











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Sexual systems of trees may determine their degree of reproductive vulnerability to forest fragmentation. It is predicted that self-compatible species could have reproductive assurance through self-pollination, which will allow them to persist successfully in fragmented habitats (Quesada et al. 2011). In contrast, dioecious and obligate-outcrossing hermaphrodites will show strong dependence not only on highly mobile pollen vectors, but also on the availability of other reproductive individuals for mating and seed production (Breed et al. 2015). Hence, obligate outcrossing species will be highly vulnerable to the loss of pollinators and reduced population size triggered by forest fragmentation (Aguilar et al. 2006). For example, a study in fragments of a semi-deciduous forest in Brazil revealed that most trees are pollinated by small insects with restricted mobility (Montoya-Pfeiffer et al. 2018). Obligate outcrossing species, as dioecious, would be sensitive to habitat degradation because they naturally occur at lower densities and are pollinated by small insects. Then, conservation actions may favor the maintenance of all sexual systems, but restoration intervention may be necessary to recover populations of obligate outcrossing species in disturbed areas.
Even when worldwide there is a high frequency of hermaphroditism among angiosperms (Campbell 2000), in the tropical dry forest, the percentage of obligate-outcrossing species may be higher than in other ecosystems (Aguilar et al. 2006). The family Fabaceae, a very important one in the tropics, include only hermaphrodite species (Banda-R et al. 2016) but a review for 345 species in 78 families showed that > 40 % of hermaphrodites show self-incompatibility (Goodwillie et al. 2005); further, this percentage may be higher for tropical trees given inbreeding depression (Eckert et al. 2010, Breed et al. 2015). Then, given a high proportion of dioecious species in tropical dry forest and a high proportion of outcrossing species within Fabaceae species, the most important mating system for dry forest floras is outcrossing.
Ecological restoration aims to initiate or accelerate the recovery of a damaged ecosystem (Gann et al. 2019). Minimal restoration intervention, also referred to as unassisted forest regeneration includes actions to remove chronic disturbance so natural succession may take place (Chazdon & Guariguata 2016, Martínez-Garza et al. 2016). Maximal intervention, also called assisted succession, involve restoration plantings (Chazdon & Guariguata 2016, González-Tokman et al. 2018). Either or both of these restoration interventions may favor not only the recovery of species but the functional diversity of reproductive systems of plants, and the pollen vectors they depend on for sexual reproduction (Menz et al. 2011). Further, restored areas may decrease hostility and increase permeability of permanent agricultural landscapes to the movement of plants and animals (Vandermeer & Carvajal 2001, Howe & Martínez-Garza 2014) but they may still show a strong reduction of tree species pollinated by small insects (Montoya-Pfeiffer et al. 2018). Therefore, it is not clear when unassisted forest regeneration will be sufficient, or when restoration plantings will be required to recover the ecological interactions lost due to degradation, including the functional component of sexual systems.
Here, we assessed the richness and density of dioecious, monoecious, and hermaphrodite tree species in four habitats: disturbed habitats, 8-years old restoration settings (sites under unassisted forests regeneration and plantings) and in conserved habitats of a tropical dry forest in central Mexico. The trees we evaluate in the plantings include plants that were intentionally planted as well as those that recruited naturally. We aimed to test the hypothesis that due to forest fragmentation, providing sufficient time has elapsed since disruption, species that produce seeds by outcrossing (dioecious and monoecious) would have lower richness and density in disturbed areas and would be better represented in conserved and restored habitats. We also expected a higher richness and density of hermaphrodite species at all habitats given that Fabaceae, the most important family in the dry forest, have this sexual system. Finally, conserved and restored habitats are expected to be the most similar in their composition of sexual systems, especially of self-compatible hermaphrodite species. By measuring sexual systems in permanent agricultural landscapes, we can guide conservation, management, and restoration actions to maintain and recover self-incompatible tree species and their ecological interactions in tropical dry forests.
Materials and methods
Study site. This study was carried out near the community of El Limón de Cuauchichinola, in the Sierra de Huautla Biosphere Reserve, state of Morelos, central Mexico (18° 20’ 10” N, 98° 51’ 20” W). The Biosphere Reserve comprises 59,030 ha with an altitudinal range from 700 to 2,200 m asl. The main vegetation type there is seasonally tropical dry forest (STDF; Dirzo et al. 2011, Banda-R et al. 2016), also called tropical deciduous forest (Rzedowski 2006). Mean annual temperature is 24.5 ºC and average annual rainfall is 840 mm (1980-2001; CONAGUA, Gerencia Regional Balsas, unpublished data), with ~ 90 % falling between late May and October. During the dry season (November to April), most of the trees lose their leaves, and soils are shallow (< 30 cm in depth) with sandy-loam texture (CONANP 2005). In the Reserve, the most important plant families for the number of species are Fabaceae, Poaceae, Asteraceae, Burseraceae, and Cactaceae (CONANP 2005). Most of the trees in this forest have compound leaves and some reach 8 to 12 m in height, with convex or flat canopies (Rzedowski 2006).
Land use history. The landscape at the Biosphere Reserve is a mosaic of conserved and secondary forest surrounded by agricultural land and small towns. El Limón de Cuauchichinola has an area of 4,256 ha, of which 12 % is secondary forest, 56 % is conserved forest, 13 % is dedicated to agriculture and 19 % is degraded land (De León-Ibarra 2005). The secondary forest resulted from the clearing of large tracts of forest ca. 50 years ago (~ 1970), which were used for maize cultivation for ca. 6 years, and later abandoned (G. Pacheco, local farmer, pers. comm.). In this habitat, the hermaphrodite species Acacia cochliacantha Humb. & Bonpl. ex Willd. (Fabaceae), Ipomoea pauciflora M. Martens & Galeotti (Convolvulaceae), Acacia farnesiana (L.) Willd. (Fabaceae), and Mimosa benthamii J.F. Macbr. (Fabaceae) are dominant (Martínez-Garza et al. 2011). These secondary forests are chronically disturbed by cattle ranching and extraction activities, hereafter referred to as disturbed habitat (Table S1; Figure 1A); during the rainy season, ca. 600 head of livestock are brought from neighboring towns to graze there (~ 7 head/ha). According to landowners and current research (B. Maldonado, unpublished data), cattle are maintained on farms during the dry season but goats, pigs, and horses graze there throughout the year. Wood extraction there is a common activity and villagers extract between 74 and 124 kg of firewood per trip (Monroy 1997). What we call conserved habitat corresponds to older growth forest that has never been deforested (> 80 years ago), and cattle from neighboring towns do not graze there (Table S1). Cattle from local owners occasionally do, but cattle are never found in densities higher than 1 head/ha (G. Pacheco, local farmer, pers. comm.). In these conserved forests wood extraction is seldom practiced, since the preferred tree species are not frequent (B. Maldonado, unpublished data). The most common canopy trees in the conserved forest are the hermaphrodite Conzattia multiflora (B.L. Rob.) Standl., Lysiloma acapulcense (Kunth) Benth., L. divaricatum (Jacq.) J.F. Macbr. (Fabaceae), several species of the dioecious Bursera (Burseraceae) and of the hermaphrodite Ceiba (Bombacaceae) (CONANP 2005). Regarding potential pollinators, the last review for the state of Morelos reported 4,752 species of insects; the most important orders were Coleoptera (2,362 species), Lepidoptera (778), Diptera (606) and Hymenoptera (413; Cano-Santana et al. 2020).

Figure 1 Secondary forests chronically disturbed by cattle ranching and extraction activities (A); view of exclusion established in 2006 (B); electric fence in 2011 (C); vegetation in exclusions in the rainy season of 2011 (D); juvenile of Pseudobombax ellipticum (yellow arrow) within plantings in 2011, cattle graze outside plots (E); Planted Leucaena sp. set fruits by the rainy season of 2011 (F) in Sierra de Huautla, Morelos, Mexico. Photos by Cristina Martínez-Garza.
Experimental design. To favor the movement of plant and animals in this permanent agricultural landscape, a large restoration experiment was established in January 2006. The experimental design consists of 14 50 × 50 m plots (3.5 ha) in four habitats (Table S1, Figure S1). Six plots were established in the disturbed (N = 3 plots) and conserved habitats (N = 3 plots) described above. Eight plots in former disturbed forest were excluded from livestock using electric fences and in four of them, natural succession was favored with no further intervention (hereafter, exclusions; Table S1; Figure 1B and 1C). In the other four plots, in addition to being fenced to exclude livestock, 20 native tree species from nine families were planted, 560 plants in each plot (hereafter, plantings; Martínez-Garza et al. 2016) (Table S1; Figure 1D and S1). Juvenile trees were planted in mixed stands in a 3 × 3 m grid, maximizing the distance between individuals of the same species. Three criteria were used to select the tree species for the plantings: i) availability of seeds in the conserved habitat to propagate them, ii) use of the species by the local community, which increased their interest in taking care of the restored areas, and iii) tree species with fleshy fruits to attract frugivores. Proportion of species of each sexual system established in the plantings were similar to what is present in the forest: 12 of the planted species are hermaphrodites, five species are dioecious, and one species is monoecious (Table 1). The number of trees with flowers or fruits (Table S2) and floral visitors were registered in a subset of plots seven years after the restoration intervention. Floral visitors, observed between 9:00 am and 3:00 pm, for one hour in each plot, belonged to 113 morphospecies in five orders of insects, with Hymenoptera being the most frequently registered (72 % of visitors), followed by Diptera (24 %); Coleoptera (2 %), Hemiptera (1 %) and Orthoptera (1 %) were seldom registered (Hernández-Galindo 2016). When compared to disturbed habitat, both exclusions and plantings shown signs of recovery of biodiversity and ecological processes (de la O-Toris et al. 2012, Martínez-Garza et al. 2016, Martínez-Garza et al. 2011, Martínez-Garza et al. 2021, Juan-Baeza et al. 2015, Table S1). The richness and density of sexual systems was calculated using a census of all planted or naturally recruited trees reaching ≥ 2.5 cm of Diameter at Breast Height (DBH); census was performed during 2013-2014 in all habitats (Table 1). By 2014, 163 planted trees reached ≥ 2.5 cm of DBH and were included in the analysis. The sexual system of all tree species was determined by consulting the literature.
Table 1 Family, sexual expressions (Sexual express), and absolute abundance of 46 tree species with DBH ≥ 2.5 cm in disturbed habitat (Dist), 8-year-old restoration intervention (exclusions [Excl] and plantings [Plant]), and conserved habitats (Cons) in the tropical dry forest of Sierra de Huautla, Morelos, central Mexico. Sexual expressions are dioecious (Dio), monoecious (Mono) and hermaphrodite (Herma); references for sexual expressions of each species are shown. The 18 tree species planted are indicated with asterisks ; for them, the number of planted trees reaching DBH ≥ 2.5 cm by 2013 is in parenthesis.
| Family | Species | Sexual express | Abundance of trees | References | |||
|---|---|---|---|---|---|---|---|
| Restored | |||||||
| Dist | Excl | Plant | Cons | ||||
| Anacardiaceae | Amphipterygium adstringens (Schltdl.) Standl. | Dio | 2 | 1 | 0 | 5 | Medina-Lemos 2000 |
| Pseudosmodingium perniciosum (Kunth) Engl. | Dio | 0 | 0 | 0 | 1 | Medina-Lemos & Fonseca 2009 | |
| Spondias purpurea L. | Dio | 1 | 4 | 5 | 2 | Bullock 1994 | |
| Apocynaceae | Plumeria rubra L. | Herma | 1 | 0 | 0 | 1 | Alvarado-Cárdenas 2004 |
| Cascabela thevetioides (Kunth) Lippold | Herma | 0 | 1 | 1 | 2 | Alvarado-Cárdenas 2004 | |
| Bignoniaceae | Astianthus viminalis (Kunth) Baill. | Herma | 2 | 0 | 0 | 0 | Rzedowski & Calderon-Rzedowski 1993 |
| Crescentia alata Kunth* | Herma | 0 | 0 | 0 (2) | 0 | Rzedowski &Calderon-Rzedowski 1993 | |
| Bombacaceae | Ceiba sp.* | Herma | 3 | 8 | 1 (8) | 9 | Pagaza-Calderón & Fernández-Nava 2004 |
| Pseudobombax ellipticum (Kunth) Dugand* | Herma | 0 | 0 | 0 (18) | 0 | Pagaza-Calderón & Fernández-Nava 2004 | |
| Burseraceae | Bursera linanoe (La Llave) Rzed., Calderón & Medina* | Dio | 0 | 0 | 5 (8) | 0 | Medina-Lemos 2008 |
| Bursera bicolor (Willd. ex Schltdl.) Engl. | Dio | 4 | 0 | 2 | 0 | Instituto de Biología 2011 | |
| Bursera bipinnata (DC.) Engl. | Dio | 2 | 0 | 0 | 0 | Medina-Lemos 2008 | |
| Bursera copallifera (DC.) Bullock* | Dio | 12 | 6 | 6 (7) | 11 | Medina-Lemos 2008 | |
| Bursera grandifolia (Schltdl.) Engl.* | Dio | 0 | 2 | 0 (2) | 1 | Medina-Lemos 2008 | |
| Bursera lancifolia (Schltdl.) Engl.* | Dio | 0 | 1 | 2 | 5 | Rzedowski &Guevara-Féfer 1992 | |
| Caricaceae | Jacaratia mexicana A. DC.* | Dio | 0 | 0 | 0 (8) | 0 | Pennington & Sarukhán 2005 |
| Convolvulaceae | Ipomoea sp. | Herma | 30 | 67 | 50 | 58 | Ayestarán-Hernández 2004 |
| Euphorbiaceae | Euphorbia schlechtendalii Boiss. | Mono | 19 | 105 | 15 | 67 | Martínez-Gordillo et al. 2002 |
| Sapium macrocarpum Müll. Arg. | Mono | 12 | 12 | 11 | 5 | Martínez-Gordillo et al. 2002 | |
| Fabaceae | Conzattia multiflora (B.L. Rob.) Standl.* | Herma | 2 | 7 | 4 (3) | 157 | Dorado et al. 2005 |
| Haematoxylum brasiletto H. Karst.* | Herma | 0 | 1 | 0 (13) | 16 | Dorado et al. 2005 | |
| Erythrina americana Mill.* | Herma | 0 | 0 | 0 (9) | 0 | Olvera-Luna et al. 2012 | |
| Eysenhardtia polystachya (Ortega) Sarg. | Herma | 1 | 0 | 4 | 3 | Dorado et al. 2005 | |
| Lonchocarpus caudatus Pittier | Herma | 1 | 0 | 2 | 7 | Dorado et al. 2005 | |
| Acacia cochliacantha Humb. & Bonpl. ex Willd. | Herma | 39 | 98 | 40 | 6 | Rico-Arce & Rodríguez 1998 | |
| Acacia farnesiana (L.) Willd. | Herma | 27 | 22 | 22 | 0 | Rico-Arce & Rodríguez 1998 | |
| Acacia pennatula (Schltdl. & Cham.) Benth. | Herma | 1 | 3 | 0 | 0 | Rico-Arce & Rodríguez 1998 | |
| Enterolobium cyclocarpum (Jacq.) Griseb.* | Herma | 0 | 0 | 1 (1) | 0 | Dorado et al. 2005 | |
| Lysiloma acapulcense (Kunth) Benth. | Herma | 4 | 4 | 1 | 1 | Dorado et al. 2005 | |
| Lysiloma divaricatum (Jacq.) J.F. Macbr.* | Herma | 8 | 12 | 12 (42) | 392 | Dorado et al. 2005 | |
| Leucaena esculenta (DC.) Benth.* | Herma | 0 | 0 | 0 (6) | 0 | Dorado et al. 2005 | |
| Leucaena leucocephala subsp. Glabrata (Rose) Zárate* | Herma | 0 | 0 | 0 (15) | 0 | Pennington & Sarukhán 2005 | |
| Mimosa benthamii J.F. Macbr. | Herma | 20 | 30 | 37 | 4 | Grether et al. 2006 | |
| Mimosa polyantha Benth. | Herma | 0 | 1 | 0 | 3 | Grether et al. 2006 | |
| Fagaceae | Quercus sp. | Mono | 0 | 0 | 1 | 0 | González-Villarreal 1986 |
| Malpighiaceae | Bunchosia canescens (Aiton) DC. | Herma | 1 | 7 | 2 | 2 | León-Velasco 2005 |
| Malpighia mexicana A. Juss.* | Herma | 8 | 3 | 2 (1) | 48 | León-Velasco 2005 | |
| Meliaceae | Trichilia hirta L. | Dio | 2 | 2 | 0 | 0 | Germán-Ramírezm 2005 |
| Swietenia humilis Zucc.* | Mono | 0 | 0 | 0 (4) | 0 | Boshier in White et al. 1999 | |
| Polygonaceae | Ruprechtia fusca Fernald* | Dio | 1 | 15 | 0 (13) | 27 | Solano & Ayala 2008 |
| Rubiaceae | Randia echinocarpa DC. | Dio | 0 | 18 | 5 | 7 | Bye et al. 1991 |
| Sterculiaceae | Guazuma ulmifolia Lam. | Herma | 2 | 3 | 3 | 9 | De León-Ibarra 2005 |
| Tiliaceae | Heliocarpus pallidus Rose | Herma | 3 | 96 | 46 | 5 | Pérez-Calix 2009, Bullock & Solís-Magallanes 1990 |
| Ulmaceae | Celtis caudata Planch. | Herma | 1 | 0 | 1 | 1 | Pérez-Calix & Carranza-González 1999, |
| Verbenaceae | Vitex mollis Kunth* | Herma | 3 | 0 | 4 (2) | 3 | Willmann et al. 2000 |
| Vitex pyramidata B.L. Rob. ex Pringle | Herma | 1 | 1 | 0 | 0 | Willmann et al. 2000 | |
Statistical analyses. Experimental plots are separated from each other by 80-1,600 m, and plots from the same habitat are separated by at least 200 m; we used plots as replicates to compare richness (total number of species per hectare) and tree density (number of trees per hectare) of three sexual systems (dioecious, monoecious and hermaphrodites) in the four habitats. Two-way analysis of variance (ANOVA) was used to analyze richness and tree density by habitat and sexual system, carrying out Tukey post hoc test when significant (Zar 1996). Tree richness and density were log-transformed to fulfil ANOVA assumptions. In the results, figures show log-transformed data, while averages and standard errors were back-transformed for clarity.
To analyze tree composition associated with habitat, we performed four non-metric multidimensional scaling analyses (NMDS; Gauch 1982), one including all sexual systems and one within each sexual system among the four habitats. Matrices of similarity coefficients were calculated using Pearson correlations with tree density at each plot. The number of NMDS dimensions was decided based on the number that yielded the lowest stress value (the lower the stress value, the better the fit) and the more meaningful solution; d-hat stress values are reported (StatSoft 2004). Additionally, we used permutational multivariate analysis of variance (PERMANOVA; Anderson 2001) to test variation in tree composition of sexual system, using tree density among habitats. Dissimilarity was calculated with a Bray-Curtis matrix. Ellipses in the NMDS are based on the PERMANOVA analysis. All analyses were carried out in STATISTICA 7.0 (StatSoft 2004), except for the PERMANOVA analysis, which was carried out in R (R Core Team 2020). The “vegan” library of R was used for this analysis (Oksanen et al. 2018).
Results
In the four habitats 2,042 trees from 46 tree species in 18 families were identified (Table 1). Overall habitats, the highest percentage of species (63 %) and trees (78 %) were hermaphrodites, the lowest percentage of species were monoecious (9 %) and the lowest percentage of trees (10 %) were dioecious. In the plantings were registered the largest number of species (37 species; Table 2) and in the conserved habitat the largest number of trees (852 trees; Table 2).
Table 2 Total number (#) and percentages (%) of species and trees (≥ 2.5 cm DBH) by sexual expression in four habitats in Sierra de Huautla, Morelos, Mexico. Number of planted species and trees are in parenthesis. Percentages are calculated with the total number of species or trees within habitats.
| Habitat | Sexual expressions | Species | Trees | ||
|---|---|---|---|---|---|
| # | % | # | % | ||
| Disturbed | Dioecious | 7 | 24 | 24 | 11 |
| Monoecious | 2 | 7 | 31 | 15 | |
| Hermaphrodites | 20 | 69 | 158 | 74 | |
| Excluded | Dioecious | 8 | 30 | 49 | 9 |
| Monoecious | 2 | 7 | 117 | 22 | |
| Hermaphrodites | 17 | 63 | 364 | 69 | |
| Plantings | Dioecious | 4(5) | 21(28) | 25(38) | 9(23) |
| Monoecious | 3(1) | 16(5) | 27(4) | 9(3) | |
| Hermaphrodites | 12(12) | 63(67) | 233(120) | 82(74) | |
| Conserved | Dioecious | 8 | 28 | 59 | 7 |
| Monoecious | 2 | 7 | 72 | 8 | |
| Hermaphrodites | 19 | 65 | 721 | 85 | |
The analysis of variance (ANOVA) showed that tree richness was statistically similar among the four habitats evaluated (Figure 2A; Table S3a). The average richness of hermaphrodites (47.66 ± 1.1 tree species/ha; mean ± standard error) was three times higher than the richness of dioecious species (14.87 ± 1.1 species/ha) and seven times higher than the richness of monoecious species (7 ± 1.1 species/ha). The ANOVA showed that there were significant differences in richness among the three sexual systems evaluated. The post hoc Tukey test revealed that richness of hermaphrodites was significantly higher than richness of dioecious and monoecious species, and that richness of dioecious species was significantly higher than that of monoecious tree species (Figure 2B; Table S3a). The interaction of habitat and sexual system was not significant (Table S3a).
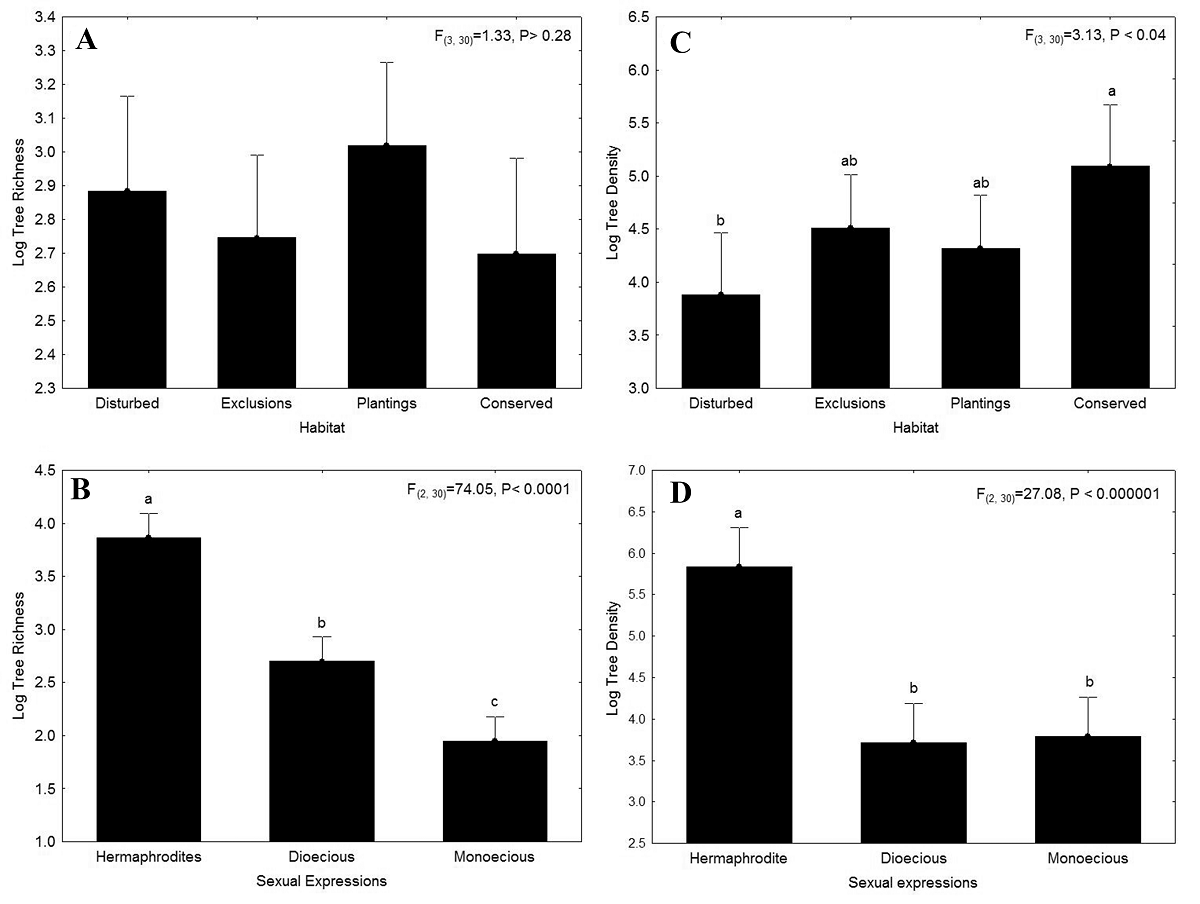
Figure 2 Natural Logarithm of A) tree richness and C) density in four habitats: disturbed, exclusions, plantings, and conserved habitat and, B) richness and D) density of three sexual expressions: dioecious, monoecious and hermaphrodite tree species at Sierra de Huautla, Morelos, Mexico. Error bars show 95 % confidence intervals. Different letters represent significant differences evaluated with a post hoc Tukey test after significant ANOVA results.
Average tree density was three times higher in the conserved (162.59 ± 1.3 trees/ha; mean ± standard error) than in the disturbed habitat (48.34 ± 1.3 trees/ha). Between the two restored habitats, density was higher in the exclusions (90.94 ± 1.3 trees/ha) than in the plantings (74.84 ± 1.3 trees/ha). The ANOVA showed significant differences in tree density among the four habitats evaluated (Table S3b). The post hoc Tukey test revealed that tree density was significantly higher in the conserved than in the disturbed habitat while tree density in exclusions and plantings was intermediate and was not statistically distinguishable from the conserved or disturbed habitats (Figure 2C). Average tree density of hermaphrodites (344 ± 1.3 trees/ha) was eight times higher than the density of dioecious (41 ± 1.2 trees/ha) and monoecious trees (44 ± 1.3 trees/ha). The ANOVA showed significant differences in tree density among the three sexual systems evaluated (Table S3b). The post hoc Tukey test revealed that the density of hermaphrodites was significantly higher than the density of dioecious and monoecious species, while the density of dioecious and monoecious species was statistically similar (Figure 2D). The interaction of habitat and sexual system was not significant (Table S3b).
When all species were included, non-metric multidimensional scaling (NMDS) ordination of the tree species was meaningful with two dimensions (stress = 0.15, Figure 3a). Composition heterogeneity was lowest for the plots in the conserved habitat (green ellipse in Figure 3a), and formed a distinct, but adjacent, group from the plots with plantings (blue ellipse in Figure 3a). Tree composition of exclusions and disturbed habitats overlapped (red and yellow ellipses in Figure 3a). PERMANOVA analysis revealed that tree composition differed significantly among habitats (F (3,13) = 3.21, P < 0.001).
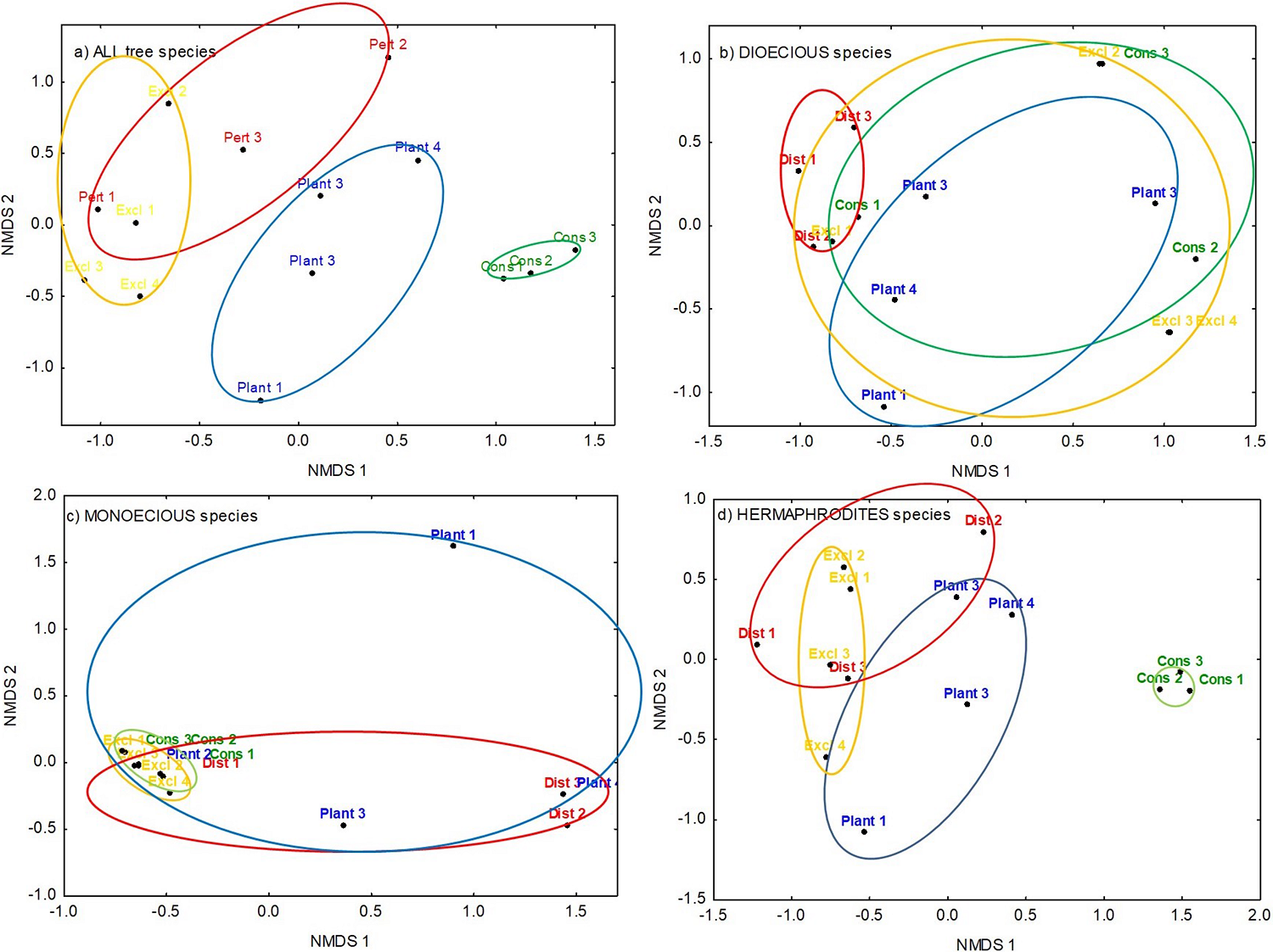
Figure 3 Non-metric multidimensional scaling (NMDS) analysis for a) ALL sexual expressions, b) dioecious species, c) monoecious species, and d) hermaphrodite species, in disturbed habitat (Dist; red), exclusions (Excl; yellow), plantings (Plant; blue), and conserved habitat (Cons; green) at Sierra de Huautla, Morelos, central Mexico. The ellipses, based on PERMANOVA results, indicate highly similar composition of tree species within habitats according to color.
When considering dioecious trees only, NMDS ordination of the tree species was meaningful with two dimensions (stress = 0.12; Figure 3b); composition heterogeneity was lowest within the disturbed habitat (red ellipse in Figure 3b) and higher for the other three habitats, whose composition overlapped (Figure 3b). PERMANOVA analysis revealed that tree composition of dioecious species was statistically similar among habitats (F (3,13) = 1.69, P > 0.08).
When considering monoecious species only, NMDS ordination of the tree species was meaningful with two dimensions (stress = 0.005; Figure 3c); composition heterogeneity was lowest within the conserved habitat (green ellipse in Figure 3c) and the excluded habitat (yellow ellipse in Figure 3c) while the highest composition heterogeneity was registered for plantings (blue ellipse in Figure 3c). PERMANOVA analysis revealed that tree composition of monoecious species was statistically similar among habitats (F (3,13) = 1.51, P > 0.18).
When considering hermaphrodite species only, NMDS ordination of the tree species was meaningful with two dimensions (stress = 0.09; Figure 3d); the lowest composition heterogeneity was registered within the conserved plots, and they constituted a different group (green circle in Figure 3d); composition heterogeneity for the other three habitats was similar and overlapped (Figure 3d). PERMANOVA analysis revealed that tree composition of hermaphrodite species differed significantly among habitats (F (3,13) = 3.84, P < 0.001).
Discussion
Contrary to our prediction, that obligate outcrossing tree species (dioecious and monoecious) were equally represented at all habitats probably due to their use value. Also, composition of dioecious tree species was as heterogeneous in restored areas as in the conserved forest, thanks to plantings. Finally, there was a higher richness and density of hermaphrodites at all habitats, as expected, but their composition differed the most between disturbed and conserved habitats with restored habitats falling in between.
Dioecious species. Contrary to our expectations, the richness, density, and tree composition of dioecious species were similar in conserved, restored, and disturbed habitats. In dry forests of Venezuela (Jaimes & Ramírez 1999) and Costa Rica (Hilje et al. 2015), dioecious tree species were more frequently found in continuous forest and in late successional environments, contrary to our results. Here, we argue that categories of use of these species and the relation of use value (i.e., average of uses reported for each species by the number of informants interviewed; Maldonado et al. 2013) with the ecological importance of species (e.g., abundance) may explain these results: The 13 dioecious tree species recorded at this permanent agricultural landscape have high use value for the local people (Table S4). According to categories of use of Maldonado et al. (2013), the most frequent use for dioecious tree species is as living fence and in second place, as medicine. Living fences are rows of stakes harvested from tree branches used to separate properties or areas within properties in agricultural landscapes in Latin America; barbed wire is attached directly to the live trees, which function as fenceposts (Budowski & Russo 1993). Also, living fences have been recognized as corridors for fauna and to favor interactions such as pollination and seed dispersal (Tewksbury et al. 2002, Harvey et al. 2006, Griscom et al. 2007, Pizo & dos Santos 2011). Further, dioecious species have up to six other uses, for example as food, ornamental, and ritual (Table S4); all these additional uses are considered as non-timber forest products and given that trees are not cut, the impact in the populations is lower (see Blancas-Vázquez et al. 2017). Therefore, these categories of use favored the presence of dioecious species in these permanent agricultural landscapes (Cruz-León et al. 2006, Mena-Jimenez 2018). In other dry forests, for example, in Chamela, Mexico, dioecious tree species have not been recorded at all in man-made pastures (Quesada et al. 2011), possibly because native tree species are less frequently used there (Burgos & Maass 2004). Also, the dioecious Bursera copallifera (DC.) Bullock and B. bipinnata (DC.) Engl. are reported as the most common species used to extract resins in the Balsas Basin in Mexico (Linares & Bye 2008), and these species are promoted and protected in agricultural areas (Abad-Fitz et al. 2020). Accordingly, in this study, tree abundance of B. copallifera was as high in conserved as in disturbed areas (Table 1). Finally, a relationship of the use value with the ecological value of species (i.e., plant density or frequency) is expected in ecosystems with presence of indigenous or mestizo populations (e.g., Galeano 2000). For dioecious species, we found a high positive correlation of the use value reported by Maldonado et al. (2013) and the abundance of trees in the disturbed habitat mostly due to B. copallifera (r = 0.80, P < 0.001; Figure S2); supporting the hypothesis that these species have been protected in the agricultural landscapes due to their use value. Finally, tree composition of dioecious species overlapped among all habitats while composition heterogeneity (i.e., size of ellipses in NMDS figures) was higher and comparable in restored and conserved habitats because two of the planted species (i.e., Bursera grandifolia (Schltdl.) Engl. and Ruprechtia fusca Fernald) were not frequent in the disturbed habitat (Table 1). Dioecious species have been favored in this permanent agricultural landscape due to their use value while composition heterogeneity increased in restored habitats due to planted species.
Monoecious species. Contrary to our expectations, the richness, density, and tree composition of monoecious species were similar at all habitats. Overall habitats, only four monoecious tree species were recorded (see Table 1): the precious tropical wood Swietenia humilis Zucc. (genuine mahogany), and one Quercus sp. are used for construction and firewood; given that they only have two uses and are seldom mentioned by informants, their use value is low (Maldonado et al. 2013). Contrary to what happen with dioecious species, these two categories of use, construction and firewood, resulted in the elimination of complete populations; specifically, the extraction of the genuine mahogany for timber started five centuries ago (Rodan et al. 1992), and this species was not registered in the conserved habitat neither in > 65-year-old forest fragments measured in other study (Beltrán-Rodríguez et al. 2018). In contrast, Euphorbia schlechtendalii Boiss. which is used for living fences and has a higher use value, was the most abundant species recorded in the four habitats and 51 % of the trees were registered in the excluded habitat (Table 1). This species recruits successfully early in succession; for example, in the study site, recruitment census in first 5 years of exclusion revealed that E. schlechtendalii was one of the most abundant species (Alba-García 2011, Martínez-Pérez 2014). This successful recruitment at all excluded plots may explain the low composition heterogeneity in that habitat. Accordingly, E. schlechtendalii was recorded as the second most abundant species in the dry forest of Nizanda, Oaxaca, Mexico (Gallardo-Cruz et al. 2005) and the most important species in the understory in the dry forest of Morelos, Mexico (Beltrán-Rodríguez et al. 2018). Finally, we found a high positive correlation of the use value reported by Maldonado et al. (2013) and the abundance of trees in the excluded (r = 0.99, P < 0.001; Figure S3) and conserved habitat (r = 0.99, P < 0.001) mostly due to E. schlechtendalii. Monoecious species once used for construction and firewood have been eliminated from this permanent agricultural landscape; composition heterogeneity was highest in the plantings due to the introduction of S. humilis and it was lowest in exclusions due to the high natural recruitment of E. schlechtendalii.
Hermaphrodites. Richness and density of hermaphrodites were similar among all habitats, as expected; also, tree composition was unique in the conserved habitat, contrary to our predictions. Given the predominance of hermaphroditism in plants everywhere (de Jong et al. 2008) and particularly in tropical floras (reviewed in Machado et al. 2006), a higher representation of hermaphrodites is expected at all habitats. For example, in an old secondary dry forest of Venezuela, 82 % of tree species recorded were hermaphrodites (Jaimes & Ramírez 1999). Also, in the mature dry forest of Chamela, Mexico, more than half of the species recorded were hermaphrodites (54 %; Bullock 1985). This representation is explained by the dominance of the Fabaceae family in dry forests; this family is composed of hermaphrodite species (Banda-R et al. 2016), although some species may be in a transitional stage to monoecy (andromonoecious; Ortiz et al. 2003, Nair et al. 2004, Casimiro-Soriguer et al. 2013, Zimmerman et al. 2013). On the other hand, tree composition of hermaphrodites differed greatly between conserved and the other three habitats evaluated, probably due to limitations in dispersal and establishment: in this landscape, 66 % of the hermaphrodite species registered as ≥ 2.5 cm DBH were also found in the seed rain of early successional environments (Martínez-Garza et al. 2011), therefore, they seem to have low dispersal limitation (failure of seeds to reach a site; Terborgh et al. 2011). However, only 23 % of the species (Ceiba sp., Ipomoea sp. A. cochliacantha, A. farnesiana, Lysiloma divaricatum, Mimosa benthamii and Heliocarpus pallidus Rose) were found recruiting in early successional environments (in the first 4 years; Alba-García 2011) which suggest a strong establishment limitation (failure of seedlings to survive and grow; Muller-Landau et al. 2002) for the other 70 % of the species. Also, according to categories of use of Maldonado et al. (2013), the most frequent use of hermaphrodite species is firewood and in second place, construction. When including all hermaphrodite species, we did not find a correlation of the use value reported by Maldonado et al. (2013) and the abundance of trees in the disturbed habitat (r = -0.16, P > 0.4; Figure S4). The most abundant hermaphrodite species in the disturbed habitat (Acacia cochliacantha, A. farnesiana, Ipomoea sp. and Mimosa benthamii) are early successional tree species used only for firewood, so they have a low use value while other two early successional species also used for firewood but less abundant in the disturbed habitat have high use value given additional uses: Guazuma ulmifolia Lam. is also used for construction, fodder, and crafts while Leucaena esculenta (DC.) Benth. is also used for fodder (Table S1, Figure S4). Excluding two early successional trees seldom registered in the disturbed habitat and with low use value (Acacia pennatula (Schltdl. & Cham.) Benth. and Mimosa polyantha Benth.), there is a negative correlation of use value and abundance in disturbed habitat for six early successional hermaphrodite species (Figure S4). Finally, detailed information about mating system is not available for all tree species, but reviews including > 700 population of > 100 species showed that ca. 40 % of hermaphrodite species may have obligate outcrossing, and they may have at least one population with mixing mating system (Whitehead et al. 2018), so they may also have reproductive limitations in fragmented landscapes. In conclusion, distribution of hermaphrodite species in this permanent agricultural landscape might be explained by successional status of species: even though more than half of the hermaphrodite species recorded in this study are dispersing in this landscape, only few early successional species are establishing there, and their main use is firewood.
Implications for management. The United Nations declared the “Decade for Ecosystem Restoration” (2021-2030; www.decadeonrestoration.org) which main aim is to reverse current forest loss. One objective towards that aim is to increase connectivity in fragmented habitats to favor the movement of plants and animals. In permanent agricultural landscapes, the concept of Forest Landscape Restoration (FLR) is particularly useful because it aims is to provide benefits to both people and biodiversity by balancing ecological, social, and economic priorities (Stanturf et al. 2017). FLR favors the establishment of productive landscape units while protecting areas for biodiversity maintenance. Related to that concept, in this study, overall sexual systems, tree richness and density was already similar at conserved habitats and restored ones after eight years within permanent agricultural landscapes. It is expected that restoration actions further accelerate successional times and therefore, our results agree with reports in Bolivia, where in land abandoned after slash and burn agriculture, tree richness recovered rapidly (< 10 years) while density reached maximal values at five years and then decreased due to an increase in basal area, to values recorded in the conserved forest (Kennard 2002). Also, in Oaxaca, Mexico, tree richness and density in successional areas increased quickly during the first years of succession; richness decreased when trees became larger while density reached the values of the mature forest after ca. 10 years (Lebrija-Trejos et al. 2010). Surprisingly, tree richness in the disturbed habitat was also similar to the one in restored and conserved habitats, probably due to the use value of tree species. For example, in the dry forest of Panama (Griscom et al. 2009), Costa Rica (Gerhardt 1993) and Chamela, Mexico (Burgos & Maass 2004), cattle ranching activities are carried out in pastures seeded with exotic grasses and most of the original trees are eliminated. At our study site, secondary forest is used for cattle ranching activities during the wet season; useful trees are not eliminated to sow exotic grasses, rather cattle consume native and ruderal grasses and tree seedlings (de la O-Toris et al. 2012). Also, in this site, it was found that even when disturbance increased richness and abundance of cactus, size of individuals was affected, with important implication for fauna feeding in adults individuals (Arias-Medellín et al. 2014). Our results suggest that tree richness has not been affected by cattle ranching activities while density and composition was affected, which may have consequences for plant-animal interaction, as pollination and seed dispersal.
Restoration plantings are established with different composition. Selection of tree species may be based on growth rates (Lugo 1997, Butler et al. 2008), attraction of frugivorous animals (de la Peña-Domene et al. 2013, 2014) or availability of species at nurseries (Montoya-Pfeiffer et al. 2018). In this study, criteria to select species for plantings immersed in agricultural landscapes included availability of seeds, use of the species by local people and provision of food to animals: eight tree species with fleshy fruits that attract frugivorous animals were chosen (Martínez-Garza et al. 2016); relevant to this discussion, in a study of the flora of Puerto Rico (Flores & Schemske 1984) and in Chamela. Mexico (Quesada et al. 2011), it was found that more than 60 % of dioecious tree species may have fleshy fruits. Therefore, for restoration plantings, some trees may provide food for frugivorous animals and pollinators and help to maintain sexual systems sensitive to fragmentation. Tree composition in restoration plantings may favor the movement of genetic diversity in the agricultural landscape by attracting pollinators that also visit conserved and productive areas, therefore, productive units will benefit from natural and restored areas (Meli et al. 2019). Further, diversifying tree species and genotypes in restoration plantings to extend phenological activity, may buffer negative effects of climate change in plant-animal interactions (Olliff-Yang et al. 2020). For example, in this study, in August, only one species was found with flowers in the agricultural landscape (Mimosa benthamii; Hernández-Galindo 2016). Also, restoration projects should be designed including systemic mosaics of contrasting treatments (systemic experimental restoration sensuHowe & Martínez-Garza 2014) containing all sexual systems. Sexual system should be a criterion to choose tree species for restoration plantings, then not only forest structure will be recovered but also ecosystem function.
Self-incompatible species will show complete dependence on pollinators and other reproductive individuals for reproduction (Quesada et al. 2011). Sex ratio may be significantly impacted by fragmentation (Barrett & Hough 2013) and by use value, because male and female trees may have different uses; for example, female trees of dioecious species with fleshy fruits consumed by humans will be preferred over males (Seethapathy et al. 2018). Also, in our study site, female trees of Amphipterygium adstringens (Schltdl.) Standl., which is debarked for medicinal use, showed higher bark regeneration compared to males (Beltrán-Rodríguez et al. 2021). Then, for these species, the frequency and spatial distribution of female and male trees in the permanent agricultural landscape and their categories of use will be important considerations which remains to be evaluated. In this study, for the subsample evaluated, the highest number of trees with flowers within one habitat were found for the monoecious E. schlechtendali (14 trees in the excluded habitat) followed by the hermaphrodite A. cochliacantha (11 trees in the excluded habitat; Table S1) while for most dioecious species, only one tree was found with flowers within one month (Hernández-Galindo 2016). These results are related to the successful recruitment of E. schlechtendali and A. cochliacantha in early successional environments, as discussed above. Then, even when tree richness and density of sexual systems was similar at all habitats, function may not be yet recovered. Future studies should evaluate recruitment potential of tree species with different sexual systems, pollinator assemblages and its health (for example, González-Tokman & Martínez-Garza 2015) in permanent agricultural landscapes to guide conservation and restoration actions (see for example, Martins & Antonini 2016). Specifically, dioecious (A. adstringens debarked for medicine use; Beltrán-Rodríguez et al. 2021) and monoecious tree species (S. humilis extracted for timber), which populations have been affected due to their use and also, late-successional hermaphrodite species missing in permanent agricultural landscapes (i.e., Haematoxylum brasiletto H. Karst., Table 1) should be selected for restoration plantings.
Supplementary material
Supplemental data for this article can be accessed here: https://doi.org/10.17129/botsci.2971
Supplementary material Figures S1-S4 and Table S1-S4.

