











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Las cuencas hidrográficas constituyen un mosaico complejo de ecosistemas, donde se observan los vínculos entre los territorios de las zonas altas y bajas, cuyas externalidades transportadas por los cursos de agua crean una conexión física entre poblaciones alejadas. Los territorios de las cuencas otorgan bienes y servicios ambientales invaluables para nuestra existencia, como son el suministro de agua dulce, la regulación del caudal de los ríos, el mantenimiento de los regímenes hidrológicos naturales, la regulación de la erosión o la respuesta a eventos naturales extremos, entre otros (Cotler 2010).
En México, las condiciones orográficas y climáticas han moldeado las cuencas en una gran diversidad de tamaños y formas, dando lugar a sistemas hidrográficos tan complejos como la cuenca del río Bravo, o sencillos, como la cuenca del río Mexcalhuacán (Cotler 2010). En particular la región de los ríos Grijalva y Usumacinta, en el sureste de México, es una de las zonas ecológicas con más alta diversidad biológica y cultural del territorio mexicano, sus ecosistemas albergan el 64 % de la biodiversidad nacional conocida según datos del Instituto Nacional de Ecología y Cambio Climático (INECC 2019). Además, es relevante por los servicios ecológicos o ambientales de importancia económica directa; los volúmenes de carbono capturado en las masas forestales, la captación y aportación de agua y nutrientes a las tierras del norte de Chiapas y Tabasco, así como a las zonas estuarinas de importancia para las pesquerías en la costa del Golfo de México (Mifsut & Castro 2010).
Los humedales y las comunidades vegetales de la cuenca Usumacinta-Grijalva son considerados como un área prioritaria para la conservación de la biodiversidad a nivel mundial, debido, en parte, a los servicios ambientales que proveen y a la elevada concentración de especies que mantienen por mencionar algunos grupos como ictiofauna con 172 especies, al menos 328 especies de aves, 180 especies de anfibios y 200 de reptiles (Lot & Novelo 1988, López-Hernández & Maldonado-Mares 1992, Arizmendi & Márquez-Valdelamar 2000, Córdova-Ávalos et al. 2009, Lot et al. 2015, INECC 2019). La región en la que se localiza la cuenca del Grijalva contiene la superficie más alta de inundación del país y alberga uno de los humedales más importantes de Mesoamérica, considerado así, por la elevada diversidad de plantas (alrededor de 20,000 especies) que posee (INE 2000, Novelo & Ramos 2005, Rzedowski 2006, INECC 2019) y ser una de las regiones terrestres, marinas e hidrológicas prioritarias a nivel nacional (Arriaga-Cabrera et al. 1998, Arriaga-Cabrera et al. 2000a, 2000b).
Los procesos de cambio en los recursos naturales dentro de la cuenca son constantes, en este sentido, el cambio en el uso del suelo se considera la principal causa de la pérdida y degradación de numerosos hábitats naturales amenazando la persistencia de especies y con ello el declive de la biodiversidad (Palomeque-De la Cruz et al. 2017). Cuevas et al. (2010a) mencionan que la pérdida y degradación de la vegetación natural, así como la velocidad a la que ocurren estos cambios, desencadenan procesos negativos a distintas escalas espaciales y temporales, comprometiendo la estabilización del suelo, la regulación del volumen, la periodicidad de los caudales y la purificación e infiltración del agua.
La extensión del área de distribución de la vegetación es una medida básica que permite conocer el estado ambiental de la cuenca. Leal-Nares et al. (2012) mencionan que esta medida se asocia con los modelos de distribución potencial y nicho ecológico de las especies. Suárez-Mota et al. (2018) afirman que los modelos de distribución potencial son ampliamente utilizados para estimar y predecir cambios en la distribución de una especie en respuesta a los diferentes procesos de cambio global. Debido a ello, en los últimos años la modelación de distribución potencial ha tomado gran auge por su utilidad como soporte en la toma de decisiones orientada a la gestión y conservación de las especies y áreas de conservación (Phillips et al. 2006, Peterson et al. 2011).
En las últimas décadas, se ha incrementado el desarrollo de algoritmos en el campo de la modelación de nichos ecológicos, hasta el punto de tener distintos métodos disponibles (Illoldi-Rangel & Escalante 2008, Elith et al. 2011, Soberón et al. 2017). El algoritmo de máxima entropía (MaxEnt) es el más utilizado debido a su flexibilidad para modelar la distribución potencial y nicho ecológico de diversas especies con buen poder predictivo (Phillips et al. 2006, Peterson et al. 2011, Merow et al. 2013). MaxEnt permite caracterizar distribuciones de probabilidad cuya información está incompleta. Se basa en el principio de que la distribución estimada de una especie debe coincidir con la distribución conocida a partir de las condiciones ambientales donde ha sido observada, evitando hacer suposiciones que no sean apoyadas por los datos. El enfoque consiste en encontrar la distribución de probabilidad de máxima entropía, que es la más cercana a la distribución uniforme, condicionada por las restricciones impuestas por la información disponible sobre la distribución observada de la especie y las condiciones ambientales del área de estudio (Peterson et al. 2011). Esta practicidad se debe a los insumos que necesita, ya que MaxEnt utiliza la información y bases de datos disponibles en herbarios y museos y la asocia con variables ambientales (Elith et al. 2005, Elith et al. 2006, Elith et al. 2011, Suárez-Mota et al. 2018). Los modelos de distribución de especies son representaciones cartográficas ideales de un espacio para la presencia de una especie en función de las variables con las que interactúan (Mateo et al. 2011, Antúnez et al. 2018, Suárez-Mota et al. 2018, Suárez-Mota & Villaseñor 2020).
En México se están deteriorando bosques, desiertos y humedales, una superficie extensa de reservas naturales se ha transformado en zona urbana y más de 30 % de los humedales han desaparecido (Torres‐Vera et al. 2009, Zepeda-Gómez et al. 2012). En la cuenca del Grijalva-Usumacinta el área de las selvas ha disminuido de forma significativa en las últimas décadas(Kolb & Galicia 2012). La pérdida de la cobertura vegetal y de los humedales en la cuenca se ha relacionado con el crecimiento de la ciudad, la deforestación con fines de uso ganadero y la explotación forestal y petrolera (Kolb 2013). El cambio de uso del suelo modificó los ciclos naturales de inundación, el trastorno de los volúmenes de agua en los ríos y zonas de anegación temporal y dinámica trófica de los humedales, lo que llevó a la fragmentación y pérdida del hábitat, así como a la disminución de la biodiversidad en las últimas décadas (Pinkus-Rendón & Contreras-Sánchez 2012, Sánchez et al. 2015). Sin embargo, la flora de ribera distribuida en la cuenca no ha sido analizada en cuanto sus patrones de distribución y el conocimiento de los factores ambientales que la determinan no han sido evaluados.
Los modelos de distribución potencial son usados para explicar los factores ambientales que determinan la distribución de las especies (Antúnez et al. 2018). La modelación de la distribución geográfica de las especies permite identificar patrones de distribución generales de la biodiversidad, por ejemplo, plantas (Zaluzania sp.) en las zonas áridas de México, lo que facilita decidir acerca de las prioridades de conservación a gran escala (Chapman & Busby 1994, Franklin 1995, Austin 1998, 2002, Guisan & Zimmermann 2000, Elith & Burgman 2003, Scott et al. 2002, Suárez-Mota & Villaseñor 2020).
Las plantas adaptadas a un régimen climático particular con frecuencia tienen morfologías similares o ciertos tipos de formas de crecimiento (Cain 1950). Los árboles constituyen, por ejemplo, el biotipo más conspicuo y, fisonómicamente hablando, el más importante, pues por lo general domina en muchas comunidades vegetales tanto por su densidad como por su biomasa (Villaseñor & Ibarra 1998). Se han utilizado los elementos del clima para delimitar grandes unidades de vegetación, como es el caso del sistema de las zonas de vida elaborado por Holdridge (1967), o bien, para relacionar las adaptaciones de los grandes biomas con ciertos intervalos de variables ambientales (Cox et al. 1976). De manera particular, cada especie tiene su propio perfil climático, por lo que el análisis de las variables que lo determinan puede servir para cuantificar las diferencias en sus áreas de distribución. Para evaluar el papel de la riqueza florística en la composición regional y su relevancia en la conservación de los bosques de la subcuenca Bajo Río Grijalva, este trabajo se plantea el objetivo conocer la distribución potencial de especies de ribera dentro de la subcuenca a partir de modelos de distribución potencial de árboles ribereños y reconocer la similitud climática de las especies seleccionadas, para así poder analizar como las actividades humanas han alterado las condiciones ambientales que propician el establecimiento de la vegetación.
Materiales y métodos
Área de estudio. La subcuenca Bajo Río Grijalva localizada al sureste de México, se ubica dentro de las coordenadas -93º 39′ 59″ y -91º 43′ 32″ de longitud oeste, 18º 38′ 39″ y 16º 45′ 20″ de latitud norte, abarcando un área total de 2,306,181 ha cubriendo un total de 64 municipios, de los cuales 14 pertenecen al estado de Tabasco, uno a Veracruz y 49 al estado de Chiapas; colinda al norte con el Golfo de México, al sur con la Sierra Norte de Chiapas, al este con la República de Guatemala y al oeste con los estados de Oaxaca y Veracruz (Figura 1).

Caracterización morfométrica de la cuenca. Los parámetros morfométricos, forma, relieve y red hidrográfica, área, perímetro, cotas máximas y mínimas, curva hipsométrica y pendiente promedio de la cuenca, se obtuvieron con el Sistema de Información Geográfica (SIG) implementado en el software ArcGis 9.3 (ESRI 2011), utilizando un modelo digital de elevación de 120 m de resolución (INEGI 2019).
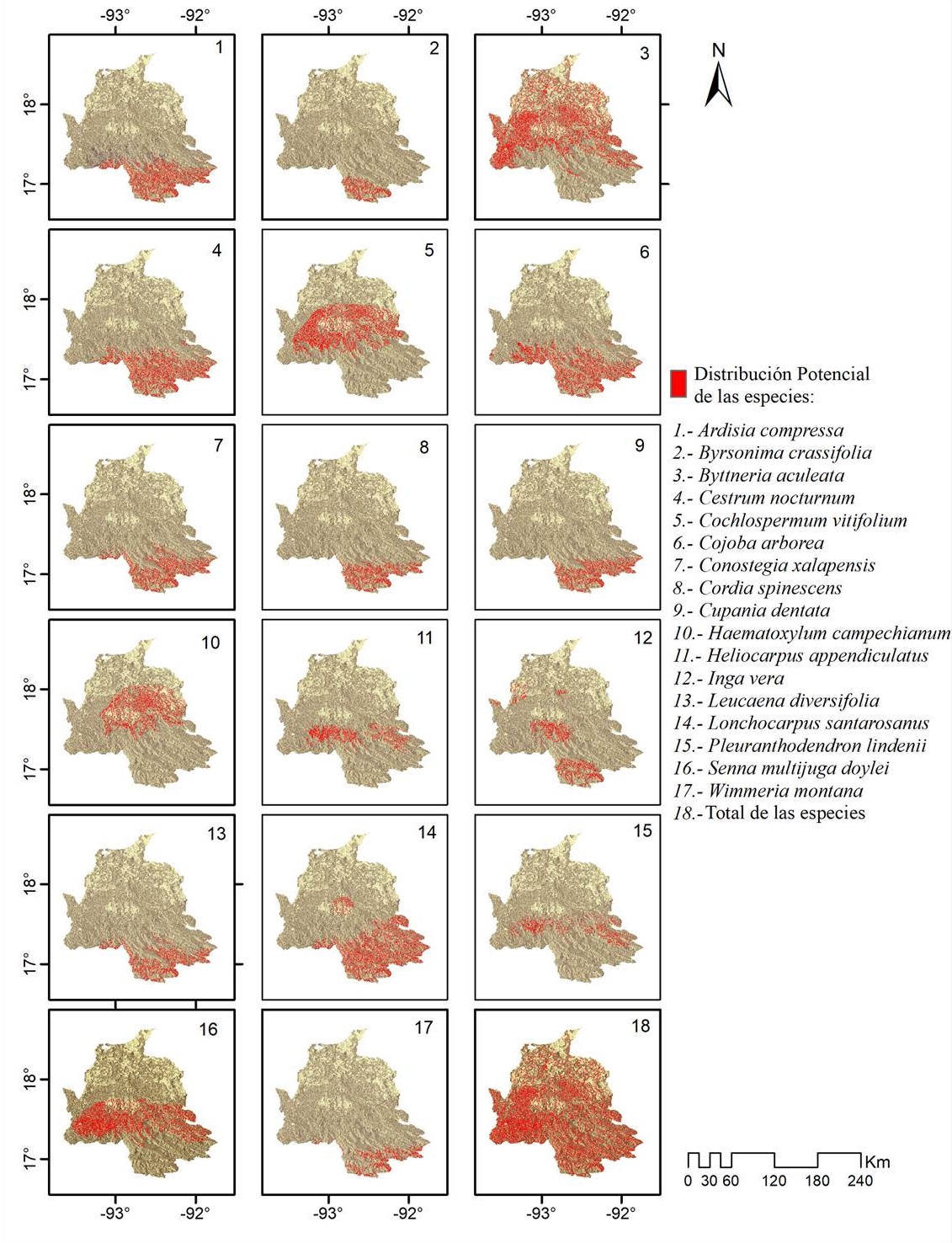
Modelos de distribución potencial y selección de especies. La recopilación de datos de ocurrencia de las especies georreferenciadas se obtuvo a través de colecciones y bases de datos de la Red Mundial de Información sobre Biodiversidad (http://www.conabio.gob.mx/remib/doctos/remib_esp.html) y de la Unidad de Informática y Biodiversidad (unibio.unam.mx) del Instituto de Biología de la Universidad Nacional Autónoma de México (UNAM). La información obtenida se depuró seleccionando especies arbóreas consideradas como vegetación de ribera, distribuidas dentro del área contigua a los cuerpos de agua y que además tuvieran como mínimo cinco registros por especie, los cuales se evaluaron para verificar su correcta determinación y georreferenciación, dando como resultado un total de 17 especies seleccionadas de un total de 598 que fueron revisadas y corregidas de las bases de datos disponibles (Tabla 1).
Tabla 1 Listado de especies arbóreas ribereñas analizadas. Se indica el total de registros, el valor del área bajo la curva (AUC) y umbral (Um) derivados de los modelos de distribución potencial.
| No. | Especie | Registros | AUC | Um |
|---|---|---|---|---|
| 1 | Ardisia compressa Kunth. | 6 | 0.846 | 0.467 |
| 2 | Byrsonima crassifolia (L.) Kunth. | 7 | 0.61 | 0.596 |
| 3 | Byttneria aculeata (Jacq.) Jacq. | 8 | 0.594 | 0.5 |
| 4 | Cestrum nocturnum Duss. | 13 | 0.743 | 0.492 |
| 5 | Cochlospermum vitifolium (Willd.) Spreng. | 6 | 0.86 | 0.405 |
| 6 | Cojoba arborea (L.) Britton & Rose. | 6 | 0.771 | 0.45 |
| 7 | Conostegia xalapensis (Bonpl.) D. Don. | 14 | 0.646 | 0.561 |
| 8 | Cordia spinescens L. | 9 | 0.877 | 0.531 |
| 9 | Cupania dentata DC. | 8 | 0.915 | 0.404 |
| 10 | Haematoxylum campechianum L. | 7 | 0.816 | 0.546 |
| 11 | Heliocarpus appendiculatus Turcz. | 6 | 0.968 | 0.289 |
| 12 | Inga vera Kunth. | 16 | 0.894 | 0.478 |
| 13 | Leucaena diversifolia (Schltdl.) Benth. | 10 | 0.912 | 0.407 |
| 14 | Lonchocarpus santarosanus Donn. Sm. | 9 | 0.819 | 0.433 |
| 15 | Pleuranthodendron lindenii (Turcz.) Sleumer | 9 | 0.955 | 0.479 |
| 16 | Senna multijuga doylei (Britton & Rose) H.S. Irwin & Barneby | 8 | 0.855 | 0.283 |
| 17 | Wimmeria montana Lundell | 6 | 0.949 | 0.528 |
Delimitación del área de movilidad (M). La delimitación del área de movilidad (M), según el diagrama BAM propuesto por Soberón & Peterson (2005) y Soberón et al. (2017), se obtuvo a partir de la información sobre el mapa de cuencas hidrográficas a escala de 1:250,000, publicado por el Instituto Nacional de Estadística y Geografía (INEGI 2007). Para cada cuerpo de agua (ríos y arroyos) se generó un área de amortiguamiento de 100 metros a partir de los bordes de los cuerpos de agua con la finalidad de tener un área homogénea a lo largo de los ríos puesto que no en todo su recorrido tienen la misma amplitud (Figura 2). El polígono formado con este amortiguamiento se empleó como área M para la modelación de las especies analizadas.

Figura 2 Cuerpos de agua con una zona de amortiguamiento de 100 m. Este polígono se empleó como área de movilidad para los modelos de distribución potencial de las especies analizadas.
Implementación de Máxima Entropía (MaxEnt). Las variables climáticas utilizadas para la generación de los modelos se obtuvieron de las 19 variables de clima propuestas por Cuervo-Robayo et al. (2013; Tabla 2) disponibles en el geoportal de la Comisión Nacional para el Uso y Conservación de la Biodiversidad (CONABIO) y tres variables topográficas. Con esta información se realizó un análisis de componentes principales (ACP) empleando el paquete vegan implementado en R (R Core Team 2008), para seleccionar las variables con menor correlación en el área (M) donde se realizaron los modelos. Los modelos de distribución potencial se obtuvieron usando el algoritmo de Máxima Entropía (MaxEnt) 3.3.3 k. Los modelos se hicieron de forma homogénea con el propósito de analizar la distribución potencial de especies arbóreas ribereñas, así como la similitud de los nichos que las especies analizadas comparten. Por lo cual, la configuración de MaxEnt se realizó con el índice de salida logístico. Para cada modelo se empleó el 75 % de los datos como entrenamiento y el 25 % restante como datos de prueba. La evaluación de los modelos se hizo con los resultados del análisis del Área Bajo la Curva (AUC por sus siglas en inglés) de la Característica Operativa del Receptor (ROC por sus siglas en inglés; Tabla 1). Los parámetros utilizados para obtener dichos modelos fueron: el multiplicador de regularización (regularization multiplier) = 1, número máximo de puntos de fondo (maximum number of background points) = 10,000, límite de convergencia (convergence limit) = 0.00001 y número máximo de repeticiones (maximum iterations) = 1,000. Por último, los archivos tipo ASCII generados por MaxEnt fueron importados a un Sistema de Información Geográfica (ArcGis 9.3) para su edición y generación de los modelos en modo binario (presencia-ausencia) empleando como umbral de corte el Maximum training sensitivity plus specificity. Una vez que se obtuvieron los modelos y evaluando los resultados de la prueba Jackniffe implementada en Maxent, se seleccionaron las variables que tuvieron mayor contribución en cada modelo de las especies, así se repitió el modelo solo con estas variables.
Tabla 2 Parámetros bioclimáticos utilizados para generar los modelos de distribución potencial. Valores de carga resultado del Análisis de Componentes Principales (ACP) que explican la mayor varianza. *Las variables con menor colinealidad seleccionadas para los modelos de distribución potencial de las especies.
| Acrónimo | Descripción | CP 1 | CP 2 | CP 3 |
|---|---|---|---|---|
| BIO1 | Temperatura promedio anual | -0.24 | 0.19 | -0.05 |
| BIO2 | Oscilación diurna de la temperatura | 0.21 | 0.04 | -0.26 |
| BIO3 | Isotermalidad | 0.25 | -0.1 | -0.03 |
| BIO4 | Estacionalidad de la temperatura estacional | -0.24 | 0.14 | -0.01 |
| BIO5 | Temperatura máxima del periodo más cálido | -0.24 | 0.2 | -0.09 |
| BIO6 | Temperatura mínima del periodo más frío | -0.24 | 0.17 | -0.02 |
| BIO7* | Oscilación anual de la temperatura | -0.12 | 0.28 | -0.4 |
| BIO8* | Temperatura promedio del trimestre más húmedo | -0.24 | 0.19 | -0.07 |
| BIO9* | Temperatura promedio del trimestre más seco | -0.25 | 0.13 | 0.02 |
| BIO10 | Temperatura promedio del trimestre más cálido | -0.24 | 0.18 | -0.05 |
| BIO11* | Temperatura promedio del trimestre más frío | -0.24 | 0.19 | -0.06 |
| BIO12 | Precipitación anual | -0.22 | -0.27 | 0.01 |
| BIO13 | Precipitación del periodo más húmedo | -0.21 | -0.27 | -0.08 |
| BIO14 | Precipitación del periodo más seco | -0.22 | -0.27 | 0 |
| BIO15* | Estacionalidad de la precipitación | 0.22 | 0.17 | -0.15 |
| BIO16 | Precipitación del trimestre más húmedo | -0.22 | -0.26 | -0.01 |
| BIO17 | Precipitación del trimestre más seco | -0.22 | -0.28 | 0.03 |
| BIO18* | Precipitación del trimestre más cálido | -0.16 | -0.35 | -0.1 |
| BIO19 | Precipitación del trimestre más frío | -0.23 | -0.22 | 0.1 |
| MDT* | Altitud m snm | 0.24 | -0.17 | 0.04 |
| ASPECT* | Orientación | 0.03 | -0.09 | -0.66 |
| SLOPE* | Pendiente | 0.05 | -0.24 | -0.52 |
| Valores de los Componentes Principales: | ||||
| Desviación estándar | 3.9 | 1.85 | 0.06 | |
| Varianza acumulada | 0.69 | 0.85 | 0.90 | |
Similitud ambiental de especies. Con los valores de las variables ambientales que determinan la distribución de las especies se construyó una matriz de datos para hacer un análisis de conglomerados y obtener la similitud ambiental entre especies. Para lo cual, se empleó la métrica de distancias de Gower integrado en el programa PATN (Gower 1971, Belbin 1989). La clasificación final se hizo mediante un agrupamiento promedio no ponderado (UPGMA por sus siglas en inglés).
Resultados
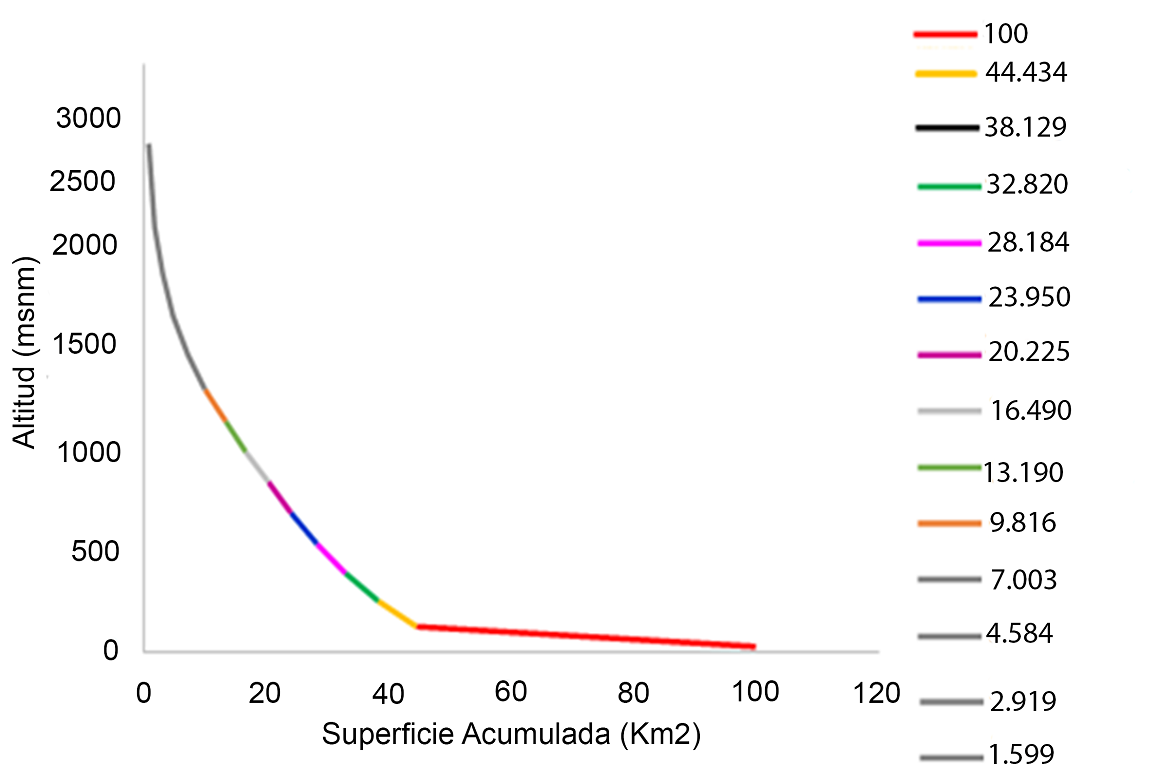
Parámetros morfométricos. Las características morfométricas del área de estudio se explican a partir de parámetros mencionados en la Tabla 3. El índice obtenido para el coeficiente de compacidad (Cc) fue 2.324, de acuerdo con Campos-Aranda (1998), indica que la subcuenca Bajo Río Grijalva tiene forma oval-oblonga a rectangular alargada lo que puede intensificar la fuerza de las avenidas al menos en las desembocaduras y la onda de crecida que puede manifestarse fuertemente antes del cauce principal (Maldonado-De-León et al. 2001).
Tabla 3 Parámetros morfométricos de la subcuenca Bajo Grijalva.
| Parámetros | Valor |
|---|---|
| Coeficiente de compacidad | 2.324 |
| Factor de forma | 0.55 |
| Coeficiente de masividad | 0.48 |
| Pendiente media de la cuenca | 12.1 |
| Elevación media | 364.62 m |
| Orden de la corriente | 4 órdenes |
| Densidad de drenaje | 3.02 km/km2 |
| Tiempo de concentración | 68.98 h |
Para el factor de forma se aplicó la fórmula de Horton (1945), el índice obtenido fue 0.55 (Tabla 3) que corresponde a una cuenca elongada lo que hace al Bajo Grijalva una zona moderadamente lenta entre el momento de la precipitación y el de crecida en la desembocadura, pero al mismo tiempo, más alto riesgo de inundaciones. El coeficiente de masividad obtenido fue de 0.48, lo que indica que la zona de la cuenca es montañosa. Sin embargo, la forma se considera una característica cuyo concepto es complejo con muchos atributos específicos lo que hace difícil caracterizar de manera real este parámetro.
La distribución del análisis hipsométrico, realizado en este trabajo, indicó que, la subcuenca Bajo Río Grijalva se encuentra en una etapa intermedia entre la fase de equilibrio relativo o de madurez, evolucionado hacia la etapa de madurez de acuerdo con el análisis hipsométrico (Figura 3), por su ubicación el área de estudio debe su estado de madurez principalmente a procesos de erosión (Guerra & González 2002). La pendiente media de la cuenca fue de 12.1 %, de lo cual se deduce que el relieve es accidentado y favorece los procesos de escorrentía y erosión.
Modelos de distribución potencial . Se obtuvieron 148 registros de las 17 especies seleccionadas (Tabla 1). Con los registros se generaron los mapas de distribución potencial de las 17 especies, 11 (64.71 %) de ellas se distribuyen en la parte con mayor altitud (2,000 a 3,000 m snm), 13 (76.47 %) en la parte media (1,000 a 2,000 m snm), y todas (100 %) en la parte con menor altitud del área de estudio (0 a 1,000 m snm); de estos datos se reconoce que dos especies (Byttneria aculeata y Heliocarpus appendiculatus) son las únicas que se registran en las partes con mayor altitud (Figura 4). Los valores del Área Bajo la Curva (AUC) obtenidos para 12 especies (70.06 %) fueron mayores de 0.8 lo cual indica que son modelos adecuados a los registros de las especies y su relación con las variables que determinan su distribución potencial (Tabla 1).
Las 17 especies mostraron un comportamiento similar en relación con las variables ambientales. La variable que mostró mayor influencia en la distribución de las especies fue la pendiente (slope), donde Pleuranthodendron lindenii presentó la relación más alta con esta variable. Los valores obtenidos con el ACP mostraron cargas muy similares entre las variables. Para los modelos realizados se seleccionaron las variables con mayor valor de carga en los primeros tres componentes por explicar la mayor varianza. De aquellas variables que tienen un mismo valor en el primer componente se seleccionó la que tuviera el mayor valor en los siguientes componentes; los valores más altos corresponden a la orientación y la pendiente en el componente tres (Tabla 2). Con estos valores y considerando los resultados de la prueba de Jackknife implementada en MaxEnt, fueron seleccionadas las variables de mayor contribución para realizar los modelos. Las variables que determinaron en gran parte la distribución de las especies y con las que se realizaron los modelos fueron: BIO7, BIO8, BIO9, BIO10, BIO11, BIO15, BIO18, la exposición (aspect), la pendiente y la altitud (Tabla 2). Heliocarpus appendiculatus fue la especie que mayor relación tuvo con las variables ambientales dentro del área de estudio (Figura 5).

Figura 5 Se indica la contribución de las variables ambientales que determinan en la distribución de especies modeladas, valores obtenidos con la prueba de Jackknife implementada en Maxent.
Similitud ambiental de las especies ribereñas. A un nivel de similitud aproximado de 0.23 se observan cinco conglomerados, de los cuales el grupo que incluye a Byrsonima crassifolia, Conostegia xalapensis, Inga vera, Cestrum nocturnum, Lonchocarpus santarosanus y Byttneria aculeata, es el más grande al agrupar seis de las 17 especies. La especie con mayor diferencia ambiental al resto fue Haematoxylum campechianum (Figura 6).

Discusión
Los índices obtenidos en este trabajo indican que la zona de estudio presenta una forma que va de oval oblonga a rectangular oblonga, además, forma una red de drenaje que tiende a ser menos compacta y los tiempos de concentración tienden a ser mayores, disminuyendo la torrencialidad (Díaz et al. 2017). La forma de la subcuenca puede intensificar el vigor de las avenidas al menos en las desembocaduras y la onda de crecida, antes del cauce principal. Este coeficiente puede ser un indicador para prevenir inundaciones o llegadas repentinas de agua en ciertos poblados cercanos a cauces o arroyos dado que la duración de los escurrimientos al cauce principal puede ser más rápido (Viramontes-Olivas et al. 2008).
El área de estudio se caracteriza por tener en la parte alta zonas con pendientes pronunciadas mientras que en la parte media y baja predomina un relieve llano presenta áreas que inician en el encañonado de las montañas del norte de Chiapas y la presa de Peñitas, seguidas de un drenaje de las laderas del volcán Chichonal que, con otros factores tales como perdida de pendiente, estructura geológica y arrastre de sedimentos en suspensión favorecen las inundaciones dentro del área de estudio, en particular al estado de Tabasco que en el 2007 registró una inundación que afectó severamente a 17 municipios (SEGOB-CENAPRED 2012).
La curva hipsométrica obtenida muestra a la cuenca en una etapa intermedia de equilibrio relativo o de madurez. Además, indica un significativo potencial erosivo, ya que la subcuenca se considera sedimentaria (Gaspari et al. 2013). Es una cuenca accidentada con una intensa erosión regresiva, que en conjunto con su altura a nivel del mar permiten la presencia de valles y pantanos en la parte baja de la subcuenca, lo cual puede presentar un peligro de inundación en zonas aledañas al cauce (CONAGUA 2014). Se han registrado fenómenos naturales de manera aislada como deslizamientos, inundaciones y hundimientos, en ocasiones se presentan de manera conectada, es decir, la presencia de una desencadena otra. Por ejemplo, un deslizamiento puede bloquear un cuerpo de agua y éste a su vez, desbordarse y provocar una inundación en una zona de hundimientos. Tal como sucedió con el cauce del río Grijalva en el 2007, cuando un gran deslizamiento provocó inundaciones en Chiapas y Tabasco y prácticamente desapareció parte de la comunidad de Juan de Grijalva en la primera entidad (Guadarrama et al. 2010).
Se registró un tiempo de concentración de 68.98 h (3 días) lo que permite altas tasas de infiltración y alimentación del flujo subsuperficial (Viramontes-Olivas et al. 2008), sin embargo, la presencia de planicies en la parte baja representa un posible peligro de inundación en zonas aledañas al cauce principal en esta zona; en especial sí la cobertura vegetal ha sufrido cambios en su estructura (Cruz-Romero et al. 2015).
A todo lo largo y ancho de la subcuenca se presentan una gran variedad de ecosistemas acuáticos y bosques ribereños (Mifsut & Castro 2010). La relación de la vegetación ribereña con cuerpos de agua permite inferir el caudal ecológico, conocer la distribución de la vegetación y ubicar las áreas con mayor deterioro.
Los modelos generados muestran la distribución potencial de las 17 especies modeladas, 11 de ellas se encuentran en la parte alta de la subcuenca, 13 en la parte media y todas en la parte baja. La parte alta de la subcuenca es la zona con el mayor conjunto de condiciones climáticas favorables para el desarrollo de diversos tipos de vegetación, y que, a su vez sustentan los medios de subsistencia para la presencia de una gran cantidad de especies de fauna (Hamilton 2009). La presencia de especies arbóreas ribereñas en las partes media y alta, indica una mayor calidad de agua debido a que se minimiza la erosión local del suelo, se reducen los sedimentos en las masas de agua (humedales, estanques, lagos, arroyos y ríos) y se detienen o filtran otros contaminantes del agua en la hojarasca y el sotobosque (Hamilton 2009). El conjunto de condiciones ambientales y topográficas presentes en la zona alta de la subcuenca la hacen clave para el manejo integrado de todo el sistema hidrográfico, y es ahí donde se infiltra la mayor cantidad de agua alimentando a los flujos subterráneos (Garrido et al. 2010).
Los diversos diagnósticos socioeconómicos que se han realizado en la cuenca del Grijalva-Usumacinta mencionan que la población que habita dentro de la cuenca se encuentra dentro de los niveles de marginación más elevados de México (Mifsut & Castro 2010). La relación que existe entre la transformación de la vegetación y la subsistencia de las poblaciones es debida a la agricultura de subsistencia, tala de zonas boscosas para áreas de cultivo y ganadería, demanda de agua y alimentos (Bedolla 2010). Además, el crecimiento de la urbe genera presión sobre el entorno natural de la cuenca, teniendo implicaciones ambientales principalmente en la degradación de suelos, que a su vez tienen influencia directa sobre el ciclo hidrológico ya que se elimina la humedad almacenada en la masa de vegetación, se incrementa la escorrentía a costa de la infiltración del agua para recarga de acuíferos y se modifican los microclimas (Bedolla 2010).
Al analizar el perfil ambiental de las 17 especies se observó que la pendiente (slope), exposición (aspect) y la altitud (mdt) fueron las variables que determinaron en mayor proporción la distribución de las especies. Donde Haematoxylum campechianum fue la especie que presentó mayor amplitud en el rango de distribución, al presentar los valores más altos en las distintas variables analizadas. Lo anterior sugiere, que la variabilidad climática y las condiciones topográficas diferencian la distribución de las especies modeladas.
El perfil ambiental y la distribución potencial sirven para reconocer sitios importantes de concentración de especies en este estudio y particularmente ribereñas. Los usos y métodos para conocer la amplitud ambiental de una especie son variadas, por ejemplo, Leal-Nares et al. (2012), emplearon el modelo de distribución y el perfil bioclimático de Pinus martinezi para ubicar zonas para colectar semillas y establecer viveros.
La similitud de las especies con respecto a las variables ambientales y los grupos formados indican la afinidad ambiental que tienen las especies considerando los mapas de distribución potencial, es decir, las especies que se mostraban en la zona alta de la subcuenca formaron un grupo, el de mayor tamaño, mientras las que se encontraban en la zona media formaban otro grupo, caso contrario a la zona baja en la cual se formó un grupo considerando a las especies distribuidas en la zona media, la razón de este comportamiento se debe a que el área de estudio presenta poca variabilidad climática. Cabe recalcar que conforme va aumentando el nivel de similitud los grupos se van definiendo de manera más amplia, por ejemplo, a un nivel de similitud de 62 %, se forman dos grupos de los cuales el primero integra 12 especies con distribución en la zona alta y media del área de estudio. El segundo grupo está conformado por cinco especies con distribución en la zona baja, media y alta del área de estudio. Las especies presentes en este grupo mostraron las amplitudes más altas.
Los resultados obtenidos proporcionan información valiosa para la ejecución de posteriores estrategias de estudio, tomando en cuenta diversos factores ambientales que determinan la amplitud de las especies, esta información permite tomar decisiones para la incorporación de especies a zonas que sean idóneas para su distribución y de esta manera contribuir a la restauración con actividades de conservación y reforestación que favorezcan al caudal ecológico.
El área de distribución potencial de las especies arbóreas ribereñas modeladas en la subcuenca Bajo Río Grijalva cubre zonas altas, medias y bajas en toda el área de estudio, por lo que se infiere que dicha zona presenta un caudal ecológico alto.
El intervalo de las variables ambientales que delimitan la distribución de las especies arbóreas modeladas es similar, sin embargo, el perfil ambiental muestra que las variables de terreno son determinantes en la distribución de las especies.

