











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Las plantas carnívoras pertenecen a un grupo polifilético diverso de plantas con flores que generalmente se encuentran restringidas a suelos húmedos y pobres en nutrientes, sus 20 géneros representan al menos 11 linajes evolutivamente diferentes que coinciden en su mayoría de un grupo de características funcionales (Ellison & Adamec 2018, Adamec et al. 2021, Lin et al. 2021) que les permiten obtener la mayoría de sus nutrientes de presas animales como arañas, crustáceos, insectos, protozoos, reptiles, roedores (Frank & O'Meara 1984, Givnish & Sytsma 1997, Ellison & Gotelli 2009, Pereira et al. 2012, Lin et al. 2021) y que incluyen un movimiento rápido de trampas reguladas electrofisiológicamente, secreción de enzimas hidrolíticas capaces de digerir quitina, proteínas y otros compuestos orgánicos, así como la absorción foliar de nutrientes, lo cual también estimula la absorción de éstos a través de las raíces así como el crecimiento (Adamec 2011, Ellison & Adamec 2018, Adamec et al. 2021). Se ha documentado la participación de metabolitos secundarios en la atracción, captura, digestión y asimilación de presas (Hatcher et al. 2020). La mayoría de las plantas con este tipo de nutrición pueden crecer sin consumir presas, pero con los nutrientes derivados de ellas crecen más rápido y se reproducen mejor al utilizar animales como fuente de nitrógeno, fósforo y potasio, las plantas carnívoras forman parte del ciclo de algunos elementos químicos y regulan ciertas poblaciones de organismos (De Ridder & Dhondt 1992, Adamec 1997, Thorén & Karlsson 1998).
La carnivoría como adaptación, parece estar limitada a los cambios en los factores ambientales, por ejemplo, la humedad, radiación fotosintéticamente activa y contenido de nitrógeno en el suelo, así como la disponibilidad de presas tiene efectos en la variación fenotípica de poblaciones. Por ejemplo, la adherencia de las hojas de Pinguicula vallisneriifolia, esta aumenta bajo mayores niveles de exposición solar (Zamora et al. 1998), en Sarracenia purpurea si la disponibilidad del nitrógeno en el suelo asciende, provocará una disminución en la producción de jarras carnívoras y estimulará el desarrollo de filodios (hojas especializadas en realizar fotosíntesis) (Ellison & Gotelli 2002), en Drosophyllum lusitanicum la adición de presas en su alimentación incrementa la biomasa (peso seco) respecto a plantas no alimentadas (Paniw et al. 2017).
Específicamente en Pinguicula moranensis se ha observado que la baja disposición de nitrógeno del suelo, está relacionado con la presencia de plantas con mayor número de glándulas adhesivas, pero cuando hay mayor disposición de este nutrientes, este se asocia a plantas con mayor número de glándulas digestivas, estas observaciones dan indicio de que el ambiente puede promover la expresión y prevalencia de fenotipos mejor adaptados a condiciones particulares (Alcalá & Domínguez 2003, 2005), pero es posible que no sea el único factor asociado al fenotipo.
En México se han reportado dos familias de plantas carnívoras: Droseraceae (con registro de dos especies: D. brevifolia y D. capillaris) y Lentibulariaceae (con registros de sus géneros: Genlisea, Pinguicula y Utricularia) (Olvera-García & Martínez 2002, Basso 2009, Villaseñor 2016). Para el estado de Tlaxcala se han reportado tres especies de plantas carnívoras: Pinguicula acuminata, P. moranensis y Utricularia livida (Olvera 1996, Zamudio-Ruíz 2005, Villaseñor 2016). P. moranensis var. neovolcanica Zamudio, se encuentra restringida a la Faja Neovolcánica Transmexicana (Zamudio 1999). Particularmente, en el estado de Tlaxcala se ha reportado en 14 municipios (Calpulalpan, Chiautempan, Españita, Hueyotlipan, Ixtacuixtla, Nativitas, Nanacamilpa, Panotla, San José Teacalco, Teolocholco, Terrenate, Tlaxcala, Tlaxco y Totolac) (Datos de herbarios MEXU y TLXM, Material Suplementario, Tabla S1). Ante la inminente afectación del hábitat de esta especie y la ausencia de estudios que consideren las características ambientales y poblacionales con fines de conservación, el propósito de este estudio fue determinar el efecto de los factores ambientales, en los caracteres morfológicos, poblacionales y reproductivos de P. moranensis en bosques de Quercus-Pinus en Tlaxcala.
Materiales y métodos
Especie de estudio. Pinguicula moranensis es una planta herbácea perenne, presenta un dimorfismo estacional el cual está definido por la presencia de una roseta de invierno, compacta en forma de bulbo e hipogea, es morfológicamente distinta a la roseta de verano en la que las hojas se distinguen y aparece la floración. Crece sobre suelos o rocas de origen ígneo, bajo laderas con sombra con pendientes muy inclinadas, principalmente en bosques de Quercus, Pinus-Quercus y Abies, dentro de un gradiente altitudinal de 1,700 a 3,300 m de altitud en el Eje Volcánico Transmexicano (Zamudio 1999).
Sitios de estudio. En julio-septiembre de 2018 se realizaron visitas a los 14 sitios de registro de la especie P. moranensis, la presencia de la especie (y su eventual ausencia) determinó la elección de los sitios de estudio que se describen a continuación. Cerro Popocatl ubicado en la localidad de San Ambrosio Texantla, municipio de Panotla, Tlaxcala (referida posteriormente como Panotla) a 2,378 m de altitud. Colinda al norte con los municipios de Xaltocan y Hueyotlipan; al sur con los municipios de Nopalucan y Texoloc, al oriente se establecen linderos con los municipios de Totolac y Tlaxcala, asimismo al poniente colinda con el municipio de Ixtacuixtla (Figura 2). La vegetación de la parte con mayor altitud del municipio está representada por Pinus pseudostrobus, Quercus spp. y Cupressus lusitanica var. benthamii (INAFED 2010).
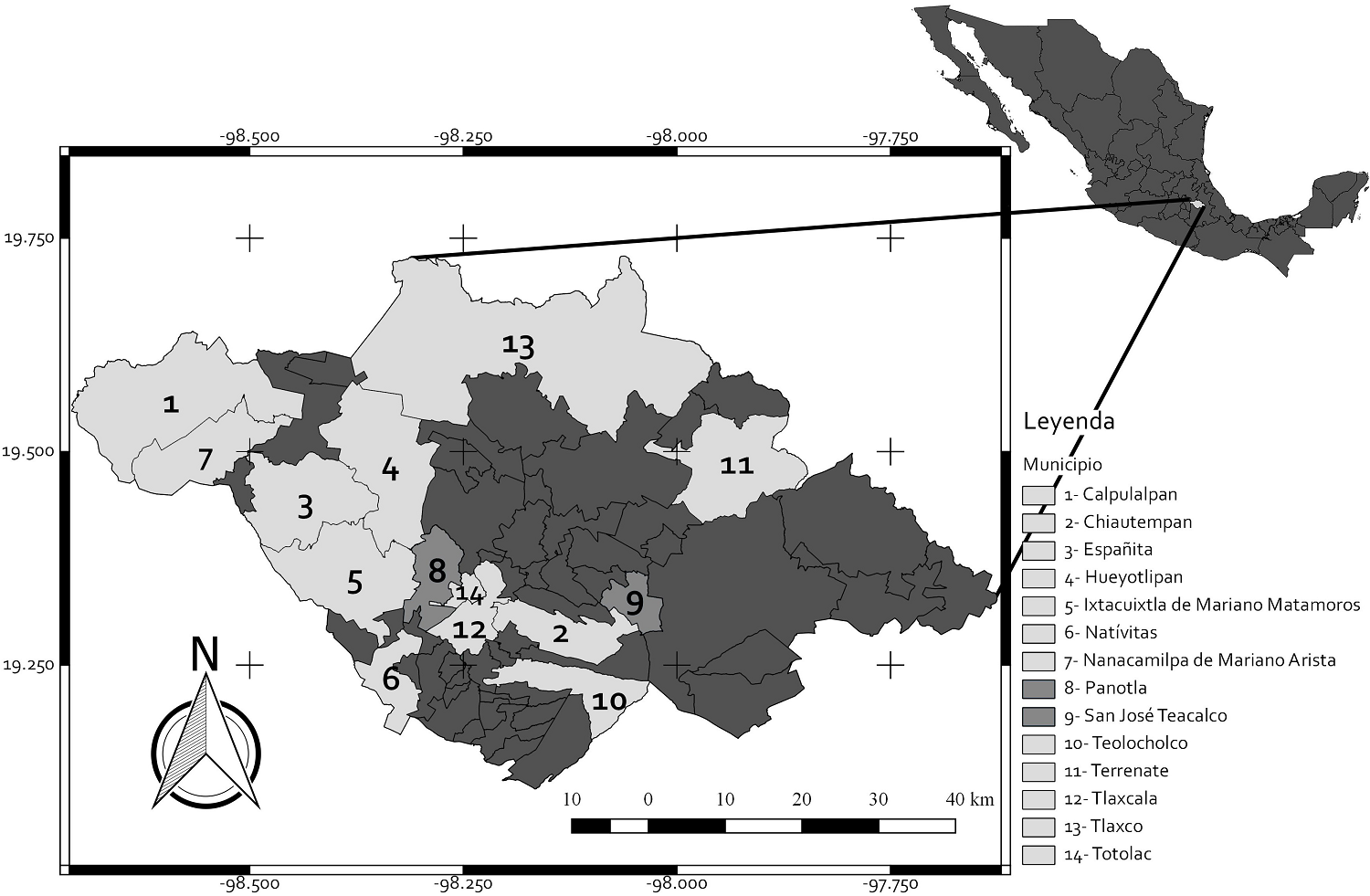
Figura 1 Municipios de Tlaxcala en los que se ha registrado P. moranensis (actualmente sin presencia de la especie) los municipios 8 y 9 son los puntos en donde se confirmó la presencia de poblaciones.
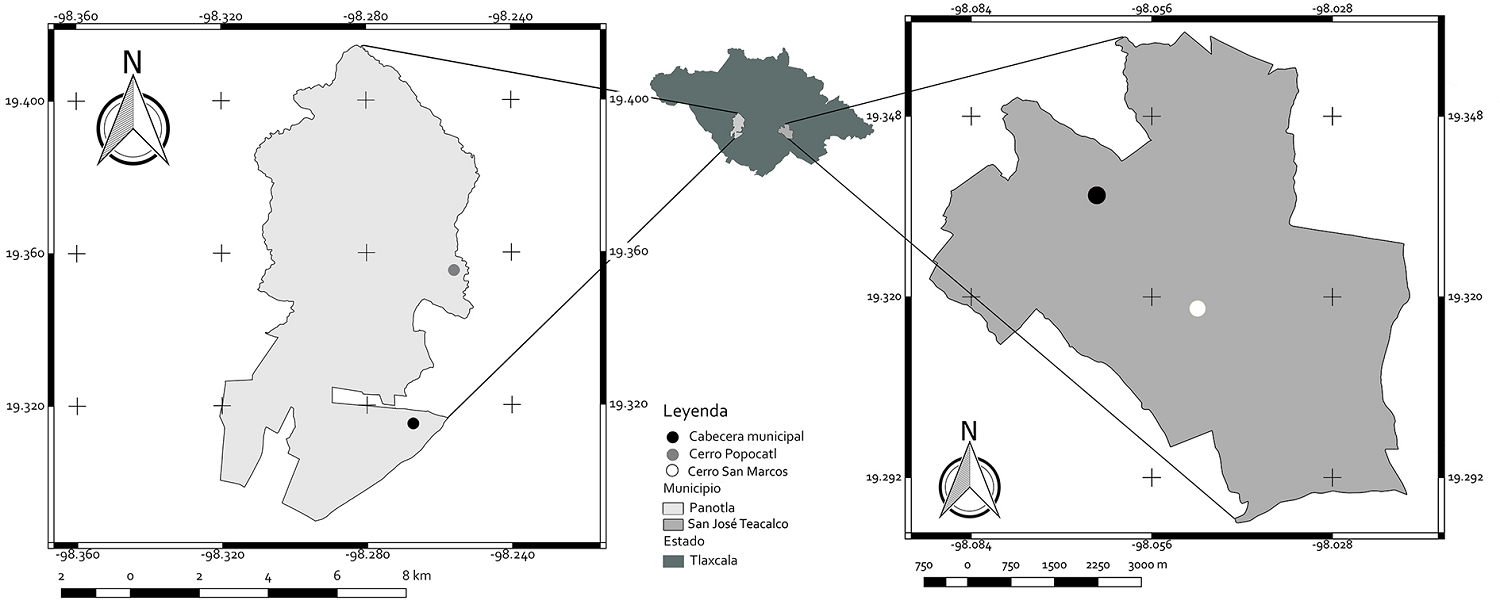
Figura 2 Ubicación de los sitios en que se localizaron las poblaciones de P. moranensis, a la izquierda el Cerro Popocatl en Panotla, Tlaxcala, a la derecha el Cerro San Marcos en San José Teacalco, la marca de la cabecera municipal indica la proximidad del área urbanizada con los sitios de estudio.
Al oriente del estado: Cerro San Marcos San José Teacalco (referida posteriormente como Teacalco) a 2,750 m de altitud. Colinda al norte con los municipios de Cuaxomulco y Tzompantepec, al sur y al oriente con el municipio de Huamantla y al poniente con los municipios de Contla de Juan Cuamatzi, Chiautempan y San Francisco Tetlanohcan. Se ubica dentro del polígono del volcán de La Malinche, la vegetación es de Abies religiosa, Alnus jorullensis, Arbutus xalapensis, Pinus moctezumae, P. pseudostrobus, P. tecote, Quercus laurina y Q. crassipes (Fernández & López-Domínguez 2005, INAFED 2010) (Figura 2).
En cada sitio de estudio se identificaron espacialmente parches con individuos de P. moranensis, los cuales consistieron en agrupaciones de plantas con una distancia entre ellos < 0.5 m, (14 en Teacalco y seis en Panotla). Se seleccionaron 50 plantas de P. moranensis, los individuos seleccionados fueron señalizados colocando en el suelo un abatelenguas de madera. Para efectos del estudio a una escala de distribución por estado se consideran los individuos de los dos sitios pertenecientes a una misma población.
Ambiente. Variables abióticas.- En julio de 2019 en ambos sitios, las lecturas de la intensidad de luz y humedad se tomaron en una única ocasión, se registró la intensidad de luz (iluminancia), expresada en unidades lux, con un Medidor digital HS1010-HS1010A LCD (Sunche, EUA). El sensor se colocó por encima de la roseta de cada planta marcada. Para medir la humedad ambiental (porcentaje) se utilizó un termómetro-higrómetro digital Taylor® 1732 (Taylor®, Illinois, EUA), este se colocó al nivel del suelo durante dos minutos.
Variables bióticas.- Se estimó la densidad de individuos, (el cociente del número de plantas dividido entre el área del parche en m2). Para determinar las especies de artrópodos que son presas de P. moranensis en los dos sitios de estudio, en julio de 2019 se realizó la colecta de presas con pinzas entomológicas en sólo una hoja de cada una de las plantas marcadas (Figura 3B). En el caso de presas visualmente indistinguibles o frágiles, se colectaron con la hoja completa, los artrópodos y/o las hojas con artrópodos se almacenaron en tubos Eppendorf de 3 ml con alcohol al 70 %, la identificación taxonómica de las presas se realizó con observaciones de los rasgos morfológicos en un microscopio estereoscópico y utilizando las claves dicotómicas de Triplehorn & Johnson (2005). Se cuantificó el número de individuos de cada orden taxonómico que fueron capturados como presas en cada hoja asumiendo un tiempo de captura de un día antes de ser digeridos y con estructuras identificables (Heslop-Harrison & Knox 1971, Primer et al. 2018), con el mismo nivel se realizó la estimación de la diversidad taxonómica promedio (average taxonomic distinctness, Δ+) Así, a mayor valor del índice, las presas capturadas pertenecerán a diferentes órdenes y clases taxonómicas de artrópodos (García-de Jesús et al. 2016). Este índice se calculó en el programa Past 4.05 (Hammer et al. 2001).

Figura 3 A: individuos de P. moranensis monitoreados, B: Presas de P. moranensis en el Parque Nacional La Malinche, C: colores de las flores-trampa, D: flores trampa, E: flores trampa en el área de muestreo, F: Semilla de P. moranensis a 6.3 x, G: embrión a 6.3 x, H-J individuos de P. moranensis con 3 (H a 6.3 x), 6 (I a 3.6 x), 12 (J a 1.7 x), 18 (K a 1.5 x) y 30 (L) días bajo condiciones in vitro.
Para determinar taxonómicamente a los visitadores florales se elaboraron flores-trampa que visualmente se percibieron como réplicas de las flores de P. moranensis, las corolas fueron de láminas de espuma sintética (foami), con colores y tonos semejantes a las que se presentan en las poblaciones naturales, los cuales se identificaron en la PANTONE MATCHING SYSTEM® (Figura 3C, D, E). Las corolas artificiales estuvieron adheridas a un frasco receptor de 8 mm de diámetro y 35 mm de longitud que contenía shampoo para cabello (Caprice®) sin diluir. El frasco se fijó a una estaca de madera de 20 cm de longitud. Se colocaron 84 flores-trampa, en cada población, distribuidas entre los individuos monitoreados con una separación entre las trampas de 20 cm y se dejaron durante siete días en ambos sitios, posteriormente el contenido de todos los frascos se recolectó en recipientes de polipropileno con alcohol al 70 % y se transportaron al laboratorio para la identificación taxonómicas de los visitadores siguiendo el método descrito para la identificación de las presas, posterior a la identificación se determinó su abundancia por población.
Planta. Caracteres morfológicos.- Se determinó el diámetro de la roseta, la longitud de la flor y del pedúnculo (cm) de las plantas marcadas con un vernier digital 784EC (Sona Enterprises, China). La longitud de la flor se consideró como la longitud entre los lóbulos superiores y el espolón (Zamudio 1999). Se cuantificó el número de flores abiertas e independientemente el número de pedúnculos que podían contener botones florales cerrados, flores abiertas o frutos.
Caracteres reproductivos.- Para cuantificar las semillas, se recolectaron cápsulas maduras en ambos sitios de estudio, se almacenaron en frascos de cristal de 1 cm3. Para reducir la humedad de las semillas e inhibir el crecimiento de hongos, los frascos se destaparon y las cápsulas secaron a temperatura ambiente en condiciones de oscuridad por diez días. Posteriormente, se extrajeron las semillas, se colocaron en una caja Petri y se contaron en un microscopio estereoscópico. El tamaño de las semillas (utilizado para discriminar las semillas fértiles de las abortadas) se estimó con ayuda del software tpsDig2 de imágenes digitales. Se registró el número de semillas totales y abortadas por cápsula, se consideró una semilla abortada cuando la longitud fue ≤ 200 µm un tamaño mayor indicó que se trataba de semillas fértiles (Figura 3F, G) (Villegas & Alcalá 2018). Después del conteo, el contenido de semillas de cada cápsula se almacenó en frascos de cristal estériles de 4 cm3 en refrigeración a 4 °C hasta sembrarlas en cultivo in vitro.
La viabilidad de las semillas se evaluó a partir del porcentaje de germinación en cultivo in vitro. Previamente, las semillas se desinfectaron con una solución de hipoclorito de sodio al 30 % (fórmula comercial Cloralex®) por 30 segundos, posteriormente se agregó un mililitro de agua destilada y una gota de Tween 20 para romper la tensión superficial y manualmente se agitó la suspensión por 10 minutos, las semillas se sumergieron en solución de hipoclorito de sodio al 6 % por cinco minutos y por último, se colocaron un minuto en una solución antibiótica-antimicótica al 0.5 % (Sigma™), entre cada procedimiento las semillas se enjuagaron por triplicado con agua destilada estéril. Para la germinación se utilizaron cajas Petri de 100 × 20 mm adicionadas con 30 mL de medio MS (Murashige & Skoog 1962), con 30 g/L de sacarosa, 3 mg/L de ácido giberélico y 5.5 g/L de Phytagel ™ como agente gelificante. El pH del medio se ajustó a 5.75. En condiciones de asepsia, se sembraron 500 semillas por población, cultivando 50 semillas por caja. Se incubaron en una cámara de crecimiento Forma Scientific modelo 3740 (Forma Scientific, USA) durante siete días en condiciones de oscuridad, a una temperatura de 25 ± 2 °C. Posteriormente se mantuvieron a un fotoperiodo de 16 h luz y 8 h oscuridad con la misma temperatura. Se realizaron observaciones cada tres días durante 30 días para registrar el porcentaje de germinación de semillas por caja Petri (Figura 3H, I, J, K, L).
La viabilidad de las semillas se calculó con la siguiente fórmula:
Donde:
a |
= Total de semillas por cápsula. |
b |
= Total de semillas abortadas por cápsula. |
c |
= Porcentaje de semillas germinadas bajo cultivo in vitro. |
Análisis estadísticos. Para determinar el efecto del ambiente y de los caracteres morfológicos poblacionales sobre factores bióticos y caracteres reproductivos de P. moranensis, se utilizaron modelos lineales generalizados (GLM) (McCullagh & Nelder 1989). Se realizaron tres modelos, dos para evaluar el efecto de las variables de humedad (%), luz (lux), longitud de flores (cm), número de pedúnculos, densidad de individuos, número de semillas fértiles y viabilidad de semillas (%) sobre la abundancia y diversidad taxonómica de presas. El tercer modelo evaluó el efecto de las variables humedad (%), luz (lux), longitud de flores (cm), número de pedúnculos, densidad de individuos y viabilidad de semillas (%) sobre el número de semillas fértiles. Los dos primeros 9GLM fueron realizados con distribución Poisson (link = "logit"). El tercer modelo se realizó con distribución binomial en R (R Core Team 2016).
Resultados
Los datos obtenidos de las mediciones de las variables abióticas, variables bióticas, caracteres morfológicos y caracteres reproductivos se presentan en la Tabla 1. La determinación taxonómica de presas de P. moranensis en Tlaxcala mostró una mayor proporción de captura de individuos de los órdenes collembola, díptera e hymenoptera (Tabla 2), así mismo para los órdenes que se registraron como visitadores florales, en el caso de hymenoptera fueron diferentes que los que se registraron como presas y fue posible determinar las familias, sin embargo, en el caso de los dípteros visitadores florales fueron los mismos que los que se determinaron como presas en los cuales también se determinaron las familias (Tabla 3).
Tabla 1 Variables y caracteres de P. moranensis registrados en dos sitios de Tlaxcala (se presenta promedio ± desviación estándar).
| Variable o caracteres | Teacalco | Panotla | |
|---|---|---|---|
| Abióticas | Luz (lx) | 5,109 ± 4,750.84 | 14,679.600 ± 21,953.09 |
| Humedad (%) | 71 ± 7.27 | 48.300 ± 13.04 | |
| Bióticas | Densidad | 7.660 ± 4.41 | 9.373 ± 5.20 |
| Presas por planta | 7.200 ± 5.47 | 5.300 ± 3.92 | |
| Morfológicos | Diámetro de roseta (cm) | 6.817 ± 1.95 | 7.914 ± 1.22 |
| Longitud de flores (cm) | 2.565 ± 0.66 | 2.751 ± 0.45 | |
| Longitud pedúnculos (cm) | 9.533 ± 3.99 | 12.552 ± 1.90 | |
| Número de flores | 1.100 ± 0.32 | 1.100 ± 0.32 | |
| Número pedúnculos | 1.700 ± 0.68 | 1.600 ± 0.70 | |
| Reproductivos | Semillas abortadas | 122.800 ± 92.22 | 114.300 ± 63.61 |
| Semillas fértiles | 608.900 ± 188.14 | 387.100 ± 70.02 | |
| Semillas totales | 731.700 ± 170.48 | 501.400 ± 61.89 | |
| Viabilidad de semillas (%) | 54.140 ± 9.01 | 38.271 ± 5.79 |
Tabla 2 Composición taxonómica de presas de P. moranensis en los sitios de muestreo Panotla y Teacalco, se reporta el número de individuos capturados y el porcentaje que representaron en cada sitio
| Taxón de las presas | Panotla (%) |
Teacalco (%) |
Total (%) |
|---|---|---|---|
| Collembola | 46 (36.51) | 80 (58.82) | 126 (48.09) |
| Diptera | 61 (48.41) | 25 (18.38) | 86 (32.82) |
| Hymenoptera | 5 (3.97) | 16 (11.76) | 21 (8.02) |
| Acari | 10 (7.94) | 9 (6.62) | 19 (7.25) |
| Araneae | 1 (0.79) | 4 (2.94) | 5 (1.91) |
| Coleoptera | 2 (1.59) | 1 (0.74) | 3 (1.15) |
| Dermaptera | 0 | 1 (0.74) | 1 (0.38) |
| Hemiptera | 1(0.79) | 0 | 1 (0.38) |
Tabla 3 Composición taxonómica de visitadores florales de P. moranensis en los sitios de muestreo Panotla y Teacalco. Se presentan principalmente órdenes y familias pero en algunos casos se determinó el género o género y especie.
| Orden | Familia | Panotla (%) | Teacalco (%) |
|---|---|---|---|
| Hymenoptera: | Halictidae (Sphecodes sp.) | 4 (23.5) | 80 (88.9) |
| Chironomidae | 4 (23.5) | ||
| Formicidae | 2 (11.8) | 3 (3.3) | |
| Diptera: | Phoridae | 4 (23.5) | |
| Sciaridae | 3 (17.6) | ||
| Ceratopogonidae | 2 (2.2) | ||
| Curtonidae | 2 (2.2) | ||
| Lepidoptera: | Hesperiidae (Autochton cellus) | 2 (2.2) | |
| Coleoptera: | Melolonthidae (Euphoria basalis) | 1 (1.1) |
Los modelos lineares generalizados permitieron discernir que la variación en la abundancia de presas capturadas en las hojas de P. moranensis estuvo influenciada significativamente por la humedad, la longitud de las flores, el número de pedúnculos, la densidad, el número de semillas fértiles y la viabilidad de las semillas, pero no por la luz (Tabla 4). La diversidad taxonómica de presas capturadas en las hojas de P. moranensis estuvo influenciada de forma significativa y positiva por el número de semillas fértiles y de manera negativa por el porcentaje de humedad. El resto de las variables no mostraron influencia significativa (Tabla 5). Con relación al número de semillas fértiles hay un efecto significativo y positivo de la humedad, de la cantidad de luz, de la densidad de individuos y la viabilidad de semillas. Por el contrario, la longitud de las flores tiene un efecto significativo, pero de forma negativa. El número de pedúnculos no presentó relación significativa (Tabla 6).
Tabla 4 Efecto de las variables abióticas, bióticas, morfológicas y reproductivas en la abundancia de presas en las hojas de P. moranensis, a través de un modelo lineal generalizado (GLM). Se muestran en negritas los valores estadísticamente significativos.
| Modelo Abundancia de presas |
Z | P |
|---|---|---|
| Humedad (%) | 10.32 | < 0.001 |
| Luz (lux) | -0.72 | 0.47 |
| Longitud de flores (cm) | -3.80 | < 0.001 |
| Número de pedúnculos | 6.23 | < 0.001 |
| Densidad de individuos | -7.13 | < 0.001 |
| Número de semillas fértiles | 9.73 | < 0.001 |
| Viabilidad de semillas (%) | -17.86 | < 0.001 |
| Devianza D2 | 82.45 % | |
| AIC | 310.54 |
Tabla 5 Efecto de las variables abióticas, bióticas, morfológicas y reproductivas en la diversidad taxonómica de presas en las hojas de P. moranensis, a través de un modelo lineal generalizado (GLM). Se muestran en negritas los valores estadísticamente significativos.
| Modelo Diversidad taxonómica de presas |
Z | P |
|---|---|---|
| Humedad (%) | -1.98 | < 0.05 |
| Luz (lux) | -0.04 | 0.97 |
| Longitud de flores (cm) | -0.10 | 0.92 |
| Número de pedúnculos | 0.24 | 0.81 |
| Densidad de individuos | -1.37 | 0.17 |
| Número de semillas fértiles | 2.02 | < 0.05 |
| Viabilidad de semillas (%) | -1.30 | 0.19 |
| Devianza (D2) | 70.78 % | |
| AIC | 26.264 |
Tabla 6 Efecto de las variables abióticas, bióticas, morfológicas y reproductivas en el número de semillas fértiles, a través de un modelo lineal generalizado (GLM). Se muestran en negritas los valores estadísticamente significativos.
| Modelo No. Semillas fértiles |
Z | P |
|---|---|---|
| Humedad (%) | 2.57 | < 0.05 |
| Luz (lux) | 2.14 | < 0.05 |
| Longitud de flores (cm) | -3.99 | < 0.001 |
| Número de pedúnculos | 1.04 | 0.30 |
| Densidad de individuos | 5.39 | < 0.001 |
| Viabilidad de semillas (%) | 21.84 | < 0.001 |
| Devianza (D2) | 74.10 % | |
| AIC | 463.83 |
Discusión
Este estudio presenta el análisis de algunas variables bióticas y abióticas asociadas al hábitat de la planta carnívora P. moranensis var. neovolcanica en Tlaxcala. En cuanto a los factores bióticos, se registró una composición taxonómica similar de presas con las reportadas en poblaciones de P. moranensis en el estado de Puebla y Morelos, aunque la estructura de las abundancias es diferente (Alcalá & Domínguez 2003, Díaz-Osorio et al. 2016). Por el contrario, la composición taxonómica de los visitadores florales en esta investigación es diferente a lo publicado por Villegas & Alcalá (2018). El único grupo similar son los lepidópteros del género Autochton (Hesperidae). Esto sugiere que aunque las poblaciones de P. moranensis se ubican en la misma región (Faja Volcánica Transmexicana) la composición de visitadores florales puede variar entre sitios debido a la gran diversidad de microambientes causada por el relieve de esta región, lo cual pudiera influir en la composición y estructura del ensamble de los visitadores florales (Morrone 2014). Es interesante resaltar las familias Phoridae, Sciaridae y Ceratopogonidae (Tabla 3), estos dípteros fueron atraídos a las flores, pero también se registraron como presas importantes de la planta. En otras especies carnívoras de los géneros Drosera y Dionaea se ha propuesto que la altura de la flor incrementa las visitas de polinizadores, en las mismas, la distancia entre las trampas de las hojas y la flor, o la coloración de la hoja contribuyen de manera importante a que los polinizadores no sean capturados (Jürgens et al. 2015, Youngsteadt et al. 2018). En estudios que han analizado el síndrome floral, las características florales de P. moranensis corresponden a polinización por lepidópteros, la flor presenta una espuela con nectarios que pueden ser alcanzados por estructuras bucales alargadas; lo cual explica que la planta no contenga alguna característica que evite que los dípteros sean capturados (Alcalá & Domínguez 2003, Villegas & Alcalá 2018, Lustofin et al. 2020a). Además, en P. moranensis se ha reportado la presencia de tricomas glandulares en la epidermis exterior de la espuela, esto implica que esta estructura tenga la posibilidad de captura de artrópodos pequeños a causa de la secreción mucilaginosa de este tipo de glándulas, sustancia en la cual quedan adheridas las presas (Lustofin et al. 2020b).
La relación positiva entre las presas más abundantes de este estudio (Collembola y Diptera) y el factor abiótico de humedad ha sido demostrada en el análisis basado en el modelo de abundancia de presas (Tabla 4), destaca la importancia de esta condición del ambiente por estar relacionada con el ciclo reproductivo de colémbolos pero también de dípteros (Chapman et al. 2013), en los hábitats en que prevalecen las plantas carnívoras la humedad también se considera un factor importante junto con otros como la luz y suelos pobres en nutrientes (Givnish et al. 2018). En el modelo de diversidad taxonómica de presas (Tabla 5) la humedad tuvo una relación significativa, se ha propuesto que la abundancia relativa de los diferentes taxones varía entre sitios, así Diptera y Acari son proclives a ser más abundantes en ambientes húmedos (Alcalá & Domínguez 2003).
El número de pedúnculos florales y el número de semillas fértiles presentaron también relación positiva significativa para el modelo de abundancia de presas, se ha propuesto que la disponibilidad de nutrientes provenientes de las presas permite mayor inversión de recursos para la reproducción en plantas carnívoras (Thorén & Karlsson 1998, Ellison 2020, Adamec et al. 2021). En el mismo modelo, las relaciones negativas y significativas se presentaron entre la abundancia de presas y longitud de flores puede explicarse por la respuesta fisiológica de las plantas ante la disponibilidad de nutrientes, en plantas no carnívoras, al disponer de menos nutrientes, se induce el desarrollo de estructuras reproductivas (Wada & Takeno 2010). El efecto negativo de la densidad de individuos sobre la abundancia de presas se relaciona con la competencia intraespecífica de las poblaciones de P. moranensis, se esperaría mayor abundancia de presas en densidades de plantas relativamente bajas, lo cual ha sido documentado en la planta carnívora Nepenthes gracilis (Lam et al. 2018). Es contrastante que la viabilidad presente una relación negativa con las presas (Tabla 4 y 5), considerando que las poblaciones de P. moranensis producen más de 500 semillas por cápsula (Tabla 1), las semillas deberían disponer de reservorios energéticos para germinar y se asumiría baja consanguinidad en las poblaciones, con lo cual se incrementaría su viabilidad (Villegas & Alcalá 2018), en el tercer modelo (Tabla 6) se demostró la relación significativa semillas fértiles-viables. El porcentaje de germinación promedio fue superior al reportado por Pérez-Sánchez et al. (2017) para P. moranensis (4 %) así como el reportado por Gonçalves et al. (2008) para P. lusitánica (20 %), aunque más aproximado al que reportó Villegas et al. (2020) con diferente medio de cultivo. La luz no fue un factor significante en el modelo de abundancia de presas, en contraparte, Alcalá & Domínguez (2003) encontraron que en P. moranensis la disponibilidad de presas se incrementa en sitios más sombreados y húmedos.
Los resultados de este estudio sugieren que la interacción entre los factores abióticos actuales y los caracteres morfológicos, y los factores bióticos determinan la prevalencia de las poblaciones de P. moranensis en Tlaxcala. Sin embargo, en las plantas carnívoras se ha propuesto que la disminución en la idoneidad del hábitat por el cambio climático produce declives en las poblaciones, sugiriendo que muchas podrían estar bajo amenaza de extinción local (Fitzpatrick & Ellison 2018). Por lo anterior, este estudio plantea la importancia de la conservación de los hábitats en la Faja Volcánica Transmexicana, y la necesidad de realizar estudios que incluyan los factores antropogénicos, así como los cambios históricos de las variables abióticas (e.g., temperatura, precipitación, humedad) que podrían estar relacionados con la persistencia futura de la especie.
Material Suplementario
El material suplementario de este artículo puede ser consultado aquí: https://doi.org/10.17129/botsci.2927

