











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Las alteraciones de los reservorios de nutrientes en los ecosistemas son indicadores que permiten entender el cambio local, regional y global de los ciclos biogeoquímicos (CONABIO 2009, Chapin et al. 2011, Schlesinger & Bernhardt 2013, Huang et al. 2017). La modificación de la cobertura vegetal tiene implicaciones directas en las dinámicas de los reservorios de carbono, afectando por ende la concentración atmosférica de CO2 (Vitousek 1992, MEA 2005, IPCC 2014). Específicamente, el cambio de uso y cobertura del suelo (CUCS) se identifica como el principal precursor de la modificación de los almacenes de carbono, pues a menudo este proceso resulta en el cambio de composición y estructura de las comunidades vegetales en los ecosistemas terrestres (Ojima et al. 1994, Lambin et al. 2001, Pielke et al. 2013, Tadesse et al. 2014, Clerici et al. 2019).
Las zonas áridas y semiáridas son de gran importancia en la dinámica del carbono a nivel global, no únicamente por el hecho de que representan alrededor del 40 % de la cobertura de la superficie terrestre, sino también por el gran porcentaje de Productividad Primaria Neta (PPN) global al que dan soporte (> 30 %) (Grace et al. 2006, Poulter et al. 2014, Ahlström et al. 2015, Huang et al. 2017, Biederman et al. 2018, Wang et al. 2019). Así mismo, estos ecosistemas representan el principal factor de variación en la concentración interanual de carbono atmosférico, debido a la gran variabilidad de su respuesta a las condiciones climáticas (Poulter et al. 2014, Ahlström et al. 2015). Por lo cual, entender el impacto del cambio de uso de suelo en los almacenes de carbono de zonas áridas, resulta de gran importancia.
Los ecosistemas áridos y semiáridos enfrentan gran presión por las actividades antropogénicas de carácter primario (agricultura y ganadería). Específicamente, se estima que 26 % de la superficie terrestre está destinada a la ganadería y un 80 % de la superficie dedicada a ésta, ocurre en zonas áridas y semiáridas (Marcos et al. 2020). Aun cuando la presión por forrajeo constituye en sí un precursor de cambio importante de la cobertura de la vegetación, la remoción de especies vegetales nativas para la introducción de especies exóticas constituye otro factor que se debe considerar al estimar la modificación global de los ecosistemas áridos (Ojima et al. 1994, Lambin et al. 2001).
Con el fin de incrementar la capacidad forrajera de los ecosistemas nativos de zonas áridas, a menudo se recurre a la introducción de especies exóticas (Castellanos et al. 2002, Morales-Romero & Molina-Freaner 2016). En Norteamérica, una de las especies exóticas más utilizadas para incrementar la cantidad de forraje para el ganado, es el zacate Cenchrus ciliaris (buffel). Los efectos ecológicos provocados por la introducción de buffel han sido de interés para la comunidad científica desde el inicio de estas prácticas. Numerosos estudios relacionados a la implementación de praderas de zacate buffel, han reportado alteraciones en la biodiversidad, en los reservorios de nutrientes (incluido el carbono) y en el microclima, como consecuencia de la introducción de la especie (Castellanos et al. 2002, Arriaga et al. 2004 Franklin et al. 2006, Marshall et al. 2012, Castellanos et al. 2016, Bracamonte et al. 2017, De Albuquerque et al. 2019, Edwards et al. 2019, Elkind et al. 2019, Wright et al. 2021).
Más del 60 % del territorio mexicano está constituido por ecosistemas áridos y semiáridos, los cuales son usados primordialmente para el pastoreo de ganado bovino (Castellanos et al. 2010, Mora et al. 2013, Montaño et al. 2016). Se estima que, de esta superficie, > 20 millones ha han sido convertidas a pastizales de buffel, siendo el Noroeste de México una de las regiones más afectadas (Arriaga et al. 2004, Castellanos et al. 2010, Mora et al. 2013, Bracamonte et al. 2017, De Albuquerque et al. 2019). Tras la modificación a la cobertura de especies leñosas, para el establecimiento del zacate, se estima que existe una reducción del reservorio de carbono, pues se ha reportado que las praderas de buffel contienen entre 0.47 y 1.88 Mg C/ha, mientras que la vegetación nativa de zonas áridas presenta entre 2.35 a 9.4 Mg C/ha, así mismo se reporta que a mayor cobertura de zacate, menor es la capacidad de almacenamiento carbono, como consecuencia de la pérdida de especies leñosas (Búrquez et al. 1998, Bradley et al. 2006, Montaño et al. 2016).
En México, los estudios relacionados al almacén de carbono en comunidades vegetales se han llevado a cabo mayormente en zonas húmedas (Montaño et al. 2016). En contraste, para zonas áridas y semiáridas, la poca información disponible se encuentra de forma dispersa y en periodos cortos de tiempo (Montaño et al. 2016, Briones et al. 2018). Esta limitación de información y la alta heterogeneidad de las zonas áridas incrementa la incertidumbre del conocimiento actual sobre los efectos en la dinámica del carbono por CUCS (Collins et al. 2014, Biederman et al. 2018, Pordel et al. 2018, Hinojo-Hinojo et al. 2019). Dada la extensión que las zonas áridas presentan en el país, es de gran importancia la caracterización de las dinámicas de los almacenes de carbono presentes en ellas.
Los métodos convencionales para la estimación de biomasa/almacén de carbono son tardados, requiriendo la cosecha y pesado de la biomasa de especies o comunidades vegetales para posteriormente estimar el contenido de los nutrientes en la misma. Lo anterior hace el muestreo a gran escala espacial y temporal complicado y costoso (Picard et al. 2012, Briones et al. 2018, Pordel et al. 2018, de Jong et al. 2019, Gallardo-Salazar & Pompa-García 2020). Por otro lado, se tienen métodos secundarios como lo es el desarrollo de ecuaciones alométricas, que consisten en relacionar biomasa con mediciones directas de características morfológicas de la vegetación (modelo de crecimiento) (Picard et al. 2012). Finalmente, para obtener estimaciones análogas a los métodos anteriormente mencionados, pero a mayores extensiones espaciotemporal, se han incorporado herramientas de percepción remota, tales como LIDAR (Light Detection and Ranging, por sus siglas en inglés) o imágenes multiespectrales que permiten comprender la estructura de los ecosistemas de manera indirecta (Mohd Zaki et al. 2017).
Como consecuencia del desarrollo tecnológico, y obedeciendo a la necesidad de obtener imágenes de alta resolución para el análisis de la vegetación (vía percepción remota), se ha incorporado el uso de cámaras RGB montadas en vehículos aéreos no tripulados (VANT). Estas imágenes nos han permitido obtener información detallada de cobertura de suelo y estructura de la vegetación (altura, área de copa, identificación de individuos y especies), permitiendo nuevas alternativas para el análisis preciso y eficiente (tiempo y recursos económicos) de reservorios de nutrientes (Kachamba et al. 2016, Vázquez-Arellano et al. 2018, Librán-Embid et al. 2020, Navarro et al. 2020). Particularmente, en México se han desarrollado estudios con VANT para el análisis de composición, estructura, distribución de vegetación y en menor medida almacén de carbono en agroecosistemas constituidos por monocultivos (Cornejo-Denman et al. 2018, Botello-Aguillón et al. 2019, Gallardo-Salazar & Pompa-García. 2020, García-Martínez et al. 2020, Gómez-Sapiens et al. 2021). Sin embargo, no se han realizado estudios, tratando de evaluar reservorios de carbono en ecosistemas áridos y semiáridos.
Buscando mejorar el entendimiento de las dinámicas del carbono en regiones áridas, así como el efecto de CUCS en las mismas, el presente trabajo tiene como objetivo analizar el almacén de carbono en biomasa aérea (ACBA) de individuos leñosos (árboles y arbustos), en comunidades vegetales de matorral subtropical y pradera de buffel en el Desierto Sonorense (región central de Sonora). Como hipótesis se establece que, a través de la medición de parámetros de la vegetación obtenidos vía percepción remota, provenientes de imágenes RGB alta-resolución, se logrará obtener estimaciones precisas de ACBA para las comunidades analizadas. Para dar respuesta a la hipótesis planteada en el presente trabajo, se llevó a cabo trabajo de campo, en combinación con técnicas de fotogrametría utilizando imágenes RGB alta-resolución (obtenidas mediante cámaras montadas en VANT) de comunidades nativas y modificadas con la introducción de zacate buffel.
Materiales y métodos
Para entender la dinámica de carbono aéreo en el matorral semiárido y la pradera de buffel, bajo uso de suelo ganadero, se realizaron mediciones de vegetación y, posterior a ello, estimaciones de ACBA usando ecuaciones alométricas (método directo). A su vez, se obtuvieron imágenes aéreas RGB alta-resolución, con el fin de obtener mediciones indirectas (por medio de percepción remota-fotogrametría) de la vegetación y poder utilizar ecuaciones alométricas para así estimar ACBA (método indirecto).
Sitio de estudio. El área de estudio se localiza en la franja central de Sonora entre los paralelos 28º 40´ y 28º 45´ N y los meridianos 110º 30´ y 110º 35´ O (Figura 1), dentro del municipio de La Colorada, el cual colinda al Oeste con el municipio de Hermosillo, capital del estado. Las condiciones del sitio son semiáridas, con una precipitación promedio anual de 476 mm, bajo un régimen bimodal. Presenta eventos de precipitación en temporada de verano e invierno, dando lugar a los eventos más prolongados e intensos en verano por la influencia del monzón de Norteamérica (Méndez-Barroso et al. 2009). La temperatura se encuentra en un rango de 5 a 40 °C, con una media anual de 22.8 °C (Hinojo-Hinojo et al. 2016).

El uso de suelo es principalmente ganadero; el manejo del ganado se rige por una rotación continua a diferentes potreros, donde se pueden encontrar extensiones con cobertura de matorral subtropical (MS) y de manera contigua pradera de buffel (BF). Particularmente, el establecimiento de la pradera consistió en el desmonte parcial de la vegetación leñosa (dejando solo algunos individuos arbóreos), y posterior al proceso de desmonte, se rastreó el suelo para sembrar el zacate (Hinojo-Hinojo et al. 2016).
La vegetación leñosa y dominante, en el matorral, la constituye Olneya tesota, Fouqueria macdougalii e Ipomoea arborescens (Hinojo-Hinojo et al. 2019), y en el caso de la pradera, presenta dominancia por Olneya tesota, Prosopis velutina, Mimosa laxiflora y Jatropha cardiophylla (Hinojo-Hinojo et al. 2019).
Inventario de comunidades vegetales. El levantamiento de métricas de la vegetación se llevó a cabo durante la temporada húmeda (agosto, septiembre y octubre), en cinco áreas constituidas por vegetación nativa, y cinco áreas en donde la cobertura vegetal es dominada por buffel en una proporción > 20 %. El arreglo experimental consistió en parcelas cuadradas anidadas de 30 × 30 m para la medición de árboles, y dentro de esta, una subparcela para los arbustos (15 × 15 m) (Bonham 2013). Posteriormente, los individuos leñosos se contaron en cada parcela, se identificaron a nivel de especie y se les midieron las variables: diámetro a la altura de pecho, altura máxima de copa y dos radios de copa (Tabla 1).
Tabla 1 Variables medidas por individuo y forma de vida, acorde a PMC 2015.
| Forma de vida | Característica morfológica | Abreviatura |
|---|---|---|
| Árboles | Diámetro a la altura de pecho | DAP |
| Árboles/arbustos | Altura máxima de copa | AMC |
| Arboles/arbustos | Radios de copa | r1 y r2 |
Estimación del almacén de carbono en biomasa aérea del estrato leñoso (método directo). Para la estimación de biomasa aérea de los individuos leñosos fueron utilizadas las ecuaciones alométricas que se presentan en la Tabla 2. La ecuación alométrica propuesta por Búrquez et al. (2010) fue desarrollada como una ecuación no lineal, para matorral, en condiciones topográficas de planicie, y que utiliza como variable independiente el volumen de vegetación, el cual se estima con la Ecuación 1 por individuo, posteriormente, los cálculos por individuo son sumados y el total se divide por el área de parcela para normalizar el valor de volumen. Complementando la ecuación anterior, fueron utilizadas las ecuaciones alométricas disponibles por especie, Ipomoea arborescens y Prosopis velutina (McClaran et al. 2013, Rojas-García et al. 2015).
Tabla 2 Ecuaciones alométricas para la estimación de biomasa aérea de vegetación de zonas áridas y semiáridas.
| Tipo de ecuación | Ecuación | Variable independiente (unidad) | Fuente |
|---|---|---|---|
| Comunidad (Matorral) | y = 896.50x1.16 | Volumen de vegetación (m3 m2) | Búrquez et al. 2010 |
| Especie (Ipomoea arborecens) | ln(y) = -4.00 + 2.65 ln(x) | Diámetro a altura de pecho (cm) | Rojas-García et al. 2015 |
| Especie (Prosopis Velutina) | Ln(y) = (-0.67+1.55(ln(x))) 1.06 | Área de copa (m2) | McClaran et al. 2013 |
Dada la alta densidad de individuos de Olneya tesota en las comunidades y a la similitud que presentan en su fisonomía con Prosopis velutina, se consideró pertinente el uso de la ecuación propuesta por McClaran et al. (2013) para la estimación de la biomasa aérea en individuos de esta especie. Para ello, se ajustó la ecuación incorporando un factor de corrección (1.25), que representa la proporción de la densidad de la madera de Olneya (densidad = 0.99 g/cm3) en relación con Prosopis (densidad = 0.79 g/cm3), la cual es reportada para ambas especies por Silva (2008).
Tras estimar la biomasa aérea, por estrato (arbustivo y arbóreo), se calculó el contenido de carbono, aplicando el factor de conversión establecido para zonas áridas de México: 47 % de la biomasa es carbono (Briones et al. 2018). De manera adicional, fue estimado ACBA utilizando únicamente la ecuación propuesta por Búrquez et al. (2010), para posteriormente comparar con las estimaciones del método indirecto, ya que es posible replicar el método para la estimación de ACBA con esta ecuación a partir de las métricas de vegetación, calculadas con imágenes aéreas capturadas mediante VANT.
Donde V es volumen de vegetación por área en m3 m2, es decir, una normalización de la variable acorde al tamaño de la parcela de muestreo (900 m2 árboles y 225 m2 arbustos). AC es el área de copa en m2 y AMC es altura máxima de copa en m.
Donde r1 y r2 son radios de copa (en m) medidos de manera ortogonal uno con respecto del otro.
Índices de diversidad. Para evaluar la diversidad en las comunidades, se utilizó el índice de riqueza específica independiente al tamaño de la muestra, índice de Margalef, el cual se basa en la relación entre número de especies y número de individuos, permitiendo la comparación entre sitios (Campo & Duval 2014). Así mismo, se determinó el índice de diversidad de Shannon-Wiener, el cual se rige por estructura y abundancia proporcional para comprender la variación de especies (Moreno 2001, Del Río et al. 2003). También, a través del índice de Pielou se analizó la equitatividad de los individuos por especie (Moreno 2001). Los índices se calcularon con las siguientes ecuaciones:
Índice de Margalef
Donde S es el número total de especies identificadas y n es el número total de individuos.
Índice de Shannon-Wiener
Donde pi es la abundancia proporcional de la especie i determinada como el número de individuos de la especie i dividido entre el número total de individuos de la muestra.
Índice de Pielou
Estimación del almacén de carbono en biomasa aérea del estrato leñoso (método indirecto). Para obtener las métricas indirectas de la vegetación, fue necesario la construcción de ortomosaicos de los sitios de muestreo directo. Se realizó el levantamiento de imágenes de alta resolución a través de una cámara RGB (cámara FC350_3.6_4000 × 2250, 12 MP y ángulo de apertura 96°) montada en un VANT (Phantom 3 Advanced, SZ DJI Technology Co, Shenzhen, Guangdong, China). Las parcelas inventariadas en terreno fueron sobrevoladas, extendiéndose a un área aproximada de una hectárea.
Los vuelos fueron automatizados, programados en la aplicación Altizure Beta (Intern at Shenzhen Zhuke Innovation Technology), con las siguientes características: altura de 35 m y traslape lateral y frontal de 90 %. La serie de imágenes por vuelo estuvo compuesta en promedio por 350 elementos. Posteriormente, las series fueron procesadas en el programa de fotogrametría PIX4D, el cual utiliza el proceso de structure from motion (SfM) para la generación de los ortomosaicos (Iglhaut et al. 2019). A partir de los ortomosaicos por sitio, se crearon capas digitales (mapas) que contenían información referente al área de copa (AC), para mapear y delimitar a los individuos leñosos en MS y BF (Figura 2). Esto se realizó mediante la interpretación y digitalización directa del AC de la vegetación leñosa presente en los ortomosaicos, utilizando los programas QGIS y ArcMap (Environmental Systems Research Institute, California, Estados Unidos). El proceso de digitalización se efectuó de manera separada para los arbustos y los árboles, para obtener capas digitales de AC por estrato.

Figura 2 A) Ortomosaico de comunidad de matorral subtropical, y B) ejemplo de la capa final de digitalización de área de copa de individuos leñosos.
Posterior a la digitalización, se analizó la relación entre las métricas censadas directamente, AC y altura máxima de copa (AMC) por medio de un análisis de regresión (Montgomery et al. 2021), con el fin de utilizar AC indirecta para el cálculo de AMC. Para el análisis de regresión se utilizaron 432 individuos leñosos, de las especies censadas en las comunidades de MS y BF.
A partir de los parámetros obtenidos de manera indirecta, es decir, AC proveniente de la digitalización, y AMC estimada con el modelo obtenido de la regresión, se calculó el volumen de la vegetación para estimar biomasa aérea, y enseguida ACBA. Finalmente, para analizar la diferencia estadística de los resultados con los métodos directos e indirectos, se utilizó la prueba de t de Student.
Resultados
Índices de diversidad. A través del censo de la vegetación leñosa se obtuvo que matorral presenta 2378 Ind./ha, mientras que la pradera presenta 915 Ind./ha. Las especies dominantes en el estrato arbóreo en MS son Olneya tesota e Ipomoea arborescens, por su parte, las praderas presentan dominancia por Prosopis velutina y Olneya tesota. Este estrato representa < 6 % de los individuos totales, en ambas comunidades (Tabla 3). Dentro del estrato arbustivo, la especie dominante es Jatropha cardiophylla en las dos comunidades.
Tabla 3 Número total de especies identificadas, número de individuos por hectárea, rango de las métricas de altura máxima de copa y área de copa, por estrato y por comunidad, matorral subtropical (MS) y Pradera de buffel (BF).
| Especies identificadas | Ind/ha | Altura máxima de copa (mínimo-máximo) m |
Área de copa (mínimo-máximo) m2 |
|
|---|---|---|---|---|
| Arbustos MS | 24 | 2,240 | 0.3-3.34 | 0.02-16.83 |
| Arbustos BF | 17 | 871 | 0.06-2.98 | 0.05-11.15 |
| Árboles MS | 9 | 138 | 2.16-14.57 | 2.54-51.67 |
| Árboles BF | 4 | 44 | 2.88-7.50 | 4.90-67.73 |
En relación con la morfología de las comunidades, se presentó una AMC promedio + DE en arbustivo de MS de 1.31 m ± 0.61 m, y en arbustos de BF, 1.27 m ± 0.53 m. Por su parte, el estrato arbóreo muestra un promedio de 4.48 m ± 2.19 m y 4.59 m ± 1.30 m, en MS y BF, respectivamente. En lo que concierne a el AC del estrato arbustivo, los promedios son 1.76 m2 ± 2.04 m2 en MS y 1.40 m2 ±1.47 m2 en BF. El AC del estrato arbóreo de MS es de 14.65 m2 ±12.26 m2, mientras que en BF es de 26.35 m2 ± 17.48 m2 (Tabla 3).
En términos de diversidad de individuos leñosos (Tabla 4), acorde al índice de riqueza de especies de Margalef, MS presenta riqueza moderada (> 2), mientras que BF presenta riqueza baja (< 2), la diferencia entre comunidades es significativa (W = 0, p < 0.05). A su vez, el índice de diversidad de Shannon-Wiener muestra que las comunidades analizadas tienen valores menores a 2, encontrando el valor más bajo para BF. En cuanto al índice de Pielou, se establece que los individuos en ambas comunidades se presentan de manera equitativa por especie, ya que se tienen valores mayores a 0.79. De acuerdo con la evidencia estadística con la que se cuenta para estos últimos índices (H´: t = -1.7734, df = 6.5, p = 0.1222, y para Ev´: t = 0.84532, df = 7.8859, p = 0.4228), no se presenta diferencias significativas entre las comunidades.
Tabla 4 Índices de diversidad, Margalef (DMg), Shannon-Wiener (H´) y Pielou (Ev´), por parcela censada, en las comunidades de matorral subtropical (MS) y Pradera de buffel (BF).
| MS | BF | |||||
|---|---|---|---|---|---|---|
| Parcela | DMg | H´ | Ev´ | DMg | H´ | Ev´ |
| P 1 | 2.32 | 2.02 | 0.88 | 1.68 | 1.43 | 0.73 |
| P 2 | 3.49 | 2.52 | 0.89 | 2.12 | 1.62 | 0.77 |
| P 3 | 2.19 | 1.73 | 0.79 | 1.61 | 1.47 | 0.91 |
| P 4 | 2.36 | 1.90 | 0.79 | 2.16 | 1.84 | 0.94 |
| P 5 | 2.42 | 1.64 | 0.68 | 2.10 | 1.88 | 0.90 |
| Promedio ± DE | 2.56 ± 0.52 | 1.96 ± 0.34 | 0.80 ± 0.08 | 1.93 ± 0.26 | 1.65 ± 0.20 | 0.85 ± 0.09 |
Almacén de carbono en biomasa aérea con método directo. El almacén de carbono en biomasa aérea total compuesto por los individuos leñosos en matorral fue de 10.35 Mg C/ha, mientras que en la pradera se presenta un total de 3.20 Mg C/ha. Al separar el reservorio de carbono aéreo por estrato, se observa que, en ambas comunidades los árboles almacenan la mayoría de éste (> 80 %; Tabla 5). Por otra parte, se encontró mayor capacidad de almacenamiento de carbono en el estrato arbustivo de MS, en comparación con BF, como se puede apreciar en la Tabla 5.
Tabla 5 Contribución por estrato al almacén de carbono de individuos leñosos, en matorral subtropical (MS) y praderas constituidas de buffel (BF). Ambas comunidades bajo uso de suelo ganadero.
| Comunidad | Estrato | Promedio ± DE almacén de carbono (Mg C/ha) |
Almacén de carbono total (Mg C/ha), estrato leñoso |
Almacén de carbono (%) |
|---|---|---|---|---|
| MS | Arbustivo | 1.65 ± 1.04 | 15.94 | |
| MS | Arbóreo | 8.70 ± 4.90 | 84.05 | |
| MS | Leñoso | 10.35 | ||
| BF | Arbustivo | 0.42 ± 0.32 | 13.12 | |
| BF | Arbóreo | 2.78 ± 1.82 | 86.87 | |
| BF | Leñoso | 3.20 |
El ACBA de MS y BF, acorde a las pruebas estadísticas, presenta diferencia significativa en el estrato arbustivo (t = -2.51, df = 4.79, P = 0.05), arbóreo (W = 2, P = 0.03) y de la misma manera, en el conjunto de todos los individuos leñosos (t = -2.13, df = 11.17, P = 0.05).
Almacén de carbono en biomasa aérea con método indirecto. A través de un análisis de regresión lineal, se encontró una correlación significativa (P < 0.01) entre AC y AMC, lo cual nos indica que AC puede ser usada para estimar AMC. En el mismo sentido, un coeficiente de determinación alto (R 2 = 0.74), indica que la variable independiente (AC) explica gran parte (74 %) de la variación en la respuesta (variable dependiente, AMC). Comprobando entonces la relación entre variables y una alta explicación de la varianza de AMC, se estableció como modelo de predicción la ecuación proveniente del análisis de regresión (AMC = 0.2809 (AC) + 0.9396).
En total, el almacén de carbono calculado para la comunidad leñosa de MS presenta 5.05 Mg C/ha, mientras que BF 2.46 Mg C/ha, en cuanto al método indirecto, MS presenta 8.01 Mg C/ha y BF 6.47 Mg C/ha. Si bien, la estimación con el método indirecto presenta una sobreestimación en contraste con el método directo, se logra explicar la varianza de este. Así mismo, se encontró correlación significativa entre ambos métodos (R 2 ( 0.7 y p < 0.05; Tabla 6 y Figura 3).
Tabla 6 Análisis de regresión entre almacén de carbono estimado con métodos directos e indirectos (variable dependiente almacén de carbono con parámetros in situ, y variable independiente almacén de carbono con parámetros indirectos), para estratos arbustivo, arbóreo y agrupados, en matorral subtropical y en praderas de buffel. Se identifica con * P < 0.01 y con ** P < 0.001.
| Estrato | Ecuación | Error DE. | Valor p | R2 | |
|---|---|---|---|---|---|
| MS | Arbustivo | y = 0.74x + 0.74 | 0.17 | < 0.05 | 0.85 |
| MS | Arbóreo | y = 0.47x + 0.20 | 0.04 | < 0.05** | 0.97 |
| MS | Leñoso | y = 0.40x + 0.95 | 0.04 | < 0.05** | 0.89 |
| BF | Arbustivo | y = 2.06x + 0.04 | 0.63 | < 0.05 | 0.77 |
| BF | Arbóreo | y = 0.33x - 0.06 | 0.11 | > 0.05 | 0.72 |
| BF | Leñosos | y = 0.29x - 0.31 | 0.04 | < 0.05** | 0.83 |
| MS y BF | Arbustivo | y = 0.87x + 0.42 | 0.12 | < 0.05** | 0.85 |
| MS y BF | Arbóreo | y = 0.43x - 0.07 | 0.09 | < 0.05** | 0.73 |
| MS y BF | Leñoso | y = 0.36x + 0.59 | 0.04 | < 0.05** | 0.75 |
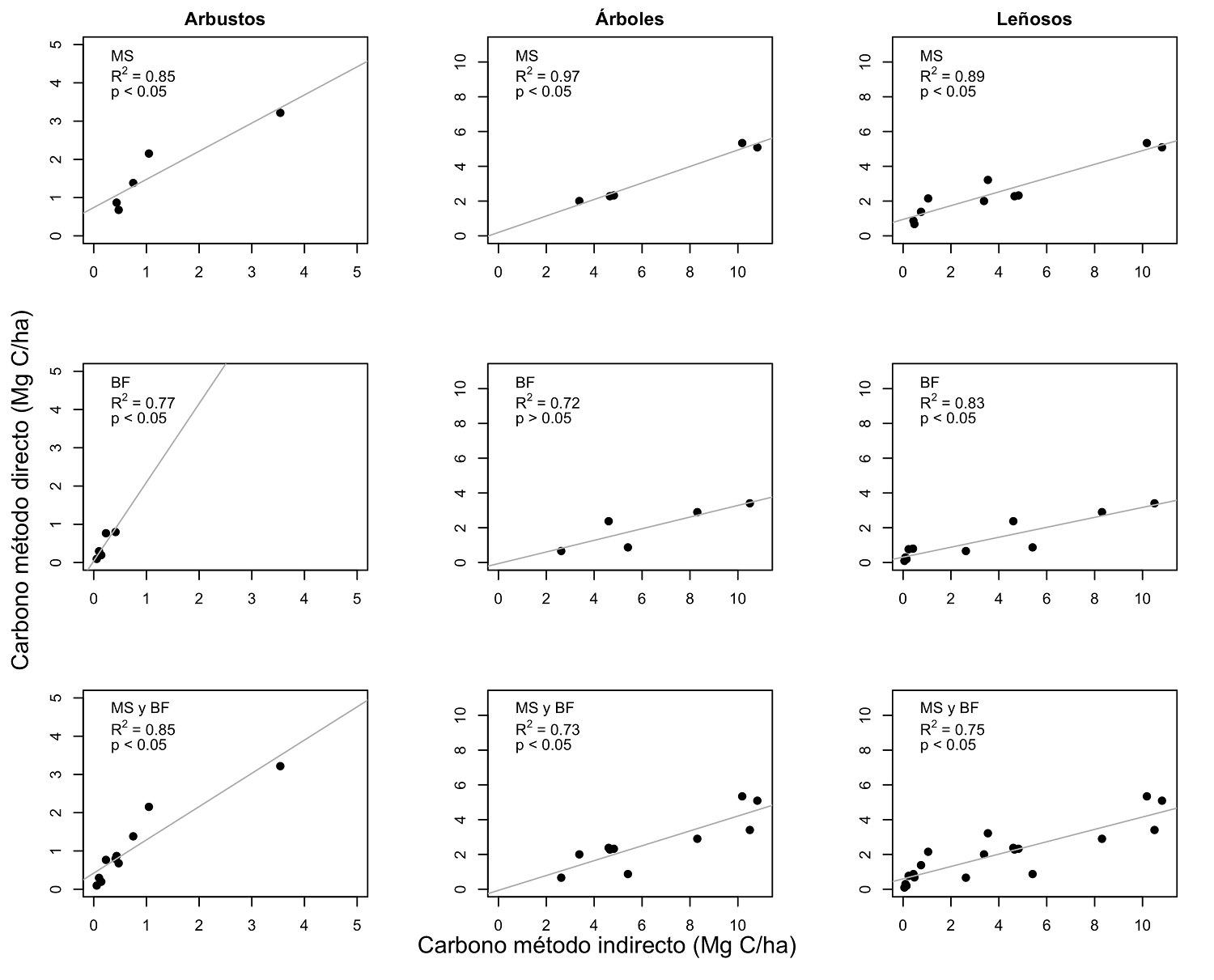
Figura 3 Diagramas de dispersión de almacén de carbono aéreo del estrato leñoso (arbustos y árboles), estimado con método directo (eje y) e indirecto (eje x), para comunidades de matorral subtropical (MS) y praderas de buffel (BF).
Si se desagregan los estratos, en MS el arbóreo presenta mayor correlación entre los métodos, observando una R 2 = 0.97, en cuanto que, el estrato arbustivo presenta una R 2 = 0.85. En cuanto a BF, se obtuvo una R 2 = 0.72 en el estrato arbóreo, mientras que en el arbustivo una R 2 = 0.77. Sin embargo, cabe destacar que aun cuando los valores obtenidos, sugieren una buena relación entre ambos métodos, las pruebas estadísticas no demuestran significancia para el estrato arbóreo en BF (p = 0.06; Tabla 6). Por otra parte, al integrar la información de ACBA de individuos arbóreos y arbustivos de las comunidades, se observan correlaciones significativas entre métodos (Figura 3), así mismo, coeficientes de determinación altos (R 2 > 0.70 y p < 0.05).
Discusión
De acuerdo con el presente análisis, en las comunidades de matorral subtropical donde se ha establecido zacate buffel, se observa una disminución tanto en el número de especies leñosas, como en el número de individuos por especie. Lo anterior es consistente con lo reportado en estudios previos, en donde se menciona que la implementación del zacate buffel con fines forrajeros a menudo desemboca en la reducción en la diversidad de la vegetación (Morales-Romero & Molina-Freaner 2008, Marshall et al. 2012, Bracamonte et al. 2017). Específicamente, se ha encontrado una reducción de individuos mayor al 50 % (tomando en cuenta el estrato arbustivo y arbóreo), y una reducción de la riqueza de especies del 30 y 50 % en arbustos y árboles, respectivamente. De acuerdo con el contraste de los índices de diversidad calculados para las comunidades, se sugiere diferencia estadísticamente significativa en la riqueza especifica de BF y MS. Sin embargo, en términos de diversidad y equitatividad, no se cuenta con suficiente evidencia estadística para establecer diferencias ambas comunidades. Lo anterior sugiere el cambio de la riqueza de especies en las comunidades, pero no en la presencia equitativa de individuos entre las especies, condición que fue reportada también por Bracamonte et al. (2017) para una comunidad del desierto Sonorense. Asimismo, Castellanos et al. (2002)yMorales-Romero & Molina-Freaner (2016) reportan alteraciones en el microclima y la composición de las comunidades vegetales de los matorrales convertidos a praderas de buffel, lo cual genera condiciones adversas para las especies nativas, y por ende la posible reducción de las mismas.
Es importante destacar que, de acuerdo a los resultados del presente estudio, la conversión de matorral subtropical a pradera de buffel, implica un decremento considerable de la cantidad de carbono almacenado en biomasa aérea (> 7 Mg C ha), siendo el reservorio de carbono del estrato arbustivo el más afectado por estas prácticas (desmonte y pastoreo). Esto es importante, ya que se ha reportado modificaciones a grandes extensiones de matorrales nativos (> 1,500,000 ha) dentro del Desierto Sonorense (Bracamonte et al. 2017).
Acorde a los resultados obtenidos en relación con los atributos morfológicos de la vegetación, la conversión a praderas de buffel, sugiere un incremento en el área de copa de los árboles presentes (Figura 4), pues del total de individuos que se presentan en estas áreas, alrededor del 45 % tiene un AC mayor a 25 m2 (solo el 15 % de los individuos arbóreos en MS presentan tallas similares). Así mismo, la variabilidad en las estimaciones del almacén de BF indica que la respuesta de MS ante la introducción de buffel no se presenta de la misma forma para todos los sitios, lo cual tiene una relación directa con el legado ecológico y la forma de uso previo de los sitios muestreados (Castellanos et al. 2010, Mora et al. 2013). Tomando en cuenta ambas aseveraciones, podemos indicar que la distribución del carbono en biomasa aérea (en especies leñosas) para las praderas de buffel, no puede ser generalizada para todos los sitios en donde se ha introducido el zacate.
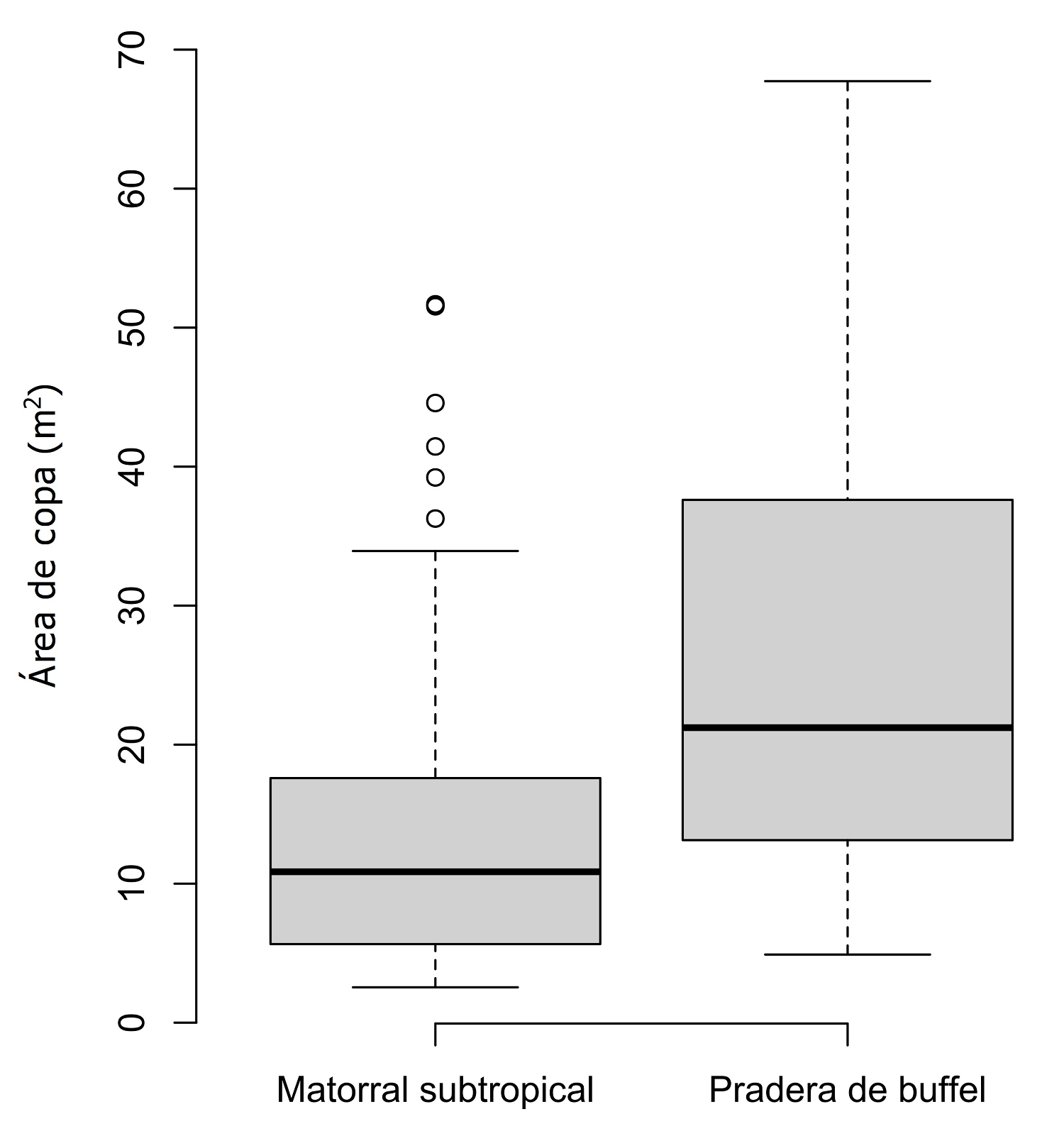
Figura 4 Diagrama de caja y bigote generado con datos de área de copa (m2) de individuos arbóreos, censados en matorral subtropical y praderas de buffel del Noroeste de México.
Se encontró suficiente evidencia estadística para comprobar relación entre los métodos directo e indirecto, sugeridos en el presente trabajo, lo cual indica que es posible estimar ACBA, a través de imágenes RGB alta-resolución. Específicamente, mediante la utilización de técnicas fotogramétricas en imágenes aéreas obtenida mediante VANT, se obtuvieron modelos adecuados para la predicción del almacén de carbono aéreo contenido en especies leñosas presentes en MS y BF. Al combinar todos los datos disponibles de ACBA (árboles y arbustos de MS y BF), se obtuvo un modelo general con buena capacidad de predicción, indicando que la cantidad de carbono aéreo en estas comunidades presenta una relación directa con el área de copa (variable utilizada en el método indirecto, al correlacionar el área de copa con la altura máxima de copa de los individuos), y por ende que es posible utilizar el mismo modelo para las dos comunidades estudiadas.
Observando la alta correlación entre el método directo e indirecto, consideramos que es posible predecir biomasa (ACBA) en los estratos leñosos de las comunidades analizadas, utilizando aproximaciones vía VANT. Lo anterior sugiere la posibilidad de hacer más eficiente el proceso de muestreo en campo. Resultados similares se pueden encontrar en otros trabajos que han estudiado agroecosistemas específicos, bosques y manglares (Botello-Aguillón et al. 2019, Effiom et al. 2019, González-Jaramillo et al. 2019, García-Martínez et al. 2020, Navarro et al. 2020).
Se concluye que el cambio de cobertura vegetal en matorral subtropical provocado para la introducción y establecimiento de buffel, implica la simplificación a la estructura de la comunidad (menor número de especies), así como la reducción del ACBA de los estratos leñosos (principalmente en el estrato arbustivo). También, se observa gran variabilidad del carbono contenido en este componente en las praderas de buffel, dificultado la predicción (tendencia) de este cambio. Por otra parte, mediante el uso de imágenes RGB alta-resolución, fue posible detectar la modificación por CUCS en los reservorios de carbono en las comunidades analizadas. Finalmente, es fundamental que en futuros trabajos de esta naturaleza se busque incluir el estrato compuesto por especies herbáceas, para comprender su contribución, así como la dinámica del carbono aéreo contenido en la biomasa vegetal en su totalidad.

