











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Germination occurs when seeds encounter suitable environmental conditions of temperature, humidity and light for their development; however, the nature of the seed itself and internal dormancy can inhibit germination, even in favorable environmental conditions (Finch-Savage & Leubner-Metzger 2006, Holdsworth et al. 2008). Besides the obstacles for germination, the species may face difficulties for establishment, such as Magnolia vovidesii A. Vázquez, Domínguez-Yescas & L. Carvajal belonging to Magnolia L. sect. Macrophylla Figlar & Noot., having high seed mortality (99.7 %) and 96.7 % mortality in the transition from seedlings to juveniles, resulting in low regeneration potential (Galván-Hernández et al. 2020).
Seeds have developed different strategies that enable them to survive adverse environmental conditions. For example, some seeds conserve energy to maintain the integrity of their genetic information and prolong their viability for several days, months, or even years (Corral-Aguirre & Sánchez-Velázquez 2006, Kisanuki et al. 2008), establishing a demographic balance in the population (Simpson et al. 1989); meanwhile, others germinate immediately when environmental conditions are right (Long et al. 2015). These strategies, whether alone or in combination, are called seed dormancy and are directly related and antagonistic to the plant life cycle (Black et al. 2006, Finch-Savage & Leubner-Metzger 2006, Bewley et al. 2013).
Sexual reproduction could result in a bottleneck when the seeds are unviable or dormant (Venable & Brown 1988). Some species of the genus Magnolia exhibit morphological or physiological dormancy, or a combination of both (Baskin & Baskin 2014). For example, some species present embryos that are not fully developed at the time of fruit dehiscence, suggesting physiological, chemical, mechanical, or morphological dormancy (Evans 1933, Baskin & Baskin 2004, 2008, Jacobo-Pereira et al. 2016, Gallegos-Mendoza et al. 2019). Seed dormancy occurs in temperate and tropical species of Magnolia (Baskin & Baskin 2014, Fernando et al. 2013) despite that the most tropical species are recalcitrant seeds with relatively short life span (Subbiah et al. 2019). In general, seeds of the genus Magnolia present low percentage of germination due to dormancy and various techniques have been used to break dormancy and increase germination (Vásquez-Morales & Sánchez-Velásquez 2011, Iralu & Upadhaya 2016, Vásquez-Morales & Ramírez-Marcial 2019).
Temperature fluctuation, including low temperature, is a requirement to activate the metabolic processes of germination and break seed dormancy in Magnolia seeds (Bewley & Black 1994, Baskin & Baskin 2004), since the slow release of food reserves from the endosperm to the embryo is probably in response to the natural temperature fluctuations that seeds experience in nature (Probert 2000, Baskin & Baskin 2014). The use of temperatures (4 to10 ºC) to break dormancy and subsequent germinative treatments increase the percent germination in Magnolia vovidesii, treated by Vovides & Iglesias 1996, and Corral-Aguirre & Sánchez-Velázquez (2006) as M. dealbata Zucc. (Vázquez-García et al. 2015, 2016a) and Magnolia schiedeana Schltdl. (Vásquez-Morales & Sánchez-Velásquez 2011).
A combination of environmental factors determines the timing of seed germination for some species. Light may regulate seed germination and early establishment of seedlings because phytochrome-mediated responses play a critical role in determining the time of germination (Baskin & Baskin 2014). Fluctuating temperatures can substitute light requirements for the germination of some positive photoblastic species (Pons 2000). Photoblastism is the seed’s response to different light conditions; 1) some seeds require light to germinate (positive photoblastism); 2) others require darkness to germinate (negative photoblastism); 3) others have a higher percentage of germination in light than in darkness; 4) others have a higher percentage in darkness than in light; and 5) some seeds germinate equally well in light and darkness (Baskin & Baskin 2014). Other authors have classified the types of photoblastism into three categories, where positive photoblastism is equivalent to category 1; negative photoblastism is equivalent to category 2; and the other three categories could be classified as neutral photoblastism or non-photoblstic (Fenner & Thompson 2005, Flores et al. 2016). It has been found that the percentage of germination of Magnolia kobus DC. and M. obovata Thunb. does not differ among red:far-red ratio under both constant and fluctuating temperatures, being an adaptive germination strategy to ensure seed germination in gaps, even for buried seeds in soil or litter (Xia et al. 2016). Seed conditions such as light exposure and soil depth, among others, can change the dynamic of a population (Bond et al. 1999).
The genus Magnolia comprises 36 species in Mexico, almost 10% of the known species of this genus worldwide (350 spp.) (Vázquez-García et al. 2016b). Despite the high species richness and endemism of this group in Mexico, little is known about its light quality requirements or the effect of temperature on the development of Magnolia embryos, which are overriding factors in the early establishment of tree seedlings in tropical humid forests (Whitmore 1996). Low temperatures treatment are often used to break seed dormancy, for instance in the deciduous Magnolia sect. Macrophylla: M. vovidesii (Corral-Aguirre & Sánchez-Velásquez 2006); several species of the evergreen sect. Magnolia: Magnolia iltisiana A.Vázquez (Saldaña-Acosta et al. 2001); M. schiedeana (Vásquez-Morales & Sánchez-Velásquez 2011), and Magnolia pugana (Iltis & A. Vázquez) A. Vázquez & Carvajal (Jacobo-Pereira et al. 2016); and two species of the evergreen sect. Michelia: Magnolia punduana Hook. f. & Thomson and M. champaca (L.) Baill. ex Pierre (Silvertown 1999, Fernando et al. 2013, Iralu & Upadhaya 2016).
Magnolia alejandrae García-Morales & Iamonico is a recently described and critically endangered species endemic to the northeast of Mexico, known only from its taxonomic description (García-Morales et al. 2017) and its interaction with the insect, Leptoglossus dilaticollis Guérin-Méneville (Gallardo-Yobal et al. 2020). The species has a narrow distribution in the Sierra Madre Oriental province (western center of the state of Tamaulipas), include five populations (www.naturalista.mx) in the tropical mountain cloud forest and the pine-oak forest between 1,500 and 1,600 m asl. At these sites, the mean annual temperature varies from 17 ºC during the day to 8 ºC at night (Fick & Hijmans 2017).
This work presents the photoblastic response of M. alejandrae seeds and the influence of temperatures on germination. The aim was to determine what effect light quality, temperature fluctuation and population provenance have on the germination of M. alejandrae. The hypothesis is that white light and fluctuating temperatures promote the germination of M. alejandrae, while M. alejandrae seeds from Los San Pedros will have a higher germination percentage than those from El Farallón, due to the first site being more humid than the second.
Materials and methods
Study species. Magnolia alejandrae is a deciduous tree, up to 16 m tall with grayish-brown bark and greenish patches (Figure 1A); obovate leaves with a silky texture, cordate leaf bases, acute to attenuate at the apex; open flowers 30-34 cm in diameter, petals oblong with attenuate base, creamy white, glabrous, with 5 or 7 well-marked stripes, with a purple spot close to their truncated bases, glutinous around the blotch (Figure 1B). Flowering occurs between April and May. Fruiting occurs from August to September.

Figure 1 Morphological characteristics of Magnolia alejandrae: bark (A), flower (B) and fruit (C). Photo credit: SIGY.
The seeds have a three-layer coat, the outer one is bright reddish and rugose-striate and is called sarcotesta (presumably a bird dispersal syndrome), fleshy and with persistent oily scents that make it impermeable to water and gas exchange, protective from some diseases and predators and containing germination inhibitors; the middle hardcover, called sclerotesta, is bony blackish and provides mechanical protection; the inner coat, a thin membrane, surrounds the endosperm, including a rudimentary embryo (Callaway 1994, Vásquez-Morales et al. 2010, Vázquez-García et al. 2021a) (Figure 1C).
Magnolia alejandrae is similar to all other eight members of section Macrophylla including the newly described M. mixteca A. Vázquez & Domínguez-Yescas and M. zotictla A. Sánch.-Gonz., Gut.-Lozano & A. Vázquez (Sánchez-González et al. 2021, Vázquez-García et al. 2021b) in having a deciduous habit, short longevity (ca. 75 years), giant flowers and large (abaxially glaucous leaves). Its morphologically closest species is M. nuevoleonensis A. Vázquez & Domínguez-Yescas (García-Morales et al. 2017) and recent molecular evidence from a multi-gen approach (Ibarra-Martínez 2020) and microsatellites (Chávez-Cortázar et al. 2021) support a close relationship between these two taxa, despite their results including a clear geographic structure, high genetic differentiation and fine-scale genetic structure supporting allopatric speciation for these species, the latter authors suggest that all five Mexican members of Magnolia sect. Macrophylla are conspecific. Here, we adopt the species concept used by de Queiroz (1998, 2007), also in agreement with García-Morales et al. (2017) in that Magnolia alejandrae differs morphologically from M. nuevoleonensis in having larger flowers, a brighter and much wider purple petal spot, and narrower fruits.
Seed collection sites. We collected seeds of M. alejandrae from two population provenances in the Sierra Madre Oriental in Tamaulipas. The first population site was El Farallón (23° 52' 03.90" N; 99° 23' 21.31" W) at 1,655 m asl, in the Altas Cumbres Natural Protected Area of the Jaumave municipality; the second is at Los San Pedros (23° 57' 26.63" N; 99° 28' 41.21" W) at 2,050 m asl, in the Güémez municipality. The climate at both sites is sub-humid temperate, with an average annual temperature of 17.4 °C and annual rainfall of 800-1,200 mm, in pine and oak forest (Fick & Hijmans 2017).
We collected seeds in May and June, 2019, when the fruits were ripe but before dehiscence to avoid losing the seeds. We collected three fruits, pest and disease-free, from each of 30 randomly distributed M. alejandrae trees in each one of the two sites, specially avoiding those frequently damaged by the leaf-footed bug, L. dilaticollis (Gallardo-Yobal et al. 2020). Distance between M. alejandrae trees was a minimum of 50 m and a maximum of 100 m in the two study sites. The fruits were placed in paper bags, labeled and transported to the laboratory for drying. They were placed on an aluminum table at room temperature to allow the follicles containing the seed to open, preventing them from being rendered unviable from excess heat, since Magnolia seeds are recalcitrant (Del Tredici 1981). Follicles dehisced within three days. Subsequently, the seeds were immersed in water for 24 hr to facilitate removal of the sarcotesta by wrapping with a plastic mesh until the sclerotesta was exposed.
Experiment design. We obtained a work sample from the batch of seeds from each study site using International Seed Testing Association (ISTA 2017) methodology. Twenty seeds were placed in each 9 cm Petri dish, with five repetitions for each light treatment (100 seeds × treatment), for two temperature treatments and two study sites, having a total of 1,600 seeds. All seeds were disinfected with 5 % bleach for 15 min and then washed and immersed in running water for 24 hr, before being removed and placed on layers of cotton moistened with distilled water under cold stratification (5 ºC) for seven days.
We performed a cold stratification as recommended by Evans et al. (1933) to break seed dormancy, and the temperature used in the stratification of M. alejandrae matched that of in situ provenance. Once we completed this process, we exposed the seeds from each population site to four light quality treatments: red, far-red, white and dark. For the red-light simulation, we covered the Petri dishes with red (Lee, number 071) filters; and for the far-red with overlapping red and blue (Lee, number 182) filters. We used the definition by Tsuboi et al. (2012) and González de León et al. (2021) to delimit the red light (650-670 nm) and far-red (730-750 nm), whose ratios were 7.22 and 0.12, respectively. We determined the light quality by spectroradiometer LI-180 (Lincoln, LI-COR, EUA). The seed dishes being subjected to darkness were covered with aluminum to prevent light from entering. Finally, for the white light treatment, we exposed the dishes to unfiltered direct light. We used water agar (Meyer©) at 5 % as substrate and sealed the Petri dishes with parafilm. We exposed the seeds of each population and each light treatment to two temperature levels: in chamber I (Luministell©, model IES-OS, México), the temperature was constant at 18 °C with a 12-hr photoperiod, while the temperature in chamber II (Luministell©, model ICP-O9, México) fluctuated between 8 ºC and 18 ºC with a 12-hr photoperiod. This temperature range represents the variations that occurs in the study sites (Fick & Hijmans 2017). Each chamber was equipped with white and cold light lamps of 32 W (Phillips©). Daily recorded germinated seeds under white light except those under red, far-red and darkness treatments, recorded towards the end of the experiment (44 days after seeding). A seed was considered to have germinated when the radicle was visible (Finch-Savage & Leubner-Metzger 2006).
We calculated final germination percentage as:
where n is the number of germinated seeds and N is the total number of seeds. We calculated the speed of seed germination under white light based on Kotowski’s coefficient of velocity:
where ni is the number of germinated seeds in the day i, and ti is the number of days after seeding. KCV takes values from 0 to 100; high values represent a high germination velocity (González-Zertuche & Orozco-Segovia 1996). We calculated the time to reach 50 % germination (t50) according to Grime et al. (1981).
Statistical analyses. We analyzed the data in R version 4.0.4 (R Core Team 2021). Effects of light quality (white, red, far-red, and dark), site (El Farallón and Los San Pedros) and temperature (18/18 °C and 8/18 °C) on germination probability were analyzed using generalized linear models (GLM) with a quasibinomial distribution to correct overdispersion and a logit link function (Crawley 2007) using “lme4” package (Bates et al. 2015). We corroborated the absence of multicollinearity between variables by the variance inflation factor (VIF) function of the “car” package (Fox & Weisberg 2019). We applied the analysis of variance (ANOVA) by Wald test to determine the significance of each factor (light quality, site and temperature) and their interaction on the germination (Dunn & Smyth 2018), and subsequently multiple means comparisons (Holm, P ˂ 0.05) to evaluate possible differences between pair light quality treatments. Holm's method is an adjustment to the p-value used in multiple hypothesis tests to prevent false statistical inferences (Holm 1979).
We applied a two-way analysis of variance (ANOVA) to test the effect of temperature (two levels) and population provenance (two levels) on germination velocity and germination at t50 of M. alejandrae seeds under the white light treatment. We checked normality using the Shapiro-Wilk test.
Results
The three-way ANOVA indicated that the percentage of germination did not differ significantly among temperature, light quality, and population site combinations (P > 0.05; Table 1). However, the light quality and temperature factors had a significant effect on the germination of M. alejandrae (P < 0.05; Table 1), where dark and far-red treatments showed a negative impact on germination, and the temperature of 8/18 °C had the highest positive effect (Table 2). The population provenances and the interaction of factors studied had no significant outcome on the germination of M. alejandrae (P > 0.05; Table 1 and 2). The light quality explained more variance (70.6 %) on seed germination, than temperature (1.7 %) (Table 3). M. alejandrae seeds from each population presented a similar germination pattern under different light quality (Figure 2), the highest germination occurring under white light (~70 %), followed by red light (45 %), far-red (9 %) and darkness (~5 %). Multi-comparison tests showed significant differences among all pairs of light quality treatments (P < 0.001, Table 4).
Table 1 Three-way analysis of variance to estimate the effect of temperature, light quality, population site and their interaction on percentage of germination of Magnolia alejandrae. The significance P value of the Wald test was < 0.05.
| Df | Chisq | Pr(> Chisq) | |
|---|---|---|---|
| (Intercept) | 1 | 0.070 | 0.790 |
| Light quality (Lq) | 3 | 139.042 | ˂ 0.001 |
| Population site (Ps) | 1 | 0.080 | 0.777 |
| Temperature (T) | 1 | 4.116 | 0.042 |
| Lq × Ps | 3 | 0.298 | 0.960 |
| Lq × T | 3 | 0.963 | 0.810 |
| Ps × T | 1 | 0.001 | 0.970 |
| Lq × Ps × T | 3 | 2.612 | 0.455 |
Table 2 GLM model with quasibinomial distribution on germination of Magnolia alejandrae under different light quality, population site, and temperature regimes. The white light, El Farallón and 18/18 °C are not listed as coefficients because they were used as reference points for the model. The significance P value was < 0.05.
| Estimate | t value | Pr(> |t|) | |
|---|---|---|---|
| Intercept | 0.031 | 0.265 | 0.791 |
| Dark | -2.508 | -9.825 | ˂ 0.001 |
| Red light | -0.523 | -3.01 | ˂ 0.001 |
| Far-red light | -1.756 | -8.498 | ˂ 0.001 |
| Los San Pedros | 0.048 | 0.283 | 0.778 |
| 8/18 ºC | 0.348 | 2.029 | 0.046 |
| Dark × Los San Pedros | 0.096 | 0.274 | 0.784 |
| Red light × Los San Pedros | -0.022 | -0.093 | 0.925 |
| Far-red light × Los San Pedros | 0.114 | 0.398 | 0.692 |
| Dark × 8/18 ºC | -0.348 | -0.964 | 0.338 |
| Red light × 8/18 ºC | -0.107 | -0.44 | 0.661 |
| Far-red light × 8/18 ºC | -0.126 | -0.442 | 0.659 |
| Los San Pedros × 8/18 ºC | 0.009 | 0.037 | 0.97 |
| Dark × Los San Pedros × 8/18 ºC | 0.597 | 1.236 | 0.22 |
| Red light × Los San Pedros × 8/18 ºC | 0.127 | 0.37 | 0.712 |
| Far-red light × Los San Pedros × 8/18 ºC | -0.231 | -0.573 | 0.568 |
Table 3 Percentage of contribution of each variable on the germination of Magnolia alejandrae.
| DF | Sum Sq | Percentage of contribution | F values | Pr(> F) | |
|---|---|---|---|---|---|
| Light quality (Lq) | 3 | 62.845 | 70.63 | 57.793 | ˂0.001 |
| Population site (Ps) | 1 | 0.029 | 0.03 | 0.080 | 0.778 |
| Temperature (T) | 1 | 1.498 | 1.68 | 4.132 | 0.046 |
| Lq × Ps | 3 | 0.108 | 0.12 | 0.099 | 0.959 |
| Lq × T | 3 | 0.349 | 0.39 | 0.321 | 0.810 |
| Ps × T | 1 | 0.001 | 0 | 0.001 | 0.970 |
| L × Ps × T | 3 | 0.95 | 1.07 | 0.873 | 0.459 |
| Residuals | 64 | 23.198 | 26.07 |
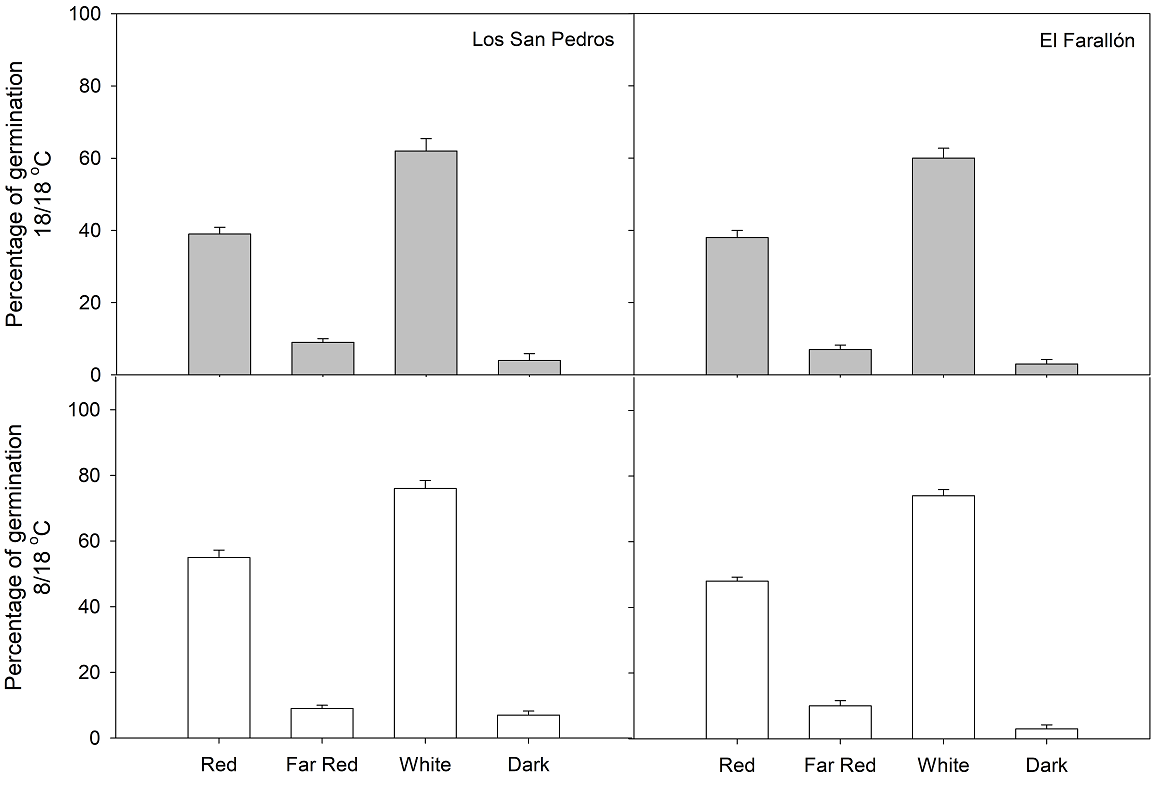
Figure 2 Germination percentage of Magnolia alejandrae under red, far-red, white and dark light. The seeds come from two populations: El Farallón and Los San Pedros, Tamaulipas.
Table 4 Paired comparisons of light quality on germination of Magnolia alejandrae. The significance P value was < 0.001.
| Contrast | Estimate | z value | Pr(> |z|) | ||
|---|---|---|---|---|---|
| White | - | Dark | 2.4849 | 20.558 | ˂ 0.001 |
| White | - | Red | 0.5535 | 6.413 | ˂ 0.001 |
| White | - | Far red | 1.8207 | 18.07 | ˂ 0.001 |
| Dark | - | Red | -1.9314 | -15.956 | ˂ 0.001 |
| Dark | - | Far red | -0.6643 | -5.042 | ˂ 0.001 |
| Red | - | Far red | 1.2671 | 12.551 | ˂ 0.001 |
The germination velocity of the seeds of M. alejandrae and the number of days to 50 % of germination (t50) were influenced by the interaction of the site and temperature (F (1, 16) = 22.7, P < 0.05; F (1, 16) = 19.6, P < 0.05, respectively; Table 5).
Table 5 Germination velocity and germination at t50 of Magnolia alejandrae under white light and constant temperature (18/18 ºC) and fluctuating temperature (8/18 ºC). Different letters mean significant differences between temperature treatments and population sites for each variable analyzed, P < 0.05.
| 18/18 ºC | 8/18 ºC | |
|---|---|---|
| Germination speed (%) | ||
| El Farallón | 2.39 ± 0.01 b | 2.40 ± 0.01 b |
| Los San Pedros | 2.36 ± 0.01 c | 2.42 ± 0.01 a |
| t50 (days) | ||
| El Farallón | 37.67 ± 0.15 b | 37.60 ± 0.23 b |
| Los San Pedros | 39.26 ± 0.46 a | 36.72 ± 0.13 b |
Germination was epigeal, and sclerotesta rupture occurred on day 31st; the seedlings remained alive for 44 days after seeding and presented high mortality at 50 days. A lapse of 10 days occurred between the emergence of the first radicles to the formation of the seedling, during which the first leaves, anchor roots and secondary roots appeared, as well as the presence of photosynthetic tissue (Figure 3).
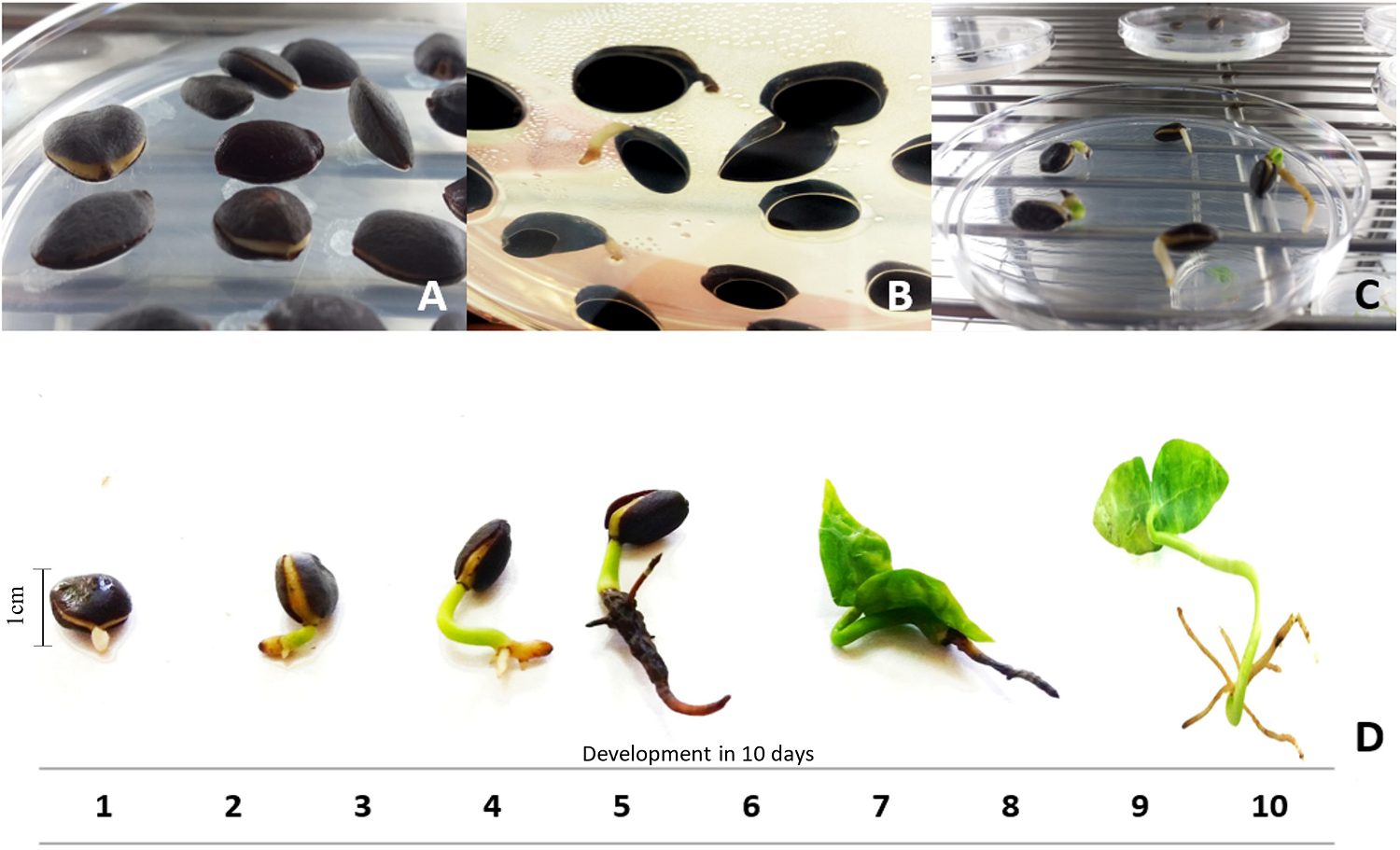
Figure 3 Development of Magnolia alejandrae seedlings in a period of 10 days under white light: A) Rupture of the sclerotesta (31 days after sowing), B) Emergence of the first radicles (epigeal germination), C) Growth of radicle and formation of the first secondary roots (shows photosynthetic tissue), and D) Seedling 10 days old, from germination (44 days).
Discussion
Light quality and temperature had a significant effect on the germination of M. alejandrae in agreement with our hypothesis. Seeds germinated more under white light (76 %) than in darkness (˂ 5 %), hence this species can be classified in category 3 (seeds that have higher percentage of germination in light than in darkness) according to Baskin & Baskin (2014). Moreover, M. alejandrae seeds can be considered neutral photoblastic or non-photoblastic, in which seed germinate both in the dark and under white light (Fenner & Thompson 2005, Flores et al. 2016). The neutral photoblastism of M. alejandrae coincides with Magnolia officinalis Rehder & E.H.Wilson (Zhou et al. 2012), and the species M. kobus and M. obovate, which are indifferent to light quality (Xia et al. 2016).
The high percentage of germination of M. alejandrae seeds under white light suggests that this species could take advantage of the forest gaps to germinate, indicating that this species responds as a pioneer species (Fenner & Thompson 2005). Moreover, pioneer seed species are stimulated by an increased range of temperature fluctuation that occurs in large canopy gaps (Pearson et al. 2002). In contrast, the low germination in the darkness of M. alejandrae seeds is probably a reflection of its adaptive capacity to ensure seed germination under forest litter, where other tree species such as Liquidambar styraciflua L. and Pinus sp. protect the Magnolia spp. seedlings from exposure to direct sunlight, avoiding desiccation (Ramírez-Bamonde et al. 2005, Arrieta & Suárez 2006).
In the field, light received by seeds has a direct effect on temperature and water availability that affect germination. Fluctuation temperature (8/18 ºC) enhanced germination of M. alejandrae seeds from the two studied populations (Table 2) and increase percent germination in tropical species (Pearson et al. 2002), including Magnolia species (Xia et al. 2016); this suggests that fluctuating temperatures can stimulate germination of tropical pioneer species in large-canopy gaps, even when seeds buried beneath the soil litter (Pearson et al. 2002).
The use of low temperatures (4 to -1 ºC) during seed stratification and subsequent exposure to constant temperatures from 20 to 28 ºC promotes germination in some species, such as Magnolia punduana and M. champaca belonging to the Magnolia sect. Michelia, a group consisting of species confined to tropical Asia (Silvertown 1999, Fernando et al. 2013, Iralu & Upadhaya 2016). Nevertheless, in our study, the prior stratification and the subsequent exposure of the seeds to temperature fluctuation increased the percent germination of M. alejandrae. Exposure to cold and warm periods is a process that occurs naturally in forest species. Species such as the Fabaceae, inhabiting pine and oak forests, are found in environments susceptible to frost and generally present physiological dormancy (Jurado & Flores 2005). Temperature is a primary factor to stimulate germination and regulate changes in the state of dormancy (Vleeshouwerset al. 1995,Benech-Arnoldet al. 2000). The low temperatures in the high mountains of the Sierra Madre Oriental could facilitate the germination of M. alejandrae seeds during fall, but field experiments are needed to prove this hypothesis. This low temperature could also facilitate assisted migration to immediately higher elevations as shown in Magnolia vovidesii (also a member of sect. Macrophylla), having an increase in relative growth rates and survival for saplings, and could become a promising conservation strategy for cloud forest trees to mitigate the impacts of climate change (García-Hernández & Toledo-Aceves 2020).
Several works on Magnolia germination have used treatments to break dormancy, including imbibition, cold moist stratification, acid (chemical) and mechanical scarification, the use of hydrogen peroxide and even hormones such as gibberellic acid (GA3) (Saldaña-Acosta et al. 2001, Corral-Aguirre & Sánchez-Velásquez 2006, Vásquez-Morales & Sánchez-Velásquez 2011, Iralu & Upadhaya 2016, Jacobo-Pereira et al. 2016, Vásquez-Morales & Ramírez-Marcial 2019). However, this study shows that M. alejandrae seeds exposed to fluctuating temperatures (8/18 ºC) enhanced the percentage of germination in comparison to 18/18 ºC.
Evans (1933) used fluctuating temperatures to germinate Magnolia grandiflora L., obtaining 10 % of seeds germination below 29 °C, while 40 % germination under alternating temperatures of 10/23 ºC. The international rules for seed testing (AOSA 2017), recommend 45 days of pre-chilling followed by alternating temperatures of 20/30 °C for M. grandiflora. For other species, such as M. officinalis, with alternated temperatures of 25/30 ºC, achieve 59 % germination, while temperatures of 20/30 ºC achieved 65 % germination (Xiao 2010). These results are similar to those reported for M. alejandrae under constant temperature (18/18 °C) that reached 62 % germination.
The germination of M. alejandrae seeds began at 31 days after seeding, reaching the seedling stage at 44 days (Figure 3). In other species of the genus Magnolia, variations have been found in the number of days to onset germination. For example, the seeds of M. grandiflora germinate after 60 days of storage in vermiculite (Hanchey & Kimbrough 1954); Magnolia perezfarrerae A. Vázquez & Gómez-Domínguez in 75 days; Magnolia sharpii Miranda in 45 days (Vásquez-Morales & Ramírez-Marcial 2019) and M. punduana in 212 days (Iralu & Upadhaya 2016). Magnolia alejandrae germinates in fewer days than the previously mentioned Magnolia species, may be as a mechanism to reduce desiccation mortality (Yirdaw & Leinonen 2002).
Rapid germination is a good indicator to assess the speed of occupation of a species in a given environment (Ferreira et al. 2001). In this respect, the germination at t50 of M. alejandrae was influenced by temperature and site interaction, having significant differences between population provenance under constant temperature treatment, with an average of 38.4 days. In the case of M. vovidesii, it takes 39.4 days to germinate 50 % of the seeds (Toledo-Aceves 2017), matching that of M. alejandrae in the Los San Pedros population under constant temperature but is higher than the seeds from El Farallón in the two temperature treatments (Table 5). Similarly, species from the tropical montane cloud forest, such as Carpinus caroliniana Walt., require 36.5 days to germinate 50 % of the seeds after being stored for two months, matching the results for M. alejandrae under fluctuating temperatures. Other species, like L. styraciflua, require only 5.5 days to germinate 50 % of the seeds (Linera & Pérez 2005), reflecting the capacity of the species from the tropical montane cloud forest to use environmental resources differentially.
In summary, we demonstrate that 1) Magnolia alejandrae seeds are neutral photoblastic and 2) light quality and fluctuating temperatures have a direct effect on germination; this is relevant in understanding how environmental factors, such as temperature and light, influence the initial stage of seed development. The environmental factors required for germination vary from one species to another, hence, being relevant to recognize the abiotic and biotic factors necessary for seed germination to determine the type of dormancy a species presents; likewise, microclimatic requirements are fundamental for the seeds to become established since these directly influence the obtaining of seedlings for population recovery. However, further ecophysiological studies of Magnolia alejandrae seeds are needed, such as the embryo/endosperm relationship, rupture of the sclerotesta, successful in situ germination, establishment, and survival.

