the magnitude of positive plant - plant interaction on beneficiary species is positively related to their crown cover. Another issue that must be considered before employing positive plant-plant interactions to restore TMCF is that the magnitude of the positive effects that nurses exert on beneficiary species relates positively with the cover of their canopies (Tewksbury & Lloyd 2001, Peláez et al. 2019, Liu et al. 2020). Thus, using facilitation as restoration strategy may be a time-consuming process if crowns of pioneer trees transplanted in degraded areas require reaching some minimum size to facilitate late-successional species and, for this reason, these actions should be implemented with fast-growing nurses. Further, even when nurses enhance the recruitment rates of other species, it was widely documented that they can decrease the growth rates of beneficiary plants because of aboveground competition for light and/or belowground competition for water and mineral nutrients (Franco & Nobel 1989, Walker et al. 2001, Endo et al. 2008, Prieto et al. 2011). However, since TMCF late successional tree have extremely low establishment rates on fallow land, some authors suggested that nurse plants are the most viable option to restore them and recover the ecosystem services they provide (Aide et al. 2011, Avendaño-Yáñez et al. 2014).
Considering these issues, we propose that success in establishing late-successional species on fallow can be accelerated through the joint transplanting of fast-growing pioneer trees together with saplings of mid- and late-successional species. We hypothesized that this strategy, to be effective, requires transplanting mid- and late-successional species in the close neighborhood of putative nurses, so as the quick canopy development of the latter species should facilitate the establishment of the former ones in the short term. If so, the magnitude with which nurses enhance the performance of mid- and late-successional trees should increase with the cover their canopies despite the competitive effects that might arise between the benefactor and the beneficiary species. To test this proposal, we conducted a two-year field experiment addressed to answer the following questions: (1) do pioneer trees actually facilitate the survivorship and growth of mid- and late-successional species in a fallow? (2) does the establishment of mid- and late-successional trees vary according to the cardinal direction on which they are transplanted beneath nurses? (3) are there competitive effects between nurses and beneficiary species? and (4) is the magnitude of these positive/negative interspecific interactions affected by the canopy cover of nurses?
Materials and methods
To test our predictions, we selected a fallow land of 5 ha in Cerro La Silleta (21° 27' 22.74" N, 99° 3' 50.79" W, 1,373 m), a communal protected area (i.e., local inhabitants protect it to promote ecotourism) at the municipality of Xilitla, state of San Luis Potosí, Mexico. Mean annual temperature in this region is 24 °C and mean annual precipitation is 2,152 mm, but temperature can surpass 30 °C in summer and 72 % of rainfall concentrates in this season (Medina-García et al. 2005). Although local TMCF have been fragmented due to the opening of clearings for crop and livestock farming, there are well-preserved forest parches that contain remnants of the native vegetation (Reyes-Hernández et al. 2017). These forest patches are dominated by Heliocarpus appendiculatus Turcz (Malvaceae), Quercus germana, Quercus acutifolia Née (Fagaceae), Oreopanax xalapensis Decne. & Planch. (Araliaceae), Trema micrantha (L.) Blume (Cannabaceae), Ulmus mexicana Planch. (Ulmaceae), Wimmeria concolor Schltdl. & Cham. (Celastraceae), Dendropanax arboreus (L.) Decne. & Planch. (Araliaceae), Magnolia dealbata Zucc., Magnolia schiedeana (Magnoliaceae), Morus celtidifolia Kunth (Moraceae), Zanthoxylum caribaeum Lam. (Rutaceae), Clethra pringlei S. Watson (Clethraceae), Colubrina greggii S. Watson (Rhamnaceae), Gymnanthes longipes Müll.Arg. (Euphorbiaceae) and Liquidambar styraciflua (Altingiaceae) (Rzedowski 1966, Fortanelli-Martínez et al. 2014).
For this study, we selected a tree species. Liquidambar styraciflua was selected as the pioneer nurse tree species because it is widely distributed in disturbed cloud forests, and has been used to accelerate restoration in cloud forests in Mexico (Pedraza &Williams-Linera 2003, Ramírez-Bamonde et al. 2005). Quercus germana and Magnolia schiedeana are late and intermediate species, and it has been used very abundant in cloud forest with high litter production (Ramírez-Bamonde et al. 2005). We focused on these species because they are produced in local nurseries for commercial purposes and, therefore, they are easily to acquire for their developing restoration proposals. The pioneer species, L. styraciflua, is a fast-growing tree with wide round canopies that distributes from southeast USA to Central America and, in Mexico, it is widely used to recover soils degraded by overgrazing or intensive crop farming (Pedraza & Williams-Linera 2003, Ramírez-Bamonde et al. 2005). The mid-successional tree, M. schiedeana, is endemic to TMCF of Mexico and their saplings have extremely low recruitment rates in deforested areas with elevated solar exposure, but it recruits successfully in sites where forests has been partially removed because of selective logging (Vásquez-Morales et al. 2017). The late-successional species, Q. germana, is also endemic to Mexican TMCF and it is a shade-tolerant tree that mainly recruit in forest understories (Guzmán-Guzmán & Williams-Linera 2006). In the nurseries, the saplings of L. styraciflua are developed from tree cuttings because this allows getting 1.4-1.7 m tall individuals in a year or less, while saplings of M. schiedeana and Q. germana are developed from seeds collected in the forests. For our restoration trial, we acquired one-year-old saplings of these species and measured their heights. On L. styraciflua, we also measured length of the major and minor axes of their canopies and estimated their cover as the area of an ellipse. All individuals of L. styraciflua were 1.5-1.6 m tall when acquired, but the cover of their canopies varied between 0.25 and 1.3 m2. Saplings of M. schiedeana and Q. germana, on the other hand, were 0.4-0.5 m tall when acquired. The individuals of all species were transplanted in the target site on 04 March 2016 without a previous acclimatization period following the experimental design described below. The current landscape is heavily fragmented, with anthropogenic disturbance due to crop and livestock farming (Reyes-Hernández et al. 2017). The land was previously occupied by TMCF and was cleared in 1956. Corn and bean crops were raised with fallow periods of three to five years until 1984. Subsequently, for a period of 30 years, it sustained the introduced pasture grass Cynodon nlemfuensis Vanderyst. which is cultivated for fodder. In 2014 the land was cleared, and at the start of the experiment, it had been fallowed for one year (personal communication with the owners of the land).
As the target site is located a gentle slope (~8° inclination), we used a clinometer (Omni-Slope, Brunton, USA) to draw two parallel contour lines (165 m length each) that were spaced 18 m between them. On each contour line, we transplanted a L. styraciflua sapling every 15 m (12 individuals per line, 24 individuals in total). After that, we dug planting holes 0.3 m away from the stems of these putative nurses on the four main cardinal directions - north (0°), east (90°), south (180°), and west (270°) - and randomly assigned them to saplings of M. schiedeana and Q. germana until transplanting 12 individuals of each species on each cardinal direction (Figure 1). The saplings of these mid- and late successional species were assumed to be nurse-protected in the short term. On all cardinal directions, we also dug planting holes 2 m away from the stems of L. styraciflua individuals and repeated the former transplanting procedure until getting 12 saplings of M. schiedeana and Q. germana that were not expected to be protected by nurses and they were used as controls (Figure 1). Neither watering nor fertilizers were applied after individuals were transplanted.

Figure 1 Schematic diagram of the planting design used to test the viability of using positive plant-plant interactions to restore Mexican TMCF. This design involved transplanting of saplings of a pioneer tree (Liquidambar styraciflua) in the target site and, after that, transplanting saplings of a mid-successional tree (Magnolia schiedeana) and a late-successional tree (Quercus germana) close to their stems (green area) and away from the area of influence of their canopies on the four main cardinal directions.
We returned to the target site on 09 November 2018 (31 months after the transplant of individuals). By this date, all L. styraciflua were fully established and expanded their canopies fully covering the transplant microsites located 0.3 m away from their stems, while all transplant microsites located 2 m away from their stems remained uncovered. We counted the number of M. schiedeana and Q. germana saplings that recruited beneath and outside the canopies of L. styraciflua on each cardinal direction. In all cases, we assumed that individuals whose leaves and stems were fully withered died over the last two years. This binary data (alive/dead saplings) was used to estimate and compare the establishment probabilities of the mid- and late-successional species between nurse-protected and unprotected individuals across cardinal directions. For each beneficiary species, we run a generalized linear model (GLM) based on logit function with binomial error distribution and full-factorial design, estimating the establishment probabilities as marginal means (Zuur et al. 2009). We used bootstrap without-replacement resampling routines to assess the accuracy with which GLM estimate and compare establishment probabilities of M. schiedeana and Q. germana. This procedure was repeated 1,000 times randomly selecting 75 % of the data run the GLM, while the remaining 25 % of the data was used to build receiver operating characteristic curves and calculating the area beneath them (AUC). The 1,000 AUC values were averaged and, as they range between 0 (zero) and 1 (one), the accuracy of GLM to estimate the establishment probabilities was assumed to increase as AUC approached to 1 (Fielding & Bell 1997).
After assessing the effects of L. styraciflua on the establishment of M. schiedeana and Q. germana, we focused in determining whether the putative nurse competes with these beneficiary species. For this, we measured the height of all surviving individuals of M. schiedeana and Q. germana and computed the difference between their initial and final height, estimating their net growth rates as the ratio between this difference and the time elapsed since the beginning of the restoration trial, which was equivalent to 2.68 years. Net annual growth rates of M. schiedeana and Q. germana were compared separately between nurse-protected and unprotected saplings across cardinal directions with two-way factorial ANOVA. These analyses were addressed to test the hypothesis that, if the nurse competes with the beneficiary species, annual growth rates of M. schiedeana and Q. germana saplings established beneath the canopy of L. styraciflua should be smaller than those of unprotected saplings.
We finally focused on the effects of nurse canopy cover on the performance of beneficiary species. The cover that canopies of L. styraciflua individuals reached after two years in the field was estimated as described above (i.e., measuring the length of their major and minor axes). This information was used to evaluate the effects of canopy cover on the establishment of M. schiedeana and Q. germana saplings with logistic regressions with categorical variables. These analyses were conducted separately of each species and only included saplings protected by nurse canopies. In this way, we estimated the recruitment probabilities of beneficiary species as a function of canopy cover and compared these relationships across cardinal directions, while the accuracy of regression models to estimate the response variable was assessed computing AUC values as described for GLM. We later focused on the effects of canopy cover on the annual growth rates of M. schiedeana and Q. germana established beneath the canopy of L. styraciflua. For this, we performed linear regressions with categorical variables to determine whether values of this variable relate with the canopy cover of nurses and whether these relationships differ across cardinal directions. In these two regression analyses, negative relationships should be obtained if competitive effects of the nurses on beneficiary species increase with canopy cover. All analyses described in this section were conducted in R 4.2 (R Core Team 2022).
Results
Establishment probabilities of M. schiedeana and Q. germana were estimated with elevated accuracy, as AUC values of GLM were above 0.7 for both species (Table 1). These analyses indicated higher establishment probabilities of M. schiedeana and Q. germana saplings beneath the canopies of L. styraciflua than outside them, but neither effects of the cardinal direction nor its interaction with nurses were found (Table 1, Figure 2).
Table 1 Results of statistical analyses used to determine whether Liquidambar styraciflua facilitates the establishment and growth of saplings of Magnolia schiedeana and Quercus germana, as well as to assess the effects of the canopy cover of nurses on these two variables. The table includes the results of the GLM comparing establishment probabilities between nurse-protected and unprotected saplings, and the results of the two-way factional ANOVA comparing annual growth rates between nurse-protected and unprotected saplings. These two analyses also assessed the effects of the cardinal direction on which saplings were transplanted and the effects of the interaction between levels of this variable and nurse protection. After that, the table shows the results of the logistic regressions with categorical variables used to determine whether increasing nurse canopy cover influences the establishment probabilities of beneficiary species, and the results of the linear regressions assessing whether variation in nurse canopy cover affects annual growth rates of beneficiary species. In both regression analyses, cardinal directions on which saplings were transplanted were included as levels of the categorical variable.
| Statistical analysis | Values of statistics | Magnolia schiedeana | Quercus germana |
|---|---|---|---|
| Results of GLM | AUC value of the GLM | AUC = 0.701 | AUC = 0.876 |
| Effect of the nurse plants | Wald - Ⲭ2 = 6.968, d.f. = 1, P = 0.008 | Wald - Ⲭ2 = 11.688, d.f. = 1, P = 0.001 | |
| Effect of cardinal directions | Wald - Ⲭ2 = 1.501, d.f. = 3, P = 0.682 | Wald - Ⲭ2 = 0.533, d.f. = 3, P = 0.912 | |
| Effect of interaction between factors | Wald - Ⲭ2 = 1.150, d.f. = 2, P = 0.563 | Wald - Ⲭ2 = 2.758, d.f. = 2, P = 0.252 | |
| Results of two-way factorial ANOVA | Effect of the nurse plants | F(1,57) = 7.089, P = 0.011 | F1,57 = 6.151, P = 0.016 |
| Effect of cardinal directions | F(3,57) = 0.235, P = 0.871 | F(3,57) = 0.237, P = 0.870 | |
| Effect of interaction between factors | F(3,57) = 0.494, P = 0.688 | F(3,57) = 1.193, P = 0.321 | |
| Results of logistic regressions with categorical variables | AUC value of the GLM | AUC = 0.923 | AUC = 0.902 |
| Effect of the nurse plants | Log - likelihood = 15.561, Ⲭ2= 6.968, d.f. = 1, P < 0.001 | Log - likelihood = 12.956, Ⲭ2= 13.968, d.f. = 1, P < 0.001 | |
| Effect of cardinal directions | Log - likelihood = 21.894, Ⲭ2= 2.540, d.f. = 3, P < 0.468 | Log - likelihood = -12.449, Ⲭ2= 1.013, d.f. = 3, P < 0.798 | |
| Effect of interaction between factors | Log - likelihood = 11.709, Ⲭ2= 4.784, d.f. = 2, P < 0.091 | Log - likelihood = 19.546, Ⲭ2= 0.788, d.f. = 2, P < 0.852 | |
| Results of linear regressions with categorical variables | Coefficient of determination | R2 = 0.202 | R2 = 0.163 |
| Goodness-of-fit ANOVA | F(3,57) = 1.119, P = 0.377 | F(7,31) = 0.917, P = 0.506 |
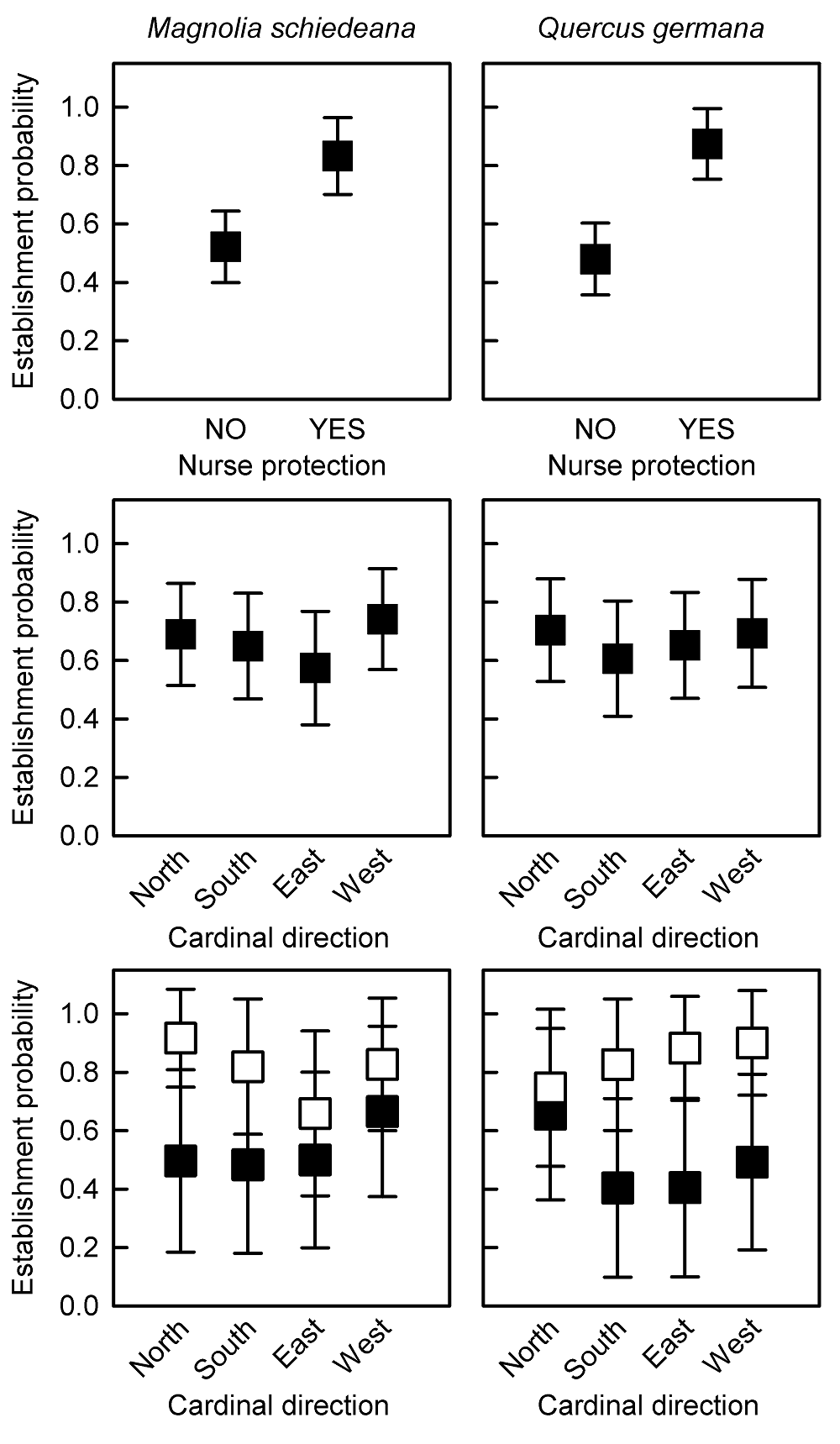
Figure 2 Establishment probabilities (marginal means ± 95 % C.I.) of Magnolia schiedeana (left panels) and Quercus germana (right panels) estimated with GLM. Upper panels show the establishment probabilities of saplings transplanted without nurse protection and beneath the canopies of Liquidambar styraciflua, irrespective of the cardinal direction. Middle panels show the establishment probabilities of saplings at each cardinal direction, irrespective of nurse protection. Lower panels show the establishment probabilities of saplings for between-level combinations of these two factors (black symbols = unprotected saplings; white symbols = nurse-protected saplings).
The results of the two-way factorial ANOVA indicated that growth rates of M. schiedeana saplings were lower beneath L. styraciflua canopies than outside them, while annual growth rates of Q. germana saplings were higher beneath the nurses (Table 1, Figure 3). Nevertheless, neither the cardinal direction nor its interaction with nurses affected growth rates of beneficiary species (Table 1, Figure 3).

Figure 3 Annual growth rates (averages ± 95 % C.I.) of Magnolia schiedeana (left panels) and Quercus germana (right panels). Upper panels show the growth rates of saplings transplanted without nurse protection and beneath the canopies of Liquidambar styraciflua, irrespective of the cardinal direction. Middle panels show the growth rates of saplings at each cardinal direction, irrespective of nurse protection. Lower panels show the growth rates of saplings for between-level combinations of these two factors (grey symbols = unprotected saplings; white symbols = nurse-protected saplings).
The logistic regressions indicated the establishment probabilities of saplings of M. schiedeana and Q. germana increase as canopies of L. styraciflua individuals become larger, but no differences in the magnitude of these effects were found among cardinal directions (Table 1, Figure 4). Conversely, annual growth rates of both beneficiary species did not relate with the canopy cover of nurses at any cardinal direction (Table 1, data not shown).

Figure 4 Effects of increasing canopy cover of Liquidambar styraciflua on the establishment probabilities of Magnolia schiedeana (left panel) and Quercus germana (right panel). These relationships were estimated with logistic regressions and a single response curve is shown for each species because no effects of cardinal directions were found.
Discussion
Our results indicate that transplanted L. styraciflua successfully establish in deforested areas. This confirms that this species is a pioneer tree that colonizes human- disturbed areas of Mexican TMCF (Pedraza & Williams-Linera 2003, Ramírez-Bamonde et al. 2005). Our results also suggest that, after L. styraciflua establish and develop their canopies, they act as effective nurses facilitating the establishment of mid- and late successional species - in this case, M. schiedeana and Q. germana, respectively. These findings support the use of positive plant-plant interactions for restoring Mexican TMCF and increasing establishment rates of trees belonging to advanced successional stages in early- successional environments (Ramírez-Marcial et al. 2006, Williams-Linera et al. 2016). Similar positive effects for L. styraciflua in TMCF were registered in the state of Veracruz (eastern Mexico), where human-planted individuals of this tree also enhanced the establishment of late-successional species beneath their canopies (Ramírez-Bamonde et al. 2005). However, despite the elevated diversity of pioneer plants that may act as nurses for late-successional species in disturbed areas of Mexican TMCF, the wide distribution range that L. styrciflua has in this country and its elevated capability for recruiting in open habitats (Williams-Linera et al. 2016) makes it a promising nurse tree for accelerating the restoration of these forests.
Although we did not measure microenvironmental conditions beneath and outside canopies of L. styraciflua, its positive effects on other tree species are likely due to the provisioning of shade and the amelioration of extreme high temperatures. This suggestion relies in the results of studies conducted in both TMCF and other ecosystems, which have shown that the elevated solar radiation and temperatures that predominate in deforested areas induce photoinhibition processes in saplings of late successional trees, constraining their establishment in these habitats (González-Salvatierra et al. 2013, Quevedo-Rojas et al. 2018). Therefore, the amelioration of these extreme environmental conditions could explain the higher establishment probability estimated for M. schiedeana and Q. germana saplings that grew beneath the canopies of L. styraciflua, as compared with the recruitment probability estimated for nurse-unprotected saplings. On the other hand, although the improvement of soil moisture beneath nurse canopies has been widely reported as another key mechanism that explains positive plant-plant interactions (Badano et al. 2009, Armas et al. 2011, Prieto et al. 2011), we propose that this process is unlikely to occur in TMCF because water availability usually does not constitute a limiting factor for plant recruitment in these forests (Jarvis & Mulligan 2011).
As we did not find effects of cardinal directions on the recruitment and performance of M. schiedeana and Q. germana saplings that grew beneath the canopies of L. styrciflua, it was not a critical determinant for the magnitude of the positive effects that these nurses have on beneficiary species. This contradicts previous studies reporting that beneficiary plants preferentially establish and grow on certain cardinal directions where microenvironmental conditions are less harsh (Franco & Nobel 1989, Valiente-Banuet & Ezcurra 1991, Drezner & Garrity 2003). Nevertheless, as far as we are aware, the effects that the interaction between cardinal directions and nurse plants may have on the establishment of tree species belonging to advanced successional stages have not been evaluated in TMCF, and our results suggests that, in these ecosystems, these effects would be negligible - i.e., the effects of nurses on beneficiary species are similar on all cardinal directions.
Despite the facilitative effects that L. styraciflua exerts on the establishment of tree species belonging to advanced successional stages, our restoration trial indicated that the association between nurses and beneficiary species may also induce negative interactions. These negative effects were found on M. schiedeana, whose saplings displayed lower net annual growth rates beneath the canopies of L. styraciflua than outside them. This suggests that interspecific competition between these nurses and young individuals of mid-successional trees may be operating in opposition to facilitation. In TMCF, water availability usually does not constrain plant growth (Jarvis & Mulligan 2011), but soils of these forests are nutrient poor and trees strongly compete for these resources (Long et al. 2015, Fahey et al. 2016). Therefore, while protection form unfavorable environmental conditions would explain the facilitative effects of L. styraciflua on the establishment of M. schiedeana saplings, belowground competition for soil nutrients could be responsible of the lower growth rates that saplings of the beneficiary species had when associated with the nurse. Here, it is important to note that facilitation and competition operate at the same time in plant communities and the final output of this interplay between positive and negative interactions depends on the balance between them - i.e., positive effects result when the intensity of facilitation is higher than that of competition, while negative effects result in converse situation (Bertness & Callaway 1994, Bruno et al. 2003). Thus, our findings suggest that, although L. styraciflua facilitates the establishment of M. schiedeana and this can enhance the success of restoration programs conducted in TMCF, the competitive effects that nurses exert on the growth of mid-successional trees may delay the recovery of ecosystem services.
Conversely to that reported for M. schiedeana, saplings of Q. germana displayed both higher recruitment probabilities and higher growth rates when protected beneath the canopies of L. styraciflua. This indicates that the establishment and development of saplings of late-successional trees in fallow lands of Mexican TMCF is enhanced by facilitation. Such an elevated reliance on nurses may be attributed the elevated shade-tolerance of these species, which mainly germinate and establish in forests understories rather than in open areas (Rüger et al. 2008, Muñiz-Castro et al. 2012). This does not mean, however, that negative interactions between nurse plants and saplings of late-successional trees are not occurring, but instead indicates that facilitation widely overcome the effects of any competitive effect. Therefore, our results suggest that reintroducing late-successional trees in fallows requires considering the use of nurse plants in the design of restoration strategies.
Another nurse effect that must be considered during the design of programs addressed to restore TMCF is the size of the canopies of nurse plants. On this issue, our results indicated that recruitment probabilities of both the mid- and late-successional trees increase with growing canopy cover of L. styraciflua. Further, as growth rates of these species did not relate with this nurse tree trait, it seems that growing canopy sizes of L. styraciflua does not promote competitive effects with the beneficiary species. This leads us to propose that forest restoration practitioners should introduce nurse plants with large, well-developed canopies in fallows because they are more likely to facilitate the establishment of mid- and late-successional tress than nurse plants with small canopies, while this would not affect the growth of these species.
It should be noted that although a greater establishment of individuals under the canopy sizes of L. styraciflua began, it should be considered that a treatment was not added in the design where the individual performance of the species without the presence of L. styraciflua will be evaluated, which prevents evaluating the effect size of interaction on individual performance.
From this study, we conclude that the joint transplanting of nurse plants with saplings of mid- and late-successional trees in their close neighborhood is an effective strategy for restoring Mexican TMCF in fallow lands. Thus, although we recognize that more detailed experiments are required to test whether the facilitation/competition mechanisms proposed above are actually operating between nurses and beneficiary species, we are confident that the restoration strategy proposed here will allow recovering the ecosystem services that these forests provide much faster than applying other restoration techniques.

