











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Mountains are landforms that stand out conspicuously from the surrounding plateaus and generally have complex relief and steep slopes (Hoorn et al. 2018). Mountains located in regions of high biodiversity, such as Mexico, are species-rich sites due to abrupt changes in altitude and climate over short distances (Körner et al. 2011). These changes lead to the characteristic topographic heterogeneity of mountainous regions (commonly known as “sierras” in Mexico), generating a wide variety of habitats and a rich diversity of species. In Mexico, mountains can be considered islands of temperate biota surrounded by more thermophilic biota found in the surrounding dry or humid tropical forests. This archipelagic behavior has resulted in the evolution of many endemic taxa over the course of the geological development of the mountains.
Mountain slopes form hydrographic basins and modify the regional climate, which allows different biomes to be found along this intricate pattern (Hoorn et al. 2018). They influence the organization of vegetation zones along altitudinal gradients, each gradient characterized by unique sets of species. It is therefore difficult to establish a precise boundary between the mountain and the lowland hills that receive the runoff from the higher elevations. This result, for example, in different concepts of mountainous regions in Mexico, as exemplified in Figure 1, where two different conceptualizations of the Sierra Madre Oriental (SMOR) result in different placement of the region’s boundary.

Figure 1 A. Two different circumscriptions of the Sierra Madre Oriental (SMOR), the most important mountain range in northeastern Mexico. The more restrictive limit proposed by CONABIO (1997) is shown in dark gray and the broader area defined by Cervantes-Zamora et al. (1990) is in light gray. B. Mountains of northeastern Mexico (colored areas) as defined by Körner et al. (2011) overlaid with the SMOR polygon proposed by Cervantes-Zamora et al. (1990) (black outline).
It is evident that the SMOR as defined by Cervantes-Zamora et al. (1990) or CONABIO (1997) is not entirely composed of mountains (Figure 2). There are flat portions of the territory that are included in the sierra whose floristic composition is likely different from that observed in the more rugged areas. Therefore, it is important to differentiate characteristic features of the mountainous parts of the SMOR from those corresponding to the plateaus and flatter surrounding areas.
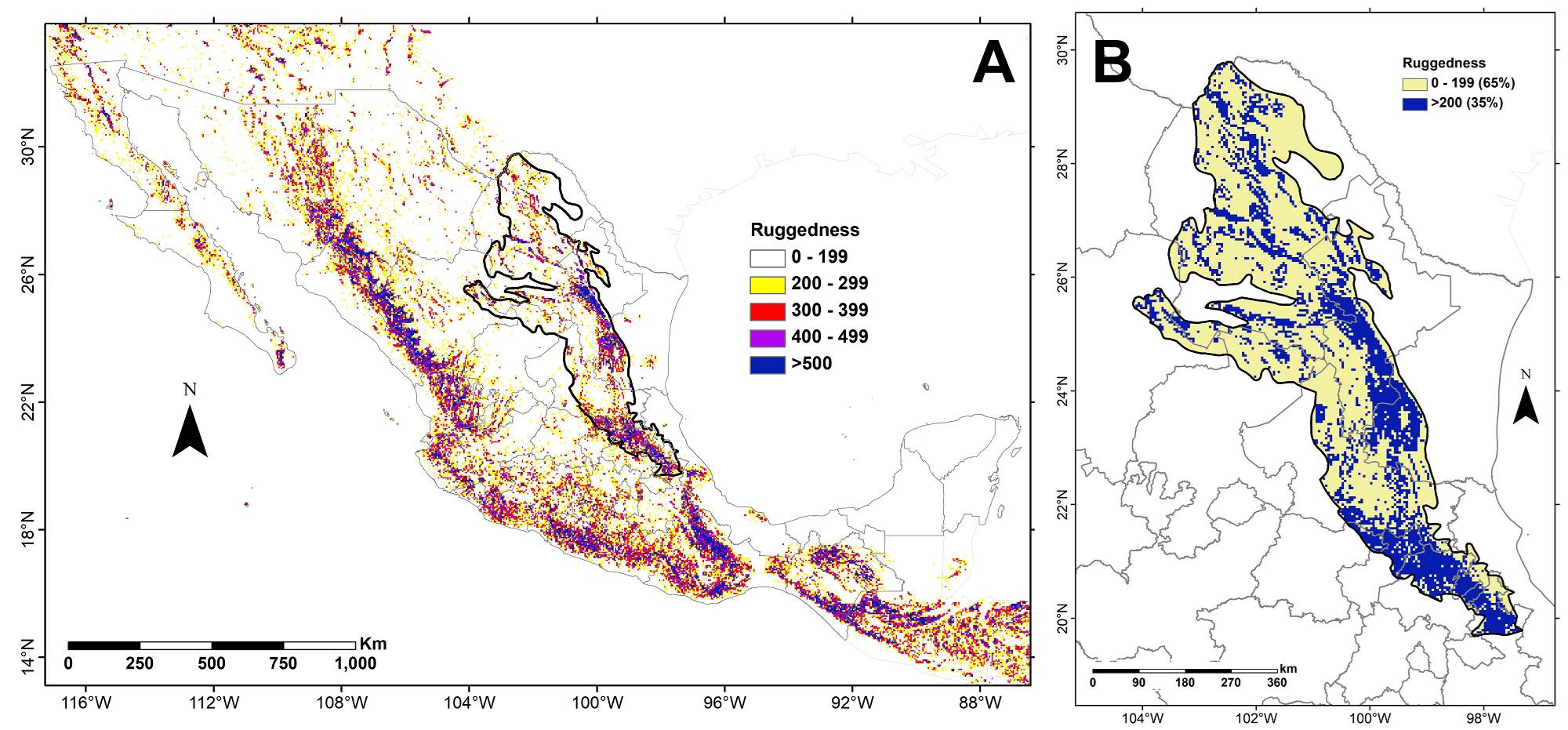
Figure 2 A. Ruggedness map of Mexico indicating the mountainous regions. B. Detail of the SMOR, adapted from the ruggedness map provided by Körner et al. (2011) who consider mountains when a ruggedness index greater than 200 is observed. The black outline in both panels indicates the SMOR polygon as proposed by Cervantes-Zamora et al. (1990).
The Sierra Madre Oriental, located in northeastern Mexico (Figure 1), is a complex land mass bordered by another mountain range (Trans-Mexican Volcanic Belt) to the south, the highland Mexican Plateau to the west, and the coastal plain of the Gulf of Mexico to the east. Within the conceptual limits of the SMOR, neighboring regions contain a variety of biomes, climates, and other factors that make it difficult to precisely identify their boundaries and, consequently, their species diversity.
For example, Figure 3 shows the distribution of the five main biomes proposed by Villaseñor & Ortiz (2014) throughout the SMOR and neighboring areas. The map shows that the SMOR is not only made up of montane biomes, but also includes large portions of other biomes, such as xerophilous scrub or tropical forests. This complex mosaic of intermingled biomes raises the question of what exactly constitutes the SMOR, considered to be a mountain range characterized by assemblages of mountains as defined above.

Figure 3 Main biomes according to Villaseñor & Ortiz (2014) within the SMOR polygon as defined by Cervantes-Zamora et al. (1990). The sierra was divided into grid cells of 1° latitude and longitude (N = 42) with a total area included: 215,069 km2. The biome abbreviations in the legend correspond to Table 3.
Floristic studies in the Sierra Madre Oriental. There is important information published on the floristic richness throughout the SMOR, which is useful to define and establish strategies to increase knowledge of its floristic diversity. Without a doubt, the synthesis of this information into a single checklist will facilitate its consultation and use. Salinas-Rodríguez (2015) synthesized a first approximation of the flora of the SMOR, reporting 7,748 species, distributed in 1,852 genera and 203 families. Recently Salinas-Rodríguez et al. (2022) updated the former data publishing a floristic checklist of the SMOR including 6,981 species. Additionally, botanical exploration has led to the publication of some 42 local floristic inventories carried out in the SMOR and neighboring regions that are sometimes considered part of the mountain range (Table 1). A summary of all the species cited in these inventories, once the scientific names used have been standardized, totals 5,993 species of vascular plants.
Table 1 Floristic inventories carried out in and around the Sierra Madre Oriental. State acronyms: COAH = Coahuila, HGO = Hidalgo, NLE = Nuevo León, PUE = Puebla, QRO = Querétaro, SLP = San Luis Potosí, TAMS = Tamaulipas, VER = Veracruz.
| State | Study region | Surface studied (Km²) |
Species | Reference |
|---|---|---|---|---|
| SMOR | Sierra Madre Oriental | 220,151.50 | 6,981 | Salinas-Rodríguez et al. 2022 |
| COAH | Municipio de Acuña | 11,451.90 | 367 | Villarreal-Quintanilla 2001 |
| COAH | Municipio de Ocampo | 25,894.90 | 847 | Villarreal-Quintanilla 2001 |
| COAH | Municipio de Saltillo | 6,429.40 | 1,060 | Villarreal-Quintanilla 2001 |
| COAH | Sierra de Jimulco | 625 | 339 | Alba-Ávila 2011 |
| COAH | Sierra de Parras | 95 | 692 | Rodríguez-Gámez et al. 1994 |
| COAH | Sierra de Zapalinamé | 450 | 934 | Encina-Domínguez et al. 2016 |
| COAH | Sierra de la Paila | 1700 | 778 | Villarreal-Quintanilla 1994 |
| NLE | Cañón de Iturbide | 422 | 698 | Salinas-Rodríguez et al. 2013 |
| NLE | Cerro Potosí | 4.3 | 130 | García-Arévalo & González-Elizondo 1991 |
| NLE | Municipio de Lampazos de Naranjo | 2,109 | 377 | Briones-Villarreal 1986 |
| NLE | Municipio de Santiago | 620.5 | 344 | Valdéz-Támez 1981 |
| NLE | Municipio General Zaragoza | 1118.7 | 312 | Treviño-Garza 1984 |
| NLE | Parque Nacional Cumbres de Monterrey | 1773.9 | 1,231 | Estrada-Castillón et al. 2013 |
| TAMS | ANP Altas Cumbres | 303.3 | 757 | García-Morales 2009 |
| TAMS | Región de Gómez Farías | 400 | 449 | Valiente-Banuet et al. 1995 |
| TAMS | Puerto Purificación | 136 | 68 | Treviño-Barbosa 2004 |
| TAMS | Sierra de Guatemala | 185.8 | 706 | Johnston et al. 1989 |
| TAMS | Sierra San Carlos | 1422.6 | 418 | Briones-Villarreal 1991 |
| SLP | Municipio de Guadalcázar | 3800 | 773 | Torres-Colín et al. 2017 |
| SLP | Sierra de Alvarez | 22.8 | 357 | García-Sánchez et al. 1999 |
| SLP | Sierra de Catorce | 739.1 | 494 | Giménez de Azcárate & González-Costilla 2011 |
| SLP | Sierra de Monte Grande | 41.8 | 386 | Reyes-Agüero et al. 1996 |
| SLP | Sierra del Abra-Tanchipa | 214.8 | 420 | De-Nova et al. 2019 |
| QRO | Cuenca del Río Estórax | 1,200 | 674 | Zamudio-Ruíz 1984 |
| QRO | Municipio de Landa | 742.1 | 728 | Cartujano et al. 2002 |
| QRO | Municipio de San Joaquín | 212 | 287 | Fernández-Nava & Colmenero-Robles 1997 |
| QRO | Sierra Gorda | 3830 | 1,402 | Arreguín-Sánchez & Fernández-Nava 2004 |
| HGO | Barranca de Metztitlán | 400.6 | 240 | CONANP 2003, Mendoza-Cariño & Quevedo-Nolasco 2019 |
| HGO | Barranca de Tolantongo | 10 | 287 | Hiriart-Valencia & González-Medrano 1983 |
| HGO | Eloxochitlán y Tlahuelompa | 27.2 | 375 | Alcántara-Ayala & Luna-Vega 2001 |
| HGO | Molocotlán | 13.3 | 351 | Mayorga-Saucedo et al. 1998 |
| HGO | Monte Grande, Lolotla | 253.1 | 349 | Ponce-Vargas et al. 2006 |
| HGO | Municipio de Tecozautla | 556.4 | 458 | Rojas et al. 2013 |
| HGO | Municipio de Tenango de Doria | 67.7 | 439 | Alcántara-Ayala & Luna-Vega 1997 |
| HGO | Municipio de Tlanchinol | 380.5 | 407 | Luna-Vega et al. 1994 |
| HGO | Sierra de Pachuca | 600 | 924 | Barrios-Rodríguez & Medina-Cota 1996 |
| HGO | Sierra Tezontlalpan | 216 | 386 | Equihua-Zamora 1983 |
| HGO | Sierra de Zacualtipan | 54.9 | 183 | Martínez-Jiménez 1995, Pérez-Paredes et al. 2012 |
| HGO | Valle del Mezquital | 2,773.10 | 566 | González-Quintero 1968, Soriano-Martínez & López-Soto 1984 |
| PUE | Xochitlán, Sierra Norte | 127.7 | 177 | Villalobos-Contreras 1994 |
| VER | Cofre de Perote | 1,300.80 | 834 | Nárave-Flores 1985 |
| VER | Municipio de Huayacocotla | 98.5 | 512 | Vargas-Ajuria 1982 |
| VER | Norte de Veracruz | 490 | 488 | Gutiérrez-Báez 1993 |
Another important source of information on plant species records are the databases of herbarium material incorporated into two digital collections that contain information on the richness of Mexican species. One is the National Biodiversity Information System (SNIB) of the National Commission for the Knowledge and Use of Biodiversity (CONABIO, www.conabio.gob.mx) and the other is the open data platform of the National Herbarium of Mexico (MEXU-UNIBIO: www.ibdata.ib.unam.mx), of the Institute of Biology of the National Autonomous University of Mexico (UNAM). The SNIB includes the information in digital format of more than 80 herbaria from both within and outside Mexico.
Combining the georeferenced records obtained from the query of the two databases and located within the polygon defined by Cervantes-Zamora et al. (1990) and the data obtained from the floristic inventories, a list of 8,453 species that constitute the diversity of vascular plant species of the SMOR has been compiled (Villaseñor, unpublished data available upon request). This figure slightly exceeds the number of species reported by Salinas-Rodríguez (2015), probably because new information is continuously accumulated in these living databases and because different criteria on local floristic inventories were considered. This broad floristic diversity in the SMOR is a mixture of species that have evolved in parallel with the development of the arid and semi-arid zones of the Mexican Highland (Chihuahuan Desert) due to the Pleistocene glaciations, along with older elements associated with the humid mountain or temperate forests that characterize the typical roughness of the SMOR.
This study analyzes the geographical distribution of a particular set of species, hereafter referred to as “characteristic species” (with half or more of their known distribution range in Mexico inside the SMOR, see materials and methods), with the aim of identifying patterns of species richness and locating the main centers of floristic richness of these characteristic species. Once the patterns were identified, our second objective was to propose a regionalization of the SMOR, determining species that constitute its floristic elements and evaluating species shared between their biogeographic regions (phytochorions) identified.
Materials and methods
Study area. The Sierra Madre Oriental (SMOR) is recognized as a physiographic province of Mexico (Cervantes-Zamora et al. 1990, Espinosa et al. 2004, Salinas-Rodríguez et al. 2017). According to Cervantes-Zamora et al. (1990) it covers an area of 215,069 km2 that includes part of the states of Coahuila (45 % of the SMOR surface), Durango (3 %), Guanajuato (1 %), Hidalgo (4 %), Nuevo León (15 %), Puebla (2 %), Querétaro (2 %), San Luis Potosí (16 %), Tamaulipas (6 %), Veracruz (2 %), and Zacatecas (5 %).
Physiographically it is made up of a series of mountain ranges and valleys, including the Sierras y Llanuras Coahuilenses, the Serranía del Burro and the Sierra de la Paila, as well as the Sierra de Saltillo-Parras in Coahuila, and the Sierras Transversales, which run perpendicular to the main range, from northeastern Durango to northern Zacatecas. Equally important are the Sierras y Llanuras Occidentales between Nuevo León and San Luis Potosí, the Gran Sierra Plegada that runs from Nuevo León and Tamaulipas to San Luis Potosí, and finally the Carso Huasteco, at the southern end of the SMOR in the states of San Luis Potosí, Querétaro, Hidalgo, Puebla, and Veracruz (Espinosa et al. 2004). Within the SMOR, type A (tropical) climates are recorded in 7 % of the surface, type B (dry) in 78 % of the territory and type C (temperate) in 15 % of its surface (García 1998).
The soils type is Leptosol in 50 % of the SMOR, which are shallow and stony soils that can have continuous rock at or very close to the surface; leptosols are found in all types of climates (humid, dry, or temperate) and are particularly common in mountainous areas and shallow limestone plains. Calcisols occupy another 33 % of the territory and are soils with a substantial accumulation of calcareous material. They are widespread in arid and semi-arid environments and mainly come from alluvial, colluvial and aeolian deposits of weathered material. Other soil types found in lower proportions of the SMOR include Regosols (8 %), Faeozems (4 %), Cambisols (1 %), and Andosols (1 %) (SEMARNAP 1998). The biome (Villaseñor & Ortiz 2014) that occupies the largest proportion of the SMOR is xerophilous scrub (64 %), followed by temperate forest (23 %), seasonally dry tropical forest (6 %), humid tropical forest (5 %) and humid mountain forest (2 %).
Database and analysis. The data analyzed here were gathered from two main sources; the first is the published information on species inventories (Table 1) and the second is the georeferenced records available in the SNIB and MEXU-UNIBIO electronic databases (see introduction). All georeferenced points included inside the polygon defined by Cervantes-Zamora et al. (1990) were considered. As mentioned previously, the inventories listed a total of 5,993 species; after review, cleaning, and curation of more than 300,000 database records, this total increased to 8,472 species. Species nomenclature follows Villaseñor (2016).
Of the set of 8,472 identified vascular plant species that constitute the flora of the SMOR area as delimited by Cervantes-Zamora et al. (1990), we defined a set of “characteristic species”. To do this, we divided Mexico into 253 grid cells of 1° latitude and longitude (Figure 4), then determined the squares or grid cells that contain the SMOR polygon (a total of 42 squares; Figure 4). We considered a species to be "characteristic of the SMOR" if 50 % or more of its geographic range in Mexico (considered as the total grid cells counted) is included in the 42 grid squares that overlap the SMOR polygon (as defined by Cervantes-Zamora et al. 1990). For example, Fraxinus greggii A. Gray (Oleaceae) was recorded in 47 grid cells throughout Mexico, 31 of which overlap the SMOR, qualifying as a characteristic species of the SMOR. By contrast, Esenbeckia berlandieri Baill. ex Hemsl. (Rutaceae) has been recorded in 57 grid cells in Mexico, but only 15 of those fell within the SMOR, so it did not qualify as a characteristic species. To determine the characteristic species, we calculated the number of grid cells in Mexico and in the SMOR where each of the 8,472 species had been recorded. The incidences per square were used to generate maps of richness and endemicity, determining both alpha diversity values (richness per grid cell), as well as the richness distribution using the Universal Kriging interpolation technique in a Geographic Information System (GIS) (ESRI 2013).

Figure 4 A. Division of Mexico in grid cells of 1° latitude and longitude. B. Detail of the Sierra Madre Oriental indicating the grid cells it includes.
An incidence matrix was used to evaluate the floristic similarities between grid cells that include the SMOR. Pairwise similarities were determined using the Sørensen-Dice similarity coefficient (SD= 2c/a+b+2c), where c is the number of species shared between two grid cells, a is the number of exclusive species in one square and b the exclusive species in the other square. The similarity matrix was transformed into a dendrogram to illustrate the floristic similarities between squares, using the weighted grouping method using arithmetic means (WPGMA). This method weights the contributions of the clusters considering the number of grid cells they contain, ensuring that each of them contributes equally to each nesting of which they are part (González-Orozco et al. 2014). The analyses were carried out using the NTSYS-pc program, version 2.21 (Rohlf 2007).
Results
As previously indicated, considering the polygon that delimits the SMOR as proposed by Cervantes-Zamora et al. (1990), Figure 1), the vascular flora of the SMOR contains a total of 8,472 species. Of this set, 3,433 were identified as "characteristic" species (Supplementary material), a figure that represents 40.5 % of the total floristic richness recorded. Of these, 2,158 are endemic to Mexico and 585 are considered strictly endemic to the SMOR territory. Interestingly, 77 of the characteristic species were exotic (introduced) species (see, Supplementary material).
Figure 5 illustrates the national level distribution of the 3,433 characteristic species of the SMOR. Most of them concentrate their distribution on the territory occupied by the polygon of the SMOR, surrounded by an important area where most species extend their distribution range.

Table 2 shows the distribution of the floristic richness in the SMOR among the states that contain SMOR. The number of characteristic species within each state is also indicated. The states of Coahuila, Nuevo León, Tamaulipas, and San Luis Potosí had particularly high richness of characteristic species. The number of characteristic species gradually decreased further south, with the lowest numbers of them in the states of Guanajuato, Puebla, and Zacatecas. It should be noted that several of the states that included low numbers of characteristic species contain only a small area of SMOR in their territory, for example Durango, Guanajuato, or Zacatecas.
Table 2 Richness of vascular plant species of the Sierra Madre Oriental (SMOR) in each of the states that contain SMOR territory.
| State | SMOR Species | Characteristic species |
Mexican endemic characteristic species |
SMOR-endemic species |
|---|---|---|---|---|
| Coahuila | 3,630 | 2,039 | 970 | 209 |
| Durango | 928 | 540 | 177 | 1 |
| Guanajuato | 2,478 | 709 | 438 | 40 |
| Hidalgo | 4,743 | 1,314 | 844 | 121 |
| Nuevo León | 3,761 | 2,022 | 1,072 | 342 |
| Puebla | 4,372 | 711 | 384 | 32 |
| Querétaro | 4,222 | 1,289 | 848 | 157 |
| San Luis Potosí | 5,125 | 1,867 | 1,136 | 217 |
| Tamaulipas | 3,860 | 1,565 | 871 | 271 |
| Veracruz | 5,268 | 1,133 | 669 | 66 |
| Zacatecas | 1,430 | 696 | 303 | 6 |
| SMOR Total | 8,472 | 3,433 | 2,158 | 585 |
Endemism in the Sierra Madre Oriental. A total of 585 species were recognized as SMOR characteristic endemics. The states of San Luis Potosí, Nuevo León, Coahuila, Tamaulipas, and Hidalgo record more than 800 species endemic to Mexico (Table 2). This order is slightly changed when the number of endemic restricted to the SMOR are considered; in this case Nuevo León, Tamaulipas, San Luis Potosí, and Coahuila occupy the most important positions (Table 2).
Distribution of characteristic species by biome. Throughout the SMOR, no grid cell was occupied by a single biome; four grid cells contained two biomes, 29 cells contained three biomes, five contained four biomes, and four cells contained all five main biomes. Table 3 indicates the number of grid cells in which each biome was recorded, and the total area occupied. It is noteworthy that xerophilous scrub (XES) comprised most of the territory of the SMOR, even though this biome is not considered typical of mountainous regions.
Table 3 Main biomes (sensu Villaseñor &Ortiz 2014) recorded throughout the Sierra Madre Oriental (SMOR) as circumscribed by Cervantes-Zamora et al. (1990); number of grid cells where the biome is recorded, number of species characteristic of the SMOR and total area occupied.
| Biome | Grid cells where it is recorded |
Characteristic species | Surface (km2) |
|---|---|---|---|
| Humid mountain forest (HMF) | 9 | 750 | 5,246.5 |
| Temperate forest (TEF) | 41 | 1,796 | 49,055.5 |
| Humid tropical forest (HTF) | 8 | 140 | 10,308.6 |
| Seasonally dry tropical forest (SDTF) | 37 | 390 | 12,328.5 |
| Xerophilous scrub (XES) | 40 | 2,029 | 138,051.4 |
Since mountainous regions are generally considered to be characterized by humid mountain or temperate forests, it is important to analyze how much of the xerophilous scrubs recorded should be considered legitimately part of the SMOR versus constituting erroneously included components of neighboring biogeographic provinces. Similar arguments must be made with respect to the location of the wet or seasonally dry tropical forests, characteristic of the lowlands located especially on the windward side of the sierra.
Table 3 shows roughly equivalent values of species richness between temperate forests (TEF) and xerophilous scrub (XES), distributed in an almost equal number of grid cells but with a contrasting surface area. It is surprising to observe that this region that is considered eminently mountainous (SMOR) is dominated by arid or semi-arid territories that are not generally considered typical mountain ecosystems.
Species richness by SMOR grid cell.Table 4 indicates the number of species (alpha diversity) and their incidences by biome recorded in each grid cell that comprises the SMOR. Virtually all grid cells included many species classified either as elements of temperate forests or xerophilous scrub, which suggests either that these two biomes are found almost sympatrically throughout the SMOR, or that the floristic composition tolerates both climates where they occur. A highly significant correlation between the species of both biomes is found throughout the SMOR (R = 0.905, P <0.05).
Table 4 Grid cells (1° latitude by 1° longitude; 42 cells total) into which the territory of the Sierra Madre Oriental (SMOR) was divided, the number of characteristic species and the number of species recorded per biome. Grid cell numbers correspond to Figure 3 and biome abbreviations correspond to Table 3. Empty values indicate the biome is absent in the grid cell. Differences in total species per grid cell and species by biome indicate lack of information in the available records.
| Grid cell | Species | HMF | TEF | HTF | SDTF | XES |
|---|---|---|---|---|---|---|
| 044 | 471 | 454 | ||||
| 045 | 757 | 661 | ||||
| 046 | 354 | 306 | ||||
| 061 | 337 | 206 | 322 | |||
| 062 | 834 | 496 | 724 | |||
| 063 | 396 | 220 | 366 | |||
| 064 | 225 | 122 | 207 | |||
| 078 | 428 | 416 | ||||
| 079 | 685 | 409 | 631 | |||
| 080 | 505 | 256 | 457 | |||
| 081 | 350 | 205 | 322 | |||
| 092 | 476 | 220 | 457 | |||
| 093 | 716 | 400 | 687 | |||
| 094 | 954 | 532 | 887 | |||
| 095 | 677 | 369 | 611 | |||
| 096 | 193 | 107 | 176 | |||
| 107 | 199 | 128 | 191 | |||
| 108 | 525 | 301 | 493 | |||
| 109 | 636 | 361 | 596 | |||
| 110 | 1,088 | 643 | 936 | |||
| 111 | 1,579 | 289 | 932 | 1,122 | ||
| 112 | 379 | 91 | 228 | 297 | ||
| 124 | 181 | 97 | 175 | |||
| 125 | 263 | 251 | ||||
| 126 | 479 | 307 | 442 | |||
| 127 | 1,059 | 691 | 152 | 759 | ||
| 128 | 1,183 | 285 | 755 | 213 | 803 | |
| 139 | 414 | 293 | 380 | |||
| 140 | 624 | 422 | 547 | |||
| 141 | 1,359 | 390 | 888 | 75 | 256 | 876 |
| 142 | 345 | 207 | 44 | 133 | 274 | |
| 152 | 962 | 237 | 632 | 192 | 785 | |
| 153 | 536 | 186 | 334 | 48 | 167 | 411 |
| 154 | 232 | 122 | 141 | 168 | ||
| 162 | 667 | 164 | 441 | 150 | 558 | |
| 163 | 1,153 | 470 | 753 | 107 | 279 | 737 |
| 16 | 375 | 207 | 240 | 72 | 135 | |
| 177 | 1,061 | 345 | 696 | 58 | 230 | 820 |
| 178 | 987 | 471 | 685 | 87 | 206 | 621 |
| 179 | 197 | 147 | 124 | 59 | 64 | 76 |
| 194 | 450 | 215 | 351 | 88 | 270 | |
| 195 | 577 | 341 | 439 | 71 | 124 | 292 |
| SMOR Total | 3,433 | 750 | 1,796 | 140 | 390 | 2,029 |
Grid squares 111 and 141 had the highest alpha diversity, both in the number of characteristic species and in the number of species of considered montane biomes. Other important grid cells with more than 1,000 recorded species were cells 110, 127, 128, and 177. All these cells had high values of alpha diversity as well as species that occupy different biomes.
Floristic similarities and regionalization of the SMOR. It is plausible that the 3,433 characteristic species of the SMOR share a common biogeographic history since their known distribution is strongly linked to the SMOR and its area of influence (Figure 5). Figure 6 illustrates the floristic similarities between the grid squares containing SMOR territory after applying the Sørensen-Dice similarity coefficient and a weighted grouping method (WPGMA).

Figure 6 Floristic similarities between the grid cells that include the Sierra Madre Oriental (Cervantes-Zamora et al. 1990). The dendrogram was generated using the Sørenson-Dice similarity coefficient and the WPGMA grouping method. Numbers as terminals correspond to the Figure 4.
The dendrogram in Figure 6 revealed five main groups that show a consistent geographic structure throughout the SMOR territory (Figure 7). Four grid cells (107, 124, 142, and 154) grouped inconsistently in the dendrogram of Figure 6; they are shown as outliers of better-consolidated geographic groups. Accordingly, they were visually assigned to the floristic regions that were closest to their geographical position.

Figure 7 A. Geographical arrangement of the floristic districts of the Sierra Madre Oriental. B. Comparison of floristic districts with the regionalization proposed by Morrone (2019).
Because they contain more than two endemic species within their limits, three of these floristic units constitute areas of endemism and could serve to regionalize the SMOR into phytogeographic districts (Table 5). District 2 (located in the extreme northeast of the sierra) and District 4 (central-eastern part of the sierra; Figure 7), did not have exclusive endemic species. Unlike the other districts, the floristic composition of these two districts was predominantly made up of species that are ecologically related to xerophilous scrubs. Thus, future studies will likely place them preferentially in neighboring biogeographic provinces that are more dominated by xerophilous shrubs, e.g., the Tamaulipas province for District 2 and the Southern Altiplano province (Zacatecano-Potosino) for District 4, as defined by (CONABIO 1997, see also Villaseñor et al. 2020).
Table 5 Characteristic species of the Sierra Madre Oriental (SMOR) from each of the five floristic districts (FD) within the SMOR and the biomes where they are recorded. The number of species endemic to the SMOR and to each floristic district is also indicated. Empty values indicate the biome is absent in the floristic district.
| FD 1 | FD 2 | FD 3 | FD 4 | FD 5 | Total | |
|---|---|---|---|---|---|---|
| Total characteristic species | 1,624 | 875 | 2,381 | 992 | 2,178 | 3,433 |
| Characteristic species exclusive to the FD | 138 | 4 | 413 | 7 | 732 | 1,294 |
| SMOR-endemic species | 54 | 14 | 392 | 42 | 253 | 585 |
| SMOR-endemic species exclusive to the FD | 8 | 0 | 257 | 0 | 160 | 425 |
| HMF | 468 | 690 | 750 | |||
| TEF | 843 | 465 | 1,315 | 615 | 1,261 | 1,796 |
| HTF | 86 | 136 | 140 | |||
| SDTF | 310 | 369 | 390 | |||
| XES | 1,355 | 756 | 1,548 | 864 | 1,339 | 2,029 |
| Grid cells that include the FD | 44, 45, 61, 62, 78, 79, 92, 93, 94, 95, 107, 108, 109 |
46, 63, 64, 80, 81, 96 |
110, 111, 112, 127, 128, 141, 142 |
124, 125, 126, 139, 140 |
152, 153, 154, 162, 163, 164, 177, 178, 179, 194, 195 |
Table 5 indicates the number of characteristic species and the number of SMOR-endemic species recorded in each phytochorion (District). The number of endemic species that are restricted to the grid cells that make up each district is also indicated. Districts 3 and 5 had the highest number of restricted endemics, as well as a high proportion of species typical of temperate environments, such as humid mountain forest (HMF) or temperate forest (TEF).
Worldwide distribution of the SMOR characteristic species.Table 6 indicates the distribution around the world of the 3,433 species considered here to be characteristic of the SMOR. The Mexican endemic component stands out, followed by North American element. Contrary to expectations, the South American element (Neotropical) is poorly represented among the characteristic species.
Table 6 Geographical distribution of the 3,433 characteristic species of the Sierra Madre Oriental. The floristic element refers to the continental part at worldwide scope where the species is located.
| Geographical distribution | Floristic element | Species |
|---|---|---|
| Mexican Endemic (species endemic to Mexico) | Endemic | 2,158 |
| SMOR-endemic (species endemic to the SMOR) | Endemic | 585 |
| North America to Mexico | North American | 942 |
| North America to Central America | North American | 39 |
| Mexico to Central America | Mesoamerican | 73 |
| Mexico to South America (including the Caribbean Islands) | South American | 46 |
| North America to South America | American | 55 |
| Distributed also in the Old World | Cosmopolitan | 71 |
| Uncertain distribution | 29 |
Discussion
The SMOR as delimited by (Cervantes-Zamora et al. 1990, Figure 1) has higher floristic richness than any other mountainous region of Mexico. Its 8,472 species identified in this work exceed the 6,981 species of the SMOR flora reported by Salinas-Rodríguez et al. (2022), and exceeds preliminary estimates for other important mountain ranges (Villaseñor, unpublished data), including the Trans-Mexican Volcanic Belt (6,751 species), the Sierra Madre Occidental (6,628) and the Sierra Madre del Sur (5,874). A progressive decrease of the geographical ranges of the characteristic species is observed, with sites further away from the SMOR recording low species numbers, showing the typical decline in floristic similarity discussed in ecology (Nekola & White 1999).
Although it is considered mainly as a usually mountainous province, it includes a mixture of elements typical of the five main biomes of Mexico (Villaseñor & Ortiz 2014). The temperate and humid mountain biomes predominate, but xerophilous scrub is also strongly represented. Examples of species known mainly in the humid tropical forests of the coastal plain of the Gulf of Mexico that penetrate the lower parts of the SMOR include Eugenia huasteca E. Sánchez-Chávez & Zamudio (Myrtaceae), Philodendron subincisum Schott (Araceae), Platymiscium calyptratum M. Sousa & Klitg. (Fabaceae) or Zephyranthes moctezumae T.M. Howard (Amaryllidaceae). On the other hand, there are elements that are more associated with seasonally dry tropical forests, such as Beaucarnea glassiana (L. Hern. & Zamudio) V. Rojas (Asparagaceae), Cleome potosina B.L. Rob (Cleomaceae), Batesimalva pulchella Fryxell (Malvaceae) or Rhynchosia rosaurae Rzed. (Fabaceae). As mentioned above, an important component of SMOR is made up of the species predominantly found in xerophilous scrub, such as Dasylirion miquihuanense Bogler (Asparagaceae), Jaimehintonia gypsophila B.L. Turner (Asparagaceae), Amauriopsis janakosana B.L. Turner (Asteraceae), Cynoglossum henricksonii L.C. Higgins (Boraginaceae), Rapicactus mandragora (Frič ex A.Berger) Buxb. & Oehme (Cactaceae) or Senegalia sororia (Standl.) Britton & Rose (Fabaceae).
More than 1,200 of the characteristic Mexican-endemic species were found exclusively in the SMOR and neighboring provinces (Altiplano and Gulf of Mexico Coast), which underscores the need to assess the boundaries between them. Probably many of the species currently considered characteristics of the SMOR belong to one of these neighboring provinces, especially those adapted to the dry environments typical of xerophilous scrub or transition zones.
The species we considered characteristic of the SMOR constitute a representative set of the component that has diversified mainly in this mountain range. This is evidenced in Figure 5, which shows that their distribution is centered on this mountain range, with a decline in the number of species recorded with increasing distance from the province. It is likely that many of these species have their centers of origin within this mountain range, which could be verified by future studies on their biogeography and relationships with other species in their respective genera.
The states of San Luis Potosí and Veracruz had the highest total number of species among the 11 states whose territories include the SMOR (Table 2). However, Coahuila, Nuevo León, and San Luis Potosí, had particularly high numbers of characteristic species. Examples of these characteristic species endemic to the SMOR, and also endemic to Coahuila or Nuevo León, include Ageratina riskindii B.L. Turner (Asteraceae), Astragalus regiomontanus Barneby (Fabaceae), Dasylirion micropterum Villarreal, A.E. Estrada & Encina (Asparagaceae), Juniperus zanonii R.P. Adams (Cupressaceae), Leucophyllum alejandrae G.L. Nesom (Scrophulariaceae), Philadelphus sargentianus S.Y. Hu (Hydrangeaceae), Pinguicula bustamanta Zamudio & Nevárez (Lentibulariaceae) or Scutellaria fraxinea Epling (Lamiaceae). These are only a few examples among the 585 species identified as characteristic and endemic to the SMOR.
The state of San Luis Potosí apparently constitutes a boundary between two floristically relevant portions of the SMOR. The physiographic provinces of the Gran Sierra Plegada and the Sierras y Llanuras Occidentales converge in the northern part of San Luis Potosí, which constitutes Districts 3 and 4 identified in this work (Table 5, Figure 6). Meanwhile the southern portion of the state comprises the Carso Huasteco physiographic province that extends throughout the southern end of the SMOR until its contact with the Neovolcanic Axis (Trans-Mexican Volcanic Belt).
In the northern part of the SMOR, Coahuila, Nuevo León and Tamaulipas have the largest number of endemic species characteristic of the SMOR (Table 2). Towards its southern part, no state dominates by its number of characteristic species; however, it is the SMOR’s portion where the highest percentage of tropical elements, characteristic of the humid mountain and tropical forests predominate (either seasonally dry or humid tropical forests).
Sierra Madre Oriental endemisms. The study of the endemic vascular plant species of the SMOR has attracted the attention of several botanists. Rzedowski (2015) published a first list of species whose distribution is mostly restricted to the SMOR; this author cites some 659 species, although only 573 of those were identified as endemic in the current study. Some of the species recognized as endemic by Rzedowski have been recorded from locations outside the limits of the SMOR, which is why they were considered non-endemic here. Later, Salinas-Rodríguez et al. (2017) reported a figure of 1,135 species endemic to the SMOR, updated to 1,542 in Salinas-Rodríguez et al. (2022): but likewise, many of the species considered endemic were excluded in this work because there are also records of collecting localities outside the limits of the province considered here.
There are reports of endemic species from Coahuila (Villarreal-Quintanilla & Encina-Domínguez 2005), Hidalgo (Villaseñor et al. 2022), Nuevo León (Alanís-Flores et al. 2011), Puebla (Rodríguez-Acosta et al. 2014) and San Luis Potosí (De-Nova et al. 2018); all these counts include state endemics distributed along the sierra, although sometimes not pointing out those restricted to the SMOR. Table 2 summarizes the number of endemic species in each of the 11 states that constitute the SMOR.
Many species reported as endemic were identified as shared elements with other neighboring biogeographic regions. For example, 1,242 of the characteristic and Mexican endemic species were shared between the SMOR and the Altiplano Province; they were therefore not considered SMOR endemics here, although many of them have been reported as SMOR endemics by other authors. The case was similar for 242 species known only from the SMOR and the Gulf of Mexico Coast province.
Other studies on the endemism of areas geographically related to the SMOR include species distributed along the SMOR, sometimes making it difficult to locate them precisely in the right region. One is the Chihuahuan Desert (Bartolomé-Hernández 2015, Villarreal-Quintanilla et al. 2017), which is part of the Altiplano Province (CONABIO 1997) that borders the SMOR on its western flank, and another is the Trans-Mexican Volcanic Belt (Eje Volcánico, Rzedowski 2020) that borders the SMOR to the south. Examples of the difficulty in the precision of endemism include Haploesthes fruticosa B.L. Turner and Psathyrotopsis hintoniorum B.L. Turner (both members of the Asteraceae family) or Turbinicarpus valdezianus (Moell.) Glass & R. Foster (Cactaceae), which are recognized as endemic species of the Chihuahuan Desert by Villarreal-Quintanilla et al. (2017) but also as endemic to the SMOR by Salinas-Rodríguez et al. (2017). In summary, considering 8,472 species as the SMOR flora and the standardized nomenclature of the species cited in the works on endemism cited above, results in a list of 585 species that are strict endemics of the SMOR. This figure is lower than that reported by Rzedowski (2015) and less than half of the species considered endemic by Salinas-Rodríguez et al. (2017).
Floristic richness by biome and physiographic subprovince. The complete lack of grid cells containing a single biome and the existence of cells containing all five major biomes indicates the high environmental heterogeneity throughout the area occupied by the SMOR. Since biomes are defined mainly based on the climatic and floristic composition, the presence of several biomes within the same territory is inextricably linked to the high alpha richness found in most of the grid cells into which the SMOR was divided (Table 4).
It is not easy to delimit the SMOR as proposed by Cervantes-Zamora et al. (1990) based solely on floristic composition. Numerous grid cells included in the polygon contain mostly temperate and xerophytic biomes, the latter of which is not a typical element of mountainous regions. The limits proposed by CONABIO (1997, 2008) seem to provide a closer circumscription; however, it excludes several grid cells with temperate forests to the north of the proposed boundaries that are undoubtedly typical components of mountainous regions. For example, the CONABIO polygon (1997, 2008) does not include typically mountainous areas such as the Serranías del Burro or the Sierra Maderas del Carmen: two examples of the more than 12 isolated mountain ranges that characterize District 1 (Table 5), located in the extreme northwest of the SMOR.
The complex physiography of the SMOR makes it difficult to propose precise phytogeographic units, as revealed by the different biogeographic studies carried out in the sierra (see below). The SMOR is subdivided into eight physiographic subprovinces (INEGI 1991) and except for the Carso Huasteco subprovince which corresponds exclusively to District 5 of the regionalization proposed here, the territory of the other subprovinces is divided among two or more districts. The subprovinces of the Sierras y Llanuras Coahuilenses and Serranía del Burro are immersed in District 1. These subprovinces have isolated mountain ranges, but they predominantly contain elements of the xerophilous scrub biome that extends continuously into the neighboring subprovinces that are not part of the SMOR. These include the Llanuras de Coahuila and Nuevo León (Grandes Llanuras de Norteamérica province) to the north or the Llanuras y Sierras Volcánicas subprovince (Sierras y Llanuras del Norte province) to the south.
District 3 includes the subprovinces of the Sierra de la Paila and that of the Pliegues Saltillo-Parras, but it also includes part of the subprovinces of the Sierras Transversales (shared with District 4), of the Gran Sierra Plegada (shared with District 5) and the Sierras y Llanuras Occidentales (shared with Districts 4 and 5). All these physiographic subprovinces form a topographic complex where many species are distributed continuously, none of them exclusively found in just one subprovince.
Biogeographic regionalization of the SMOR physiographic province. There are proposals for the regionalization of the SMOR using different data sets from plants or animals. For vascular plants, none of these proposals addresses the analysis of the full floristic diversity of the province. Morrone (2019) provides a synthesis of the works that have discussed biogeographic aspects that are relevant to the regionalization of the SMOR, and the approaches used. His synthesis makes evident the paucity of studies involving plant species, since it only mentions as examples of endemic species or characteristic of the sierra nine species of vascular plants, of which only three are included in this work as characteristic species.
Morrone (2019), in his analysis of the different biogeographic proposals of the SMOR regionalization summarizes that the province can be divided into two subprovinces and four districts. However, the polygon used in that study is much narrower than that considered by Cervantes-Zamora et al. (1990), leaving out important portions to the north of its limits which in the present study constitute Districts 1 and 2, which mainly comprise the physiographic subprovinces of the Sierras y Llanuras Coahuilenses and the Serranía del Burro (Figure 7, Table 5).
The Austral-Oriental Subprovince of Morrone (2019) comprises the Saltillo-Parras and Potosí Districts. The Saltillo-Parras District mainly includes the physiographic subprovinces of the Gran Sierra Plegada, the Pliegue Saltillo-Parras and the Sierras Transversales, here considered part of Districts 1, 3 and 4. The Potosí district comprises mainly part of the Gran Sierra Plegada subprovince and part of the Sierras y Llanuras Occidentales subprovince; in this work, Morrone’s Potosí District corresponds in part with Districts 3, 4 and 5 (Figure 7).
The Hidalguense Subprovince of Morrone (2019) also includes two districts (Sierra Gorda and Zacualtipán), that in this work were considered a single phytochorion within the physiographic subprovince of Carso Huasteco and characterizing District 5. Floristically, District 5 constitutes one homogeneous unit, although with high species turnover values at the level of local inventories. The high floristic diversity of the district identifies it as a phytogeographic unit, although specific locations within the district do not necessarily share the same species. Examples were shown by Salinas-Rodríguez (2015), who, for example, also identifies the entire Carso Huasteco as a phytogeographic unit, but with low floristic similarities between the Sierra Gorda in Guanajuato and in Querétaro.
The identified floristic regions (Districts) are delimited by a set of species with similar geographic distribution within them (Table 5); several of these floristic elements are even endemic to the district, elevating those districts to the status of areas of endemism. For example, District 1 contains eight exclusive endemic species (Ageratina riskindii B.L. Turner, Berlandiera burroana B.L. Turner, Brickellia wendtii B.L. Turner, Erigeron vicinus G.L. Nesom, Nama constancei J.D. Bacon, Phemeranthus calycinus (Engelm.) Kiger, Phlox pattersonii Prather, and Stevia burrana B.L. Turner), while districts 3 and 5 contain 257 and 160 exclusive endemic species, respectively. Only Districts 2 and 4 had no endemic species restricted to their territory (Table 5), although District 2 contains four characteristic species, in the SMOR distributed exclusively in their territory (Euphorbia georgei Oudejans, Linum berlandieri Hook., Pediomelum humile Rydb. and Symphyotrichum drummondii (Lindl.) G.L. Nesom) while District 4 records seven species (Agave oroensis Gentry, Apodanthera cucurbitoides Lundell, Halimolobos pedicellata (Rollins) Rollins, Hedeoma rzedowskii B.L. Turner, Lycium barbinodum Miers, Opuntia chaffeyi Britton & Rose, and Sedum catorce G.L. Nesom). The influence of the Altiplano province is manifested in these two districts, since all the SMOR characteristic species distributed exclusively in their territory are shared with this province, characterized by its arid and semi-arid environments where xerophilous scrubs predominate (Figure 8).

Figure 8 Distribution of the main biomes of Mexico in the floristic districts of the Sierra Madre Oriental. The biome abbreviations are the same as in Table 3.
Regional endemism becomes evident when evaluating endemic SMOR species shared between neighboring districts. For example, 26 species are shared between Districts 1 and 3 (Ageratina gypsophila B.L. Turner, Allium hintoniorum B.L. Turner, Astranthium ciliatum (Raf.) G.L. Nesom, Baccharis crassicuneata G.L. Nesom, Bidens carpodonta Sherff, Castilleja galehintoniae G.L. Nesom, Coryphantha pseudoechinus Boed., Crataegus cuprina J.B. Phipps, Flyriella leonensis (B.L. Rob.) R.M. King & H. Rob., Geranium potosinum H.E. Moore, Hedeoma oblatifolia Villarreal, Hedeoma quercetora Epling, Hieracium gypsophilum B.L. Turner, Lithospermum nelsonii Greenm., Mammillaria glassi R.A. Fosteri, Mammillaria melanocentra Posel., Pinus stylesii Frankis ex Businskyi, Pleopeltis fallacissima (Maxon) A.R. Sm. & Tejero, Pseudognaphalium hintoniorum (G.L. Nesom) Hinojosa & Villaseñor, Rapicactus mandragora (Frič ex A.Berger) Buxb. & Oehme, Salvia potus Epling, Senecio carnerensis Greenm., Seymera tamaulipana B.L. Turner, Solidago hintoniorum G.L. Nesom, Thelypodium lobatum Brandegee and Verbesina daviesiae B.L. Turner). On the other hand, four species are shared between Districts 2 and 3 (Anoda leonensis Fryxell, Koanophyllon reyrobinsonii B.L. Turner, Notholaena brachycaulis Mickel and Sedum diffusum S. Watson), while 54 species are shared between Districts 3 and 5. (Supplementary material) includes the grid cells where species distribute, and Table 5 indicates to which district the grid cells are assigned.
The SMOR is located within the Mexican Transition Zone (Morrone 2019), a point of view supported by numerous authors due to the mixture of elements from different biogeographic kingdoms; for plants in particular this condition is supported at the genus level (Villaseñor et al. 2020). However, by its characteristic species, it becomes evident that the North American floristic kingdom has a stronger influence than the Neotropical kingdom. In the SMOR 942 characteristic species are distributed from Mexico to North America, while only 55 species are distributed from Mexico to South America (Table 6). Although many genera constituting the SMOR flora show a Neotropical affinity, their species by geographic distribution are mostly endemic at the national or regional level.
Considering our results, it is evident that there continues to be a lack of academic consensus on the delimitation of the SMOR province in biogeographic, floristic (see Salinas-Rodríguez et al. 2022), and physiographic terms. The evidence analyzed highlights the need to carry out similar analyses in neighboring provinces, including the Altiplano province to the west and the Gulf of Mexico Coast to the east. Only a combined analysis will make it possible to undoubtedly discriminate the characteristic species of each one and thus more clearly define their biogeographic limits. The high percentage of species that is shared between the SMOR and the Altiplano province, especially those that are preferentially distributed in xerophilous scrub, requires a more precise discrimination of which elements are typical of mountainous areas and which belong more properly to the plains that do not meet the ruggedness criteria to be considered montane elements.
As a result of the differences found between this work and Morrone’s (2019) proposal, it is preferable not to propose a nomenclature for each recognized district (Figure 7). Morrone’s synthesis (2019) critically summarizes the nomenclatural proposals applied to the biogeographic subdivisions of the SMOR, showing the great diversity of names used. It also highlights the important differences found among various studies, especially due to the use of different biological elements which surely have different evolutionary histories. Despite the maxim that biota and land evolve together, at finer scales within the SMOR, the in situ evolution of endemic species is not exclusively due to historical factors, but also to species ecology and environmental conditions. Probably this results in contrasting biogeographic units when different taxonomic groups are analyzed.
Combining data from both plants and animals (see study cases in Morrone 2019) would help reach a hypothesis about the biogeographic similarities between the different units into which the province is subdivided. In this way, a unified classification of the biogeographic scenario of Mexico would be achieved, as proposed by Villaseñor et al. (2020).
Supplementary material
Supplemental data for this article can be accessed here: https://doi.org/10.17129/botsci.3086

