











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Las montañas han sido consideradas como áreas que concentran biodiversidad (Körner 2000, Körner 2004, Mutke & Barthlott 2005, Körner & Spehn 2019, Rahbek et al. 2019). Los factores que generan esta concentración se deben a la presencia de cinturones termales (Körner et al. 2011), heterogeneidad topográfica (Ricklefs 1977, Richerson & Lum 1980, Burnett et al. 1998, Nichols et al. 1998, Badgley et al. 2017), heterogeneidad climática (Suárez-Mota et al. 2017), microclimas (Kutzbach et al. 1993, Raupach & Finnigan 1997, Lembrechts et al. 2019), diferencias en radiación solar (Bennie et al. 2008), geodiversidad (During & Willems 1984, Brooks 1987, Gray 2004, Hjort et al. 2015, Moreno et al. 2020, Van Santen & Linder 2020) y diferencias en elevación y pendiente (Zhirnova et al. 2020). También la geología, la heterogeneidad del relieve y la mezcla de tipos de suelo han sido correlacionados con la diversidad (Hoorn et al. 2013, Antonelli 2015, Payne et al. 2017, Zizka & Antonelli et al. 2018, Körner & Spehn 2019, Rahbek et al. 2019, Silveira et al. 2019, Perrigo et al. 2020). Por ejemplo, ciertos procesos como la disolución de la roca sedimentaria por acción del intemperismo, han generado paisajes kársticos y sustratos compuestos con un alto contenido de carbonatos de calcio como calizas, lutitas, areniscas y yesos, entre otros, que hacen heterogéneos los hábitats (Mason 1946, Clements et al. 2006, Li et al. 2013, Feng et al. 2020), factor importante para muchas especies microendémicas de plantas (Meyer et al. 1992, Sosa & De-Nova 2012, Salinas-Rodríguez et al. 2017). En este contexto, las montañas, su posición, su extensión y su historia geológica (Hoorn et al. 2013, Antonelli 2015, Zizka & Antonelli et al. 2018, Hobohm et al. 2019, Rahbek et al. 2019, Silveira et al. 2019, Perrigo et al. 2020) han tenido un impacto directo en la evolución y distribución de la biota mexicana y la diversidad de plantas vasculares podría correlacionarse también a rasgos fisiográficos (Harrison et al. 2004, Kruckeberg 2004, Clements et al. 2006, Salinas-Rodríguez et al. 2017, Rahbek et al. 2019).
Existen varias propuestas de delimitación para la Sierra Madre Oriental (SMOr) basadas en rasgos fisiográficos, geológicos, biológicos etc. (Rzedowski 1978, Casas-Andreu & Reyna-Trujillo 1990, Cervantes-Zamora et al. 1990, Ferrusquía-Villafranca 1990, Ramírez & Castro 1990, CONABIO 2008, Espinosa et al. 2004, Morrone 2005, Morrone et al. 2017). Sin embargo, presentan diferencias y aún hoy los límites exactos y sus posibles subdivisiones siguen siendo imprecisas y suscitan discusiones (Lugo-Hubp 1990). En este trabajo se ha decidido usar como polígono de estudio la Provincia Fisiográfica de la SMOr que de forma general es un conjunto de cadenas montañosas ubicadas en el noreste del país, producto de orogenias ocurridas desde el periodo Cretácico (Eguiluz-De Antuñano et al. 2000) que culminaron en un complejo arreglo geomorfológico compuesto de rocas de origen sedimentario en forma de pliegues surcadas por llanuras, valles y lomeríos (Álvarez 1961), éstas se agrupan por la edad e historia de los sustratos, mucho más antiguos que otras cadenas de montañas de México.
La delimitación de esta Provincia abarca una extensión de alrededor de 22,015,151 hectáreas o el 11 % de la superficie continental del país y fue propuesta por Cervantes-Zamora et al. (1990) y retomada por el INEGI (2001a, b). Esta delimitación se refiere a una visión general de las formas del relieve que integran conjuntos paisajísticos sobre áreas muy extensas, los cuales se unificaron considerando las topoformas, la geología, la climatología, la hidrología y la edafología que derivó en unidades relativamente homogéneas llamadas Provincias Fisiográficas (INEGI 2001a, b, d). Para unificar las Provincias Fisiográficas de México se hizo un análisis integral de mapas topográficos, refiriéndose a un sistema de topoformas que incluyó para la SMOr en orden de abundancia decreciente: a) sierras, que se refiere a las montañas, ya fueran altas escarpadas, plegadas, complejas, de laderas convexas etc. (54 %), b) bajadas, que se refiere a las laderas o pie de monte de la topoforma sierra (20 %), c) llanuras, que son áreas planas intermontanas o rodeadas de sierras (17 %), d) valles, que se refiere a zonas planas estrechas y/o alargadas intramontanas (3 %), e) lomeríos, que son zonas onduladas entre las sierras (2 %), f) mesetas, que son áreas planas en la parte alta de las sierras, y g) cañones, que se refiere a áreas angostas de paredes verticales intramontanas que han sido esculpidas por la acción erosiva del agua (1 %) (INEGI 2001c). La Provincia Fisiográfica de la SMOr quedó conformada por ocho Subprovincias Fisiográficas que de noroeste a sureste son: a) Serranía del Burro, b) Sierras y Llanuras Coahuilenses, c) Sierra de La Paila, d) Pliegues Saltillo-Parras, e) Sierras Transversales, f) Gran Sierra Plegada, g) Sierras y Llanuras Occidentales, y h) Carso Huasteco (INEGI 2001d). A diferencia de otras delimitaciones de la SMOr que se basan en lo que correspondería a la Subprovincia del Carso Huasteco y la Gran Sierra Plegada, la delimitación de Provincia Fisiográfica de la SMOr (Cervantes-Zamora et al. 1990, INEGI a, b) incluye sierras calcáreas transicionales del norte de México en los estados de Coahuila, Zacatecas, Durango, Nuevo León y San Luis Potosí, así como valles intermontanos y lomeríos con matorrales, en donde se traslapan elementos asociados al Desierto Chihuahuense (Rzedowski 1973, Villaseñor et al. 2020) que se asocian a sitios fisiográficamente considerados como sierra, pero que ecológica y biogeográficamente no lo son.
Sin embargo, desde su identidad biogeográfica, Espinosa et al. (2004) utilizan el polígono Fisiográfico de la SMOr y con base en la distribución de conjuntos importantes de endemismos de flora y fauna, recalcan una evidente división en dos subprovincias: meridional y septentrional, que se divide con el cauce del río Moctezuma (Pánuco medio). La Provincia Meridional se divide en los distritos de Sierra Gorda y Huayacocotla, la cual está relacionada con Mesoamérica y las Antillas, mientras que la Provincia Septentrional se divide en los distritos de Parras-Saltillo y Potosí, relacionados con la Sierra Madre Occidental y las zonas áridas del sureste de Estados Unidos, estos resultados también se sustentaron en los de Luna-Vega et al. (2000) quienes mencionan que los cañones del río Pánuco de frente a la vertiente oriental de la SMOr, son el límite norte de distribución de varias especies de flora tropical como Ageratina ligustrina, Aporocactus flageliformis, Campyloneurum angustifolium, Clethra mexicana, Fleischmannia pycnocephala, Oreopanax xalapensis, Palicourea padifolia, Smilax mollis y Solanum appendiculatum y que incluso marca el límite norte de organismos vágiles como mariposas, cuya distribución está desde Centroamérica hacia el norte, pero se interrumpe en los cañones del río Pánuco, de la misma manera que con especies como Quercus rysophila que se distribuyen hacia el norte de este acotamiento.
En cuanto a diversidad, Luna-Vega et al. (2004) mencionan que un área similar a la Provincia Fisiográfica de la SMOr alberga más de 2,500 especies de plantas vasculares nativas, Mittermeier (2004) propone que el Hotspot de Bosques Madrenses de Pino y Encino, alberga más de 5,300 especies de las cuales 3,975 (75 %) son endémicas. También ha sido considerada una región rica en endemismos de plantas insectívoras del género Pinguicula (Zamudio 2005), tejocotes (Crataegus) (Phipps 1997) y cactáceas, e incluso se plantea la hipótesis de que la tribu Cacteae pudo haberse originado en la SMOr y posteriormente dispersarse a lo que hoy es el Altiplano o la Meseta Mexicana durante el Mioceno - Plioceno (Vázquez-Sánchez et al. 2013, Guerrero et al. 2019, Villaseñor et al. 2020), además se considera a la SMOr junto con el Altiplano Mexicano (Hernández & Bárcenas 1995, Santa Anna-Del Conde et al. 2009) como grandes centros de diversificación.
Otras investigaciones botánicas que toman a la SMOr como un polígono de estudio incluyen en orden cronológico: la composición florística y estructura de los bosques mesófilos de montaña (Rzedowski 1996, Luna-Vega et al. 2001), las especies de gimnospermas (Contreras-Medina et al. 2004), las plantas vasculares acuáticas (Bonilla-Barbosa 2004), un análisis biogeográfico de sus cactáceas y su representatividad en las áreas de conservación (Santa Anna-Del Conde et al. 2009), la biogeografía de Hunnemannia fumarifolia (Ruíz-Sánchez et al. 2012), la estructura de los piñonares de Pinus pinceana (Villarreal-Quintanilla et al. 2009), las poblaciones de Taxus globosa (García-Aranda et al. 2012), la biogeografía de diversas especies de helechos (Sanginés-Franco et al. 2011), los árboles silvestres endémicos (Rzedowski 2015a), la flora vascular de distribución restringida (Rzedowski 2015b, Salinas-Rodríguez et al. 2017), los dominios climáticos y su relación con la diversidad florística (Suárez-Mota et al. 2017) y la efectividad de sus áreas naturales protegidas en la conservación de sus endemismos (Salinas-Rodríguez et al. 2018).
El objetivo de la presente investigación fue elaborar el inventario florístico de esta Provincia Fisiográfica, así como una actualización de sus endemismos y las afinidades geográficas de sus géneros.
Materiales y métodos
Área de estudio. El polígono utilizado en esta investigación corresponde al mapa de las Provincias y Subprovincias Fisiográficas de México de Cervantes-Zamora et al. (1990) e INEGI (2001a, b, d) (Figura 1), el cual se basa en la dominancia del sustrato geológico que corresponde al periodo Cretácico y pendientes mayores a 30 ° C de inclinación y una extensión de 22,015,151 hectáreas. Su altura promedio es de 1,313 m snm, su altura mínima es de 119 m snm (vertiente oriental de la Sierra del Abra Tanchipa en San Luis Potosí) y su altura máxima es de 3,709 m snm (cerro del Potosí en Nuevo León) (INEGI 2017a).

Figura 1 Mapa de localización de la Sierra Madre Oriental. 1. Provincia Fisiográfica de la Sierra Madre Oriental (Cervantes-Zamora et al. 1990, INEGI 2001a, b). 2. Subprovincias Fisiográficas de la Sierra Madre Oriental: A) Serranía del Burro, B) Sierras y Llanuras Coahuilenses, C) Sierra de La Paila, D) Pliegues Saltillo-Parras, E) Sierras Transversales, F) Gran Sierra Plegada, G) Sierras y Llanuras Occidentales, H) Carso Huasteco (INEGI 2001d).
Su clima es complejo ya que posee la mayoría de los tipos climáticos de México que pueden agruparse en cuatro zonas térmicas: a) la cálida con temperaturas de 22 a 26 ° C que se extienden en la vertiente oriental de la sierra y que reciben la influencia directa de los vientos alisios, b) la semi cálida de 18 a 22 ° C que es la región más amplia y se ubica en la vertiente occidental de la sierra, donde predominan las zonas áridas, un máximo de temperatura, altas subtropicales y un régimen pluviométrico muy variable, pero que por lo general las precipitaciones son notablemente menores que en la franja anterior, c) la templada de 12 a 18 ° C que se distribuye de manera fragmentada en las regiones elevadas de las montañas y d) la semi fría de 5 a 12 ° C, únicamente en las cimas de las montañas más altas donde la principal característica es la predominancia de temperaturas bajas y presencia recurrente de heladas (Hernández & Carrasco 2004).
Las unidades de vegetación propuestas en este trabajo se agrupan de acuerdo con la clasificación de Rzedowski (1978) 7y la capa de uso de suelo y vegetación del INEGI (2017b) Serie VI (Figura 2, Tabla 1).
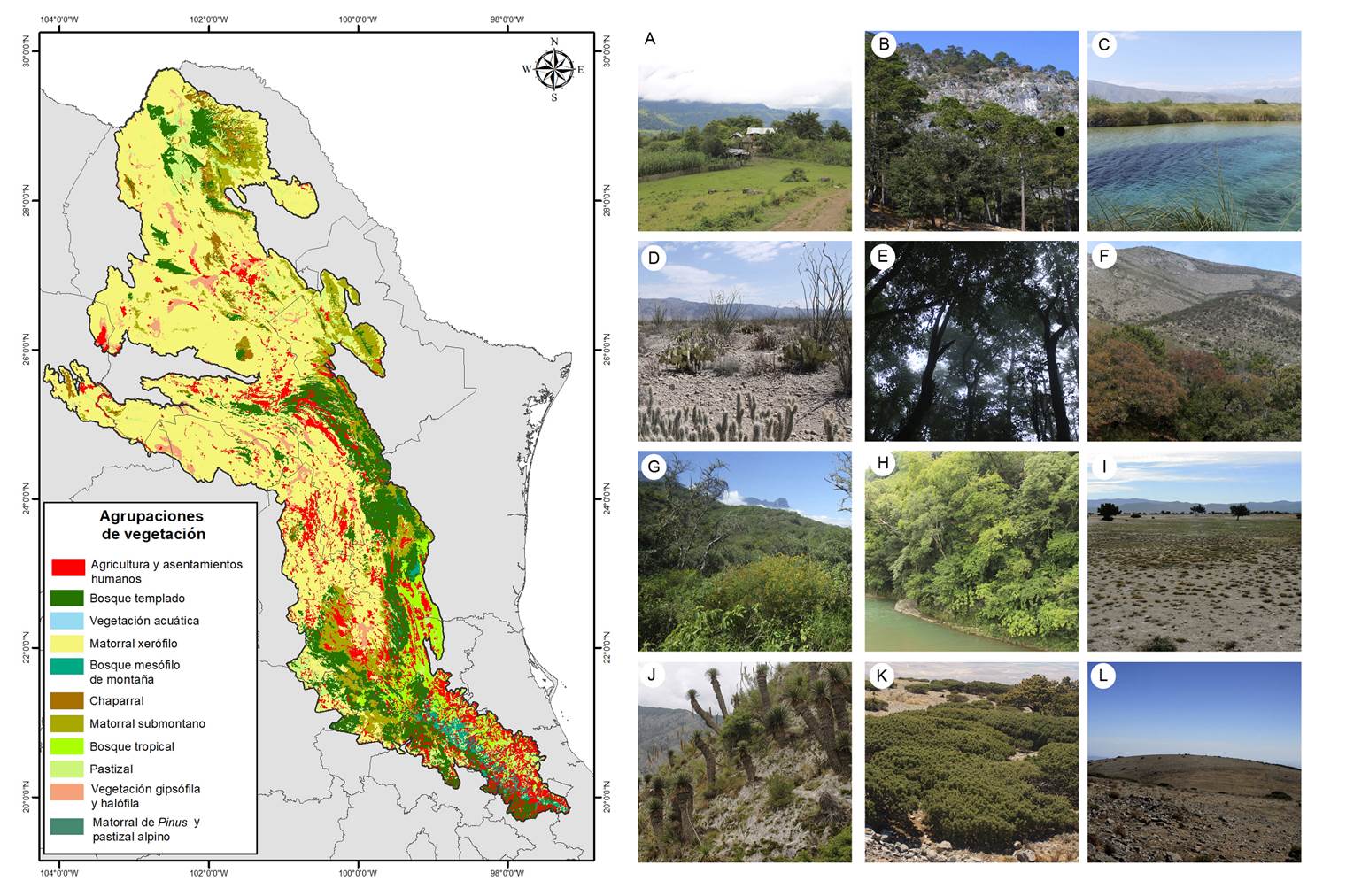
Figura 2 Agrupaciones de vegetación de la Sierra Madre Oriental basadas en Rzedowski (1978) e INEGI (2017b): A) agricultura y asentamientos humanos, B) bosque templado, C) vegetación acuática, D) matorral xerófilo, E) bosque mesófilo de montaña, F) chaparral, G) matorral submontano, H) bosque tropical, I) pastizal, J) vegetación gipsófila y halófila, K) matorral de Pinus y l) pastizal alpino.
Tabla 1 Correspondencia en las agrupaciones de la vegetación de la SMOr. Se simplifican los tipos de vegetación en agrupaciones, basándonos en Rzedowski (1978) e INEGI (2017b). Serie VI
| Agrupación usada en esta investigación | Rzedowski (1978) | INEGI Serie VI (2017b) |
|---|---|---|
| Bosque tropical | bosque tropical perennifolio | selva alta perennifolia, selva alta subperennifolia, selva mediana perennifolia, selva baja perennifolia |
| bosque subcaducifolio, bosque tropical caducifolio, bosque espinoso | selva mediana subcaducifolia, selva mediana caducifolia, selva baja subcaducifolia, selva baja caducifolia, selva baja espinosa subperennifolia, selva baja espinosa caducifolia. | |
| Bosque mesófilo de montaña | bosque mesófilo de montaña | bosque mesófilo de montaña |
| Bosque templado | bosque de Quercus, bosque de coníferas | bosque de pino, bosque de encino, bosque de pino-encino, bosque de encino-pino, bosque de oyamel, bosque de táscate, bosque de cedro, bosque de ayarín. |
| Matorral de Pinus y pastizal alpino | Matorral de Pinus | matorral de coníferas |
| pastizal | pradera de alta montaña | |
| Pastizal | pastizal | pastizal natural |
| Matorral xerófilo | matorral xerófilo | matorral crasicaule, matorral desértico rosetófilo, matorral desértico micrófilo, matorral subtropical, mezquital |
| Chaparral | matorral xerófilo | chaparral |
| Matorral submontano | matorral xerófilo | matorral submontano |
| Vegetación acuática | vegetación acuática y subacuática | selva de galería, bosque de galería, vegetación de galería |
| Vegetación gipsófila y halófila | matorral xerófilo | vegetación halófila, vegetación gipsófila |
| pastizal | pastizal halófilo |
Elaboración del inventario. Se consideraron las especies de plantas vasculares nativas, endémicas de México y endémicas de la SMOr. Se desarrolló una base de datos a partir de la búsqueda de información digital contenida en herbarios (AAU, ANSM, ARIZ, ASU, BH, CFNL, IBUG, MO, MEXU, NY, QMEX, UAMIZ, TEX) desde las bases de datos en línea, principalmente de Tropicos y Global Biodiversity Information Facility, así como artículos científicos, tesis, listados florísticos regionales, monografías y tratados taxonómicos. La nomenclatura se corroboró con ayuda del International Plant Name Index y Taxonomic Name Resolution Service v4.0. Posteriormente pasó por la revisión de cada grupo con los especialistas y se enlistaron con base en la clasificación de licofitas y helechos de Christenhusz et al. (2011) y angiospermas de APG IV (Chase et al. 2016).
Al anterior inventario también se añadió la afinidad geográfica actual a la que se relacionan los géneros en a) boreal: elementos de afinidades neárticas y paleárticas (holárticas), b) endémico mexicano: con elementos mexicanos, c) tropical: elementos de afinidades neotropicales, paleotropicales y pantropicales, d) cosmopolita: con elementos que incluyen una amplia distribución en todo el mundo y e) géneros con afinidades geográficas de las zonas áridas de Sudamérica, África y Australia.
Resultados
Riqueza taxonómica. Los resultados obtenidos sirvieron para elaborar un listado con 6,981 especies nativas de México (Tabla S1), con nombres aceptados según IPNI, pertenecientes a 225 familias y 1,521 géneros (Tabla 2). Del total, 543 tienen categorías infra específicas, 214 son subespecies y 329 variedades. Además se registraron nueve híbridos.
Tabla 2 Riqueza de especies de plantas vasculares nativas por grupo taxonómico. Este listado resulta de las especies nativas de México que tienen parte de su distribución dentro de los límites de la SMOr.
| Grupo taxonómico | Familias | Géneros | Especies |
|---|---|---|---|
| Helechos y afines | 30 | 77 | 356 |
| Gymnospermas | 6 | 13 | 62 |
| Magnólidas | 9 | 22 | 118 |
| Monocotiledóneas | 30 | 252 | 1,061 |
| Dicotiledóneas | 150 | 1157 | 5,384 |
| Total | 225 | 1,521 | 6,981 |
Las familias más ricas fueron Asteraceae (1,000 especies), Fabaceae (495 especies), Cactaceae (354), Poaceae (328), Orchidaceae (189), Euphorbiaceae (178), Lamiaceae (176), Solanaceae (174), Malvaceae (160), Rubiaceae (136), Cyperaceae (131), Asparagaceae (117), Acanthaceae (107), Rosaceae (101) y Crassulaceae (101), en la Tabla 3 se muestran más. Las primeras tres familias representarían una cuarta parte de toda la flora de la SMOr con 26 %.
Tabla 3 Riqueza por familia. Se muestran las 30 familias más ricas en especies de plantas vasculares nativas de la SMOr.
| Familia | Especies | Familia | Especies | Familia | Especies |
|---|---|---|---|---|---|
| Asteraceae | 1,000 | Cyperaceae | 131 | Verbenaceae | 86 |
| Fabaceae | 495 | Asparagaceae | 117 | Fagaceae | 85 |
| Cactaceae | 365 | Acanthaceae | 107 | Plantaginaceae | 78 |
| Poaceae | 328 | Rosaceae | 101 | Bromeliaceae | 74 |
| Orchidaceae | 189 | Crassulaceae | 101 | Apiaceae | 61 |
| Euphorbiaceae | 178 | Convolvulaceae | 99 | Polypodiaceae | 61 |
| Lamiaceae | 176 | Pteridaceae | 98 | Caryophyllaceae | 60 |
| Solanaceae | 174 | Apocynaceae | 90 | Amaranthaceae | 60 |
| Malvaceae | 160 | Boraginaceae | 90 | Nyctaginaceae | 53 |
| Rubiaceae | 136 | Brassicaceae | 90 | Orobanchaceae | 52 |
Los géneros con mayor número de especies son Euphorbia (93 especies), Quercus (84), Mammillaria (81), Salvia (70), Solanum (65), Dalea (64), Carex (58), Ipomoea (55) y Ageratina (53) (Tabla 4).
Tabla 4 Riqueza por género. Se muestran los 30 géneros más ricos en especies de plantas vasculares nativas de la SMOr.
| Género | Especies | Género | Especies | Género | Especies |
|---|---|---|---|---|---|
| Euphorbia | 93 | Opuntia | 43 | Nama | 33 |
| Quercus | 84 | Sedum | 43 | Asclepias | 32 |
| Mammillaria | 81 | Muhlenbergia | 42 | Cyperus | 32 |
| Salvia | 70 | Tillandsia | 41 | Echeveria | 32 |
| Solanum | 65 | Agave | 40 | Lithospermum | 32 |
| Dalea | 64 | Brickellia | 40 | Senna | 32 |
| Carex | 58 | Echinocereus | 37 | Desmodium | 31 |
| Ipomoea | 55 | Erigeron | 37 | Penstemon | 31 |
| Ageratina | 53 | Turbinicarpus | 36 | Astragalus | 30 |
| Verbesina | 44 | Physalis | 35 | Stevia | 30 |
Riqueza de endemismos. Se registran 1,542 especies endémicas de la provincia fisiográfica de la SMOr en sentido estricto. Las familias con mayor riqueza de endemismos son Asteraceae (328 especies), Cactaceae (193), Fabaceae (94), Lamiaceae (78), Crassulaceae (64) y Asparagaceae (53) que en conjunto suman 810 especies o más del 50 % del total de endemismos de la SMOr.
Los géneros más ricos en especies endémicas son Mammillaria (52 especies), Turbinicarpus (36), Sedum (27), Salvia (26), Erigeron (25), Pinguicula (24), Agave (22), Ageratina (22), Echinocereus (22), Echeveria (21), Dalea (20) y Verbesina (20) (Figura 3).

Figura 3 Géneros de plantas vasculares endémicas más diversas de la Sierra Madre Oriental. En orden decreciente de riqueza fue A) Mammillaria (52 especies), B) Turbinicarpus (36), C) Sedum (27), D) Salvia (26), E) Erigeron (25), F) Pinguicula (24), G) Agave (22), H) Ageratina (22), I) Echinocereus (22), J) Echeveria (21), K) Dalea (20), L) Verbesina (20).
Los géneros más ricos en endemismos de la familia Asteraceae son Acourtia, Ageratina, Brickellia, Cirsium, Dahlia, Erigeron, Flourensia, Grindelia, Packera, Senecio, Stevia, Thelesperma, Verbesina y Vernonia con alrededor de 50 % de las especies. Cinco géneros (Acourtia, Cirsium, Dahlia, Senecio y Verbesina) tienen especies endémicas distribuidas más al sur de San Luis Potosí, el resto de los géneros más ricos se distribuyen en su mayoría en zonas de clima templado y árido de Coahuila, Nuevo León y Tamaulipas. Los géneros más ricos en especies en esta zona son Ageratina, Erigeron y Verbesina, distribuidos en suelos calcáreos.
Para la familia Cactaceae, los géneros más ricos en endemismo son Coryphantha, Echinocereus, Mammillaria, Thelocactus y Turbinicarpus con 134 especies, lo que representa casi 70 % del total, distribuidos principalmente en zonas con fisiografía valles intermontanos, cañones, laderas abruptas y lomeríos de Durango, Coahuila, Nuevo León, Tamaulipas, Zacatecas, San Luis Potosí, Querétaro e Hidalgo asociados a las zonas de clima árido y agrupaciones de vegetación de matorral xerófilo, así como en bosques templados pero en menor proporción . Las tres especies microendémicas de Aztekium se distribuyen únicamente en el estado de Nuevo León, en cañones de paredes de conglomerado yesoso. Turbinicarpus es un género endémico de la SMOr y alcanza su mayor riqueza en las sierras de San Luis Potosí (18) y Tamaulipas (13).
Para la familia Fabaceae, cinco géneros (Astragalus, Bauhinia, Dalea, Lupinus y Phaseolus) representan 58 % de las especies endémicas de esta familia, distribuidas principalmente en la zona norte en los estados de Nuevo León, Coahuila y Tamaulipas, así como en Guanajuato y Querétaro, y aunque también abarcan zonas del Carso Huasteco en estos dos últimos estados, el 63 % de Astragalus y 88 % son de distribución norteña. Por otro lado, las especies endémicas de Lupinus habitan los boques de clima frío en la parte alta de las montañas de la Gran Sierra Plegada y el Carso Huasteco, mientras que las de Phaseolus habitan bosques templados, mesófilo de montaña e incluso matorral submontano en el estado de Durango.
Para la familia Lamiaceae, los géneros Hedeoma, Salvia, Scutellaria y Stachys aglutinan 85 % de las especies endémicas de esta familia en los bosques templados de la Gran Sierra Plegada, Sierras y Llanuras Coahuilenses, Sierras y Llanuras Occidentales y Carso Huasteco. Cabe destacar que el género Hedeoma es diverso en la SMOr. En México existen alrededor de 29 especies, de las cuales 72 % (21 especies) están distribuidas en la SMOr y 44 % (13) son endémicas, a excepción de tres especies, todas se distribuyen en Coahuila y Nuevo León. Las especies endémicas de Salvia (26) se distribuyen principalmente en sierras de Coahuila, Nuevo León, San Luis Potosí, Querétaro y Guanajuato. Para Scutellaria, la mayoría de las especies restringen su distribución a la subprovincia fisiográfica de la Gran Sierra Plegada en Nuevo León y Coahuila.
Riqueza por agrupación de vegetación. Las cuatro agrupaciones que registran la mayor cantidad de especies son el bosque templado con 2,906 especies, seguido del matorral xerófilo con 2,083 especies, el bosque mesófilo de montaña con 1,626 especies y el bosque tropical con 1,623 especies (Figura 4).

Figura 4 Riqueza de especies de plantas vasculares endémicas de la provincia fisiográfica de la Sierra Madre Oriental por agrupación vegetal en orden decreciente.
Diversidad por estado. La riqueza de estados se distribuyó de mayor a menor riqueza como sigue: Querétaro (2,803 especies), Coahuila (2,710), Nuevo León (2,406), Hidalgo (1,851), San Luis Potosí (1,659), Tamaulipas (1,506), Puebla (1,165), Guanajuato (919), Veracruz (686), Zacatecas (438) y Durango (295).
Afinidad geográfica de los géneros. El 39 % de los géneros (2,742 especies) pertenecen presentan afinidad geográfica boreal (neártica o paleártica). Después, el 29 % (2,020) presentan distribución geográfica tropical, que incluye al neotrópico, paleotrópico y todas las demás zonas tropicales del planeta. Posteriormente están los géneros con afinidad geográfica endémica de México con un 17 % (1,227), los cosmopolitas con un 12 % (803) y los géneros que comparten su afinidad geográfica con los desiertos de Argentina, Chile, Sudáfrica y Australia con 3 % (189) (Tabla 5).
Tabla 5 Afinidad geográfica del género. Se muestra la riqueza de especies por su afinidad geográfica actual. Boreal se refiere a la región holártica y paleártica, tropical al neotrópico y paleotrópico, endemismo mexicano al territorio de México, cosmopolita a todo el mundo y desiertos del mundo a regiones desérticas de Sudamérica, África y Australia.
| Afinidad de los géneros | Total de Especies |
|---|---|
| Boreal | 2,742 |
| Tropical | 2,020 |
| Endémico mexicano | 1,227 |
| Cosmopolita | 803 |
| Desiertos del mundo | 189 |
Discusión
Riqueza taxonómica. Para México se estima una riqueza de 24,728 especies de plantas vasculares nativas (incluyendo categorías infra específicas), 297 familias y 2,854 géneros, (Villaseñor 2016, Sosa et al. 2018); por lo tanto, la lista presentada en este trabajo de la flora vascular en la SMOr (6,981 especies) constituye alrededor del 28 % de la flora nacional. Las 225 familias representan alrededor del 75 % y los 1,521 géneros el 53 %. Consecuentemente, la provincia fisiográfica de la SMOr alberga casi una tercera parte de la flora mexicana, rebasando ampliamente la cifra estimada por Luna-Vega et al. (2004) de alrededor de 2,500 especies para esta región.
Riqueza por familias. La repartición de especies por familia sigue la misma tendencia registrada a nivel del país (Rzedowski 1991, Villaseñor 2004, 2016). Sin embargo, la diferencia es que en la SMOr la tercera familia más rica la integra Cactaceae con 365 especies, cuya distribución es transicional hacia el Desierto Chihuahuense (Vázquez-Sánchez et al. 2013).
Las plantas de la familia Asteraceae registradas en la SMOr (1,000) representan alrededor del 32 % de la riqueza nacional (3,057), son en su mayoría hierbas anuales o perennes que ocasionalmente pueden formar estructuras leñosas y con porte arbustico y predominan en todos los tipos de vegetación. Los géneros más ricos son Ageratina (53), Verbesina (44), Brickellia (40), Erigeron (37), Stevia (30), Senecio (29), Acourtia (23), Baccharis (19), Pseudognaphalium (19), Cirsium (19), Roldana (17), Dahlia (16) y Bidens (15). Los géneros más ricos de esta familia se distribuyeron también sobre suelos calcáreos (Villaseñor 2018).
La familia Fabaceae en la SMOr representa 26 % de las 1,903 especies de México (Villaseñor 2016). Se distribuyen en vegetación de clima templado y húmedo. Cuatro géneros de Fabaceae: Dalea (64 especies), Senna (32), Desmodium (31) y Astragalus (30) representan una tercera parte de la riqueza de la familia. Dalea y Astragalus se distribuyen principalmente en valles intermontanos de las Sierras y Llanuras Coahuilenses, mientras que otros géneros también altos en riqueza como Mimosa (24) y Phaseolus (22) se distribuyen en laderas de clima cálido y húmedo en la Gran Sierra Plegada y el Carso Huasteco, lo cual concuerda con resultados de otros autores (Salinas-Rodríguez et al. 2017).
La familia Cactaceae se distribuye principalmente en la vertiente seca de la SMOr en los estados de Coahuila, Nuevo León, Tamaulipas, San Luis Potosí, Guanajuato, Querétaro e Hidalgo; en muchos casos en las laderas bajas de la sierra que forman una franja transicional con la región biogeográfica del Desierto Chihuahuense, en donde tiene importantes concentraciones de endemismos asociados a cañones con depósitos edáficos endurecidos provenientes del carbonato de calcio en forma de conglomerados, yesos y caliche. Para esta familia, en México se estima la existencia de alrededor de 677 especies, de las que las de la SMOr representan alrededor del 52 %. Los géneros Mammillaria (81 especies), Opuntia (42), Echinocereus (37), Coryphantha (29) y Turbinicarpus (36) son los géneros más ricos de esta familia en la zona y representan la tercera parte del total. Varias contribuciones botánicas (Johnston 1943, Hernández & Godínez 1994, Hinton & Hinton 1995, Villarreal-Quintanilla & Encina-Domínguez 2005, Martínez-Avalos & Jurado 2005, Santa Anna-Del Conde et al. 2009, Villarreal-Quintanilla & Estrada-Castillón 2008, Rzedowski et al. 2012) resaltan que las regiones áridas con vegetación xerófila que se ubican en los valles intermontanos de la SMOr, especialmente en los estados de Coahuila, Nuevo León, San Luis Potosí, Tamaulipas, Querétaro e Hidalgo como hábitat de cactáceas que también biogeográficamente pertenecen al Desierto Chihuahuense. Lo anterior se debe a que como mencionan Vázquez-Sánchez et al. (2013) y Guerrero et al. (2019) la tribu Cacteae pudo haber tenido su origen en los enclaves de la SMOr hace aproximadamente 15 millones de años y posteriormente se extendieron al Altiplano Norte (Chihuahuense) y Altiplano Sur (Zacatecano - Potosino) y más tarde, el levantamiento de la Sierra Madre Occidental, aumentó la aridez, promoviendo la expansión de la mayoría de las especies suculentas durante la transición del Mioceno - Plioceno. La región de sotavento de la sierra, correspondiente a zonas áridas y semiáridas con fisiografía de valles intermintanos, lomeríos y bajadas con vegetación de matorral xerófilo, pastizal gipsófilo, pastizal halófilo y chaparral comparte una gran cantidad de especies de esta familia con lo que biogeográficamente se denomina Desierto Chihuahuense o altiplano norte y sur, coincidiendo con los resultados de Villaseñor et al. (2020).
Otras familias de número importante de especies son Poaceae (328 especies), Orchidaceae (189) y Euphorbiaceae (178). La familia Poaceae en México tiene alrededor de 1,216 especies (Dávila et al. 2018) y las especies de la SMOr representan 26 %, se distribuyen en todas las agrupaciones de vegetación y forman pastizales naturales entre las sierras en el valle de Cuatro Ciénegas y el valle de Colombia en el estado de Coahuila, con los géneros Muhlenbergia (42 especies), Bouteloua (25), Aristida (14), Bromus (11), Eragrostis (11) y Achnatherum (7). Al sur de Nuevo León y norte de San Luis Potosí forman pastizales gipsófilos y halófilos con géneros como Sporobolus (12 especies), Digitaria (7), Pappophorum (3) y Distichlis (2), así como también pastizales alpinos con géneros como Muhlenbergia (3), Festuca (2), Piptochaetium (2), Trisetum (1), Brachypodium (1), Calamagrostis (1) y Koeleria (1) en la cima de las montañas de los Estados de Coahuila, Nuevo León y Tamaulipas. En el estado de Guanajuato, Querétaro e Hidalgo, los pastizales naturales son mucho más escasos y la mayoría son pastizales inducidos para el ganado, podemos encontrar géneros como Aristida, Bouteloua, Eragrostis y Muhlenbergia.
La familia Orchidaceae presenta 189 especies en la SMOr, y 15 % de las 1,213 especies que hay en el país (Hágsater et al. 2005). Las condiciones climáticas de los bosques templados de Quercus y Pinus, chaparrales y matorrales submontanos propician la riqueza de especies de orquídeas terrestres de géneros como Malaxis (16 especies), Corallorhiza (9), Schiedeella (8), Govenia (7), Habenaria (7), Bletia (6), Hexalectris (5), Ponthieva (5), Platanthera (3) y Tamayorkis (3); mientras que las altitudes menores a 800 m snm y las condiciones tropicales de barlovento de laderas y cañones con vegetación de bosque mesófilo de montaña, bosque tropical caducifolio, bosque tropical subcaducifolio y bosque tropical perennifolio del Carso Huasteco en los estados de San Luis Potosí, Querétaro, Hidalgo, Veracruz y Puebla, propician la riqueza de epífitas de géneros como Epidendrum (11 especies), Prosthechea (8), Trichocentrum (5), Encyclia (4), Laelia (4), Oncidium (4), Isochilus (3) Lepanthes (3) y Lycaste (2).
La familia Euphorbiaceae en la SMOr también tiene una participación importante en la riqueza y representa alrededor del 25 % de las especies que hay en México (Martínez-Gordillo et al. 2002). Su riqueza se ve reflejada en las áreas con vegetación de bosque tropical que incluye bosque tropical caducifolio, bosque tropical subcaducifolio y bosque tropical perennifolio, así como el matorral xerófilo y el matorral submontano. Los géneros más ricos son Euphorbia (93 especies), Croton (25) y Acalypha (16).
Géneros. El género más rico de la SMOr es Euphorbia (93 especies), género cosmopolita muy variable, representado por plantas por herbáceas que en su mayoría habitan bosques templados y bosques tropicales, aunque también se encuentran en matorrales xerófilos, especialmente en las regiones en transición con zonas de climas secos (Martínez-Gordillo et al. 2002).
Después está Quercus (84 especies), un género de afinidades boreales que tiene una alta riqueza en México. Es el principal componente del dosel arbóreo de los bosques templados de encino o mixtos con coníferas, así como los chaparrales o también llamados encinares arbustivos de la SMOr (Sabás-Rosales et al. 2015, Pérez-Mojica & Valencia-A 2017, Morales-Pacheco et al. 2018). De acuerdo con varios autores, este género se ha diversificado ampliamente en las regiones montañosas de México, adaptándose a las condiciones climáticas de cada región, por lo que presenta características distintivas en cada ecosistema que habita (Valencia-A 2004). La mayor riqueza se ubica en los bosques mixtos de Quercus y Pinus, como por ejemplo Quercus crassifolia, Q. crassipes, Q. laceyi, Q. laeta y Q. laurina, aunque algunas especies son más afines a agrupaciones de vegetación más húmedas como el bosque mesófilo de montaña donde destacan Q. affinis, Q. castanea, Q. germana, Q. insignis, Q. obtusata, Q. rysophylla, Q. sartorii y Q. xalapensis, mientras que otros habitan vegetaciones más frías y secas como el chaparral: Q. arizonica, Q. deserticola, Q. emory, Q. rugosa, Q. sideroxyla, Q. stratula y Q. vaseyana.
Mammillaria (81 especies) es un género de cactáceas para el que se ha propuesto un origen reciente de finales del Mioceno, se cree que se diversificó en las zonas semiáridas del norte de México (Hernández et al. 2001, Hernández & Gómez-Hinostrosa 2011, Hernández & Gómez-Hinostrosa 2015), aprovechando la heterogeneidad ambiental que le proporcionó la vertiente occidental de la SMOr en donde ocurre la transición con el Desierto Chihuahuense, con quien comparte numerosos elementos (Villaseñor et al. 2020) y en donde se registran altas concentraciones de especies de cactáceas, que muestran afinidad por los ambientes rocosos que facilitan la germinación de sus semillas (Peters et al. 2008). Hernández & Gómez-Hinostrosa (2015) mencionan la existencia de seis áreas de alta riqueza del género en México, de las cuales cuatro están dentro de la provincia fisiográfica de la SMOr: a) Jaumave, b) Guadalcázar, c) San Luis Potosí y d) subregión meridional, ésta última región correspondería al desierto Queretano-Hidalguense.
Salvia (70 especies) es un género cosmopolita y el más rico de México. Presenta alta diversidad de especies en las áreas templadas y semiáridas, con especies como Salvia amarissima, S. chamaedryoides, S. coulteri, S. elegans, S. jaimehintoniana, S. melissodora, S. mexicana, S. serpyllifolia, S. stachyoides y S. urolepis (Martínez-Gordillo et al. 2013) y algunas como S. buchananii, S. carranzae, S. calderoniae, S. huastecana, S. xolocotzi y S. zamoranensis son microendemismos del Carso Huasteco y habitan los bosques templados de las montañas de Guanajuato, Querétaro, San Luis Potosí e Hidalgo.
Endemismo. Se calcula que más de 12,900 especies son endémicas de México, o 49-51 % del total de su flora (Rzedowski 1991, Sosa & De-Nova 2012, Villaseñor 2016, Sosa et al. 2018), de las cuales 1,542 son exclusivas de la SMOr (13 % de la flora endémica mexicana), pertenecientes a 109 familias y 484 géneros. Además, la SMOr aglomera 1,455 especies endémicas de México y 3,984 especies nativas que se comparten con Estados Unidos y en menor proporción con Centroamérica.
Rzedowski (2015b) menciona 672 especies endémicas exclusivas en la SMOr. En el presente trabajo se registró más del doble; sin embargo, hemos abarcado un área mayor. Concordamos con Rzedowski (2015b) en que la riqueza de familias la comparten Asteraceae, Cactaceae, Lamiaceae y Fabaceae, y las agrupaciones de vegetación más ricas en endemismos son los bosques templados y los matorrales xerófilos.
Sosa et al. (2018) mencionan que los bosques de encino y pino en las sierras kársticas de la SMOr son áreas de concentración de endemismos y mezcla de neo y paleo endemismos de México, esto concuerda con nuestros resultados. Los mismos autores destacan que las zonas montañosas de clima tropical son las áreas más importantes para el endemismo filogenético, asociadas a la complejidad topográfica y donde las regiones tropicales y subtropicales se traslapan.
Se registran 18 géneros endémicos para la SMOr de los cuales 11 coinciden con Rzedowski (2015b) y se añaden cuatro más a los señalados por Salinas-Rodríguez et al. (2017), así mismo se añaden dos géneros recientemente separados de Turbinicarpus (Vázquez-Sánchez et al. 2019). En orden de riqueza los géneros endémicos son: Turbinicarpus (36 especies), Rapicactus (6), Kadenicarpus (4), Aztekium (3), Mimophytum (3), Strombocactus (3), Geohintonia, Greenmaniella, Jaimehintonia, Mathiasella, Obregonia, Pseudonemacladus, Rzedowskia, Strotheria, Velascoa, Viereckia, Vigethia y Villarrealia (1 especie cada uno).
Mammillaria (52 especies), Turbinicarpus (36) y Echinocereus (22), son géneros especialmente diversos en la confluencia de los estados de Nuevo León, Tamaulipas y San Luis Potosí (Hernández et al. 2001, Hernández & Gómez-Hinostrosa 2011, Vázquez-Sánchez et al. 2013, Hernández & Gómez-Hinostrosa 2015, Guerrero et al. 2019, Vázquez-Sánchez et al. 2019).
Echeveria (21 especies) es un género cuasi endémico de México mientras que Sedum (27) se distribuye en las regiones templadas de todo el planeta; ambos presentan una alta riqueza en México. Son géneros adaptados particularmente a crecer en ambientes rupícolas con sustrato de rocas calizas, en laderas pedregosas, riscos y paredes verticales de la SMOr, en bosques templados, bosques tropicales y matorrales xerófilos con climas estacionales (Pérez-Calix 2008).
Salvia (26 especies) se asocia a bosques templados y fríos de coníferas, bosques de encinos y chaparrales, así como a matorrales xerófilos (Martínez-Gordillo et al. 2013).
Erigeron (25 especies), Ageratina (22) y Verbesina (20) son géneros americanos de asteráceas de amplia distribución en México, pero la mayor riqueza se concentra en bosques de clima templado y frío del centro del país. Sin embargo, en la SMOr también presentan una alta riqueza de endemismos, muchos de ellos se distribuyen en bosque templado con encinos y vegetación gipsófila, así como matorrales de clima semiárido (Suárez-Mota et al. 2017, Villaseñor 2018).
Pinguicula (24 especies) es un género de afinidad boreal y presenta su mayor riqueza en México en la SMOr, la estrecha relación entre varias especies, por ejemplo Pinguicula bustamanta, P. lilacina y P. takakii, podría estar indicando diversificación reciente, estimulada probablemente por el aislamiento geográfico de poblaciones derivadas de un ancestro común, después de la última glaciación (Zamudio 2005, Zamudio & Neváres-De los Reyes 2020).
Agave (22 especies) es un género considerado joven que se originó hace menos de 7 millones de años (Good-Avila et al. 2006, Eguiarte et al. 2021) derivado de una diversificación reciente por los cambios climáticos del pasado y asociado a la alta heterogeneidad ambiental y climática. Su diversidad se ubica principalmente en los bosques templados y en matorrales xerófilos, casi siempre sobre lechos rocosos, cuyas poblaciones a menudo crecen en paredes de 90 ° de inclinación (Salinas-Rodríguez et al. 2017). El hecho de que sea uno de los géneros con más endemismos de la SMOr concuerda con los hallado por Eguiarte et al. (2021) quienes reportan que la diversidad de agaves puede ser alta (más de 20 especies coexistiendo) en algunas áreas dentro de la provincia fisiográfica de la SMOr como porciones de San Luis Potosí y la Barranca de Metztitlán, en Hidalgo.
Concentración de endemismos. Casi la mitad de los endemismos se concentra en la Gran Sierra Plegada (45 %) al norte de la SMOr, en los estados de Coahuila (sierra del municipio de Arteaga), Nuevo León (sierras áridas de los municipios de Galeana, Aramberri, Zaragoza, Doctor Arroyo y Mier y Noriega) y Tamaulipas (valles intermontanos y sierras de los municipios de Jaumave, Miquihuana, Palmillas, Bustamante y Tula) (Salinas-Rodríguez et al. 2017). Dentro de esta misma Subprovincia Fisiográfica hay concentraciones de especies endémicas alpinas que se ubican en las montañas con mayor elevación como el cerro el Potosí (3,722 m snm), la Viga (3,710 m snm), la Marta (3,685 m snm), San Antonio Peña Nevada (3,550 m snm), el Coahuilón (3,545 m snm) y el Viejo (3,492 m snm) en la agrupación de matorral de Pinus y pastizal alpino.
La segunda región con alta riqueza de endemismos es la subprovincia fisiográfica del Carso Huasteco (33 %) al sur de la SMO en los estados de Querétaro, San Luis Potosí e Hidalgo, en zonas de orografía abrupta paisajes kársticos y clima cálido húmedo, con bosques tropicales, bosques templados y un enclave de matorrales xerófilos y matorrales submontanos que se comparten entre Landa de Matamoros en Querétaro y Pacula en Hidalgo (Sosa & De-Nova 2012, Salinas-Rodríguez et al. 2017). Las áreas que concentraron la riqueza se ubican en las montañas altas entre los municipios de Landa de Matamoros en el estado de Querétaro y el municipio de Xilitla en San Luis Potosí, así como los matorrales xerófilos de Querétaro e Hidalgo, que abarcan el Área Natural Protegida Barranca de Metztitlán.
Se registró cierto contraste entre las áreas que concentraron la diversidad y las áreas que concentraron endemismo. Las áreas de alta diversidad, pero con bajo endemismo se ubican en la vertiente de barlovento en los estados de San Luis Potosí, Hidalgo, Veracruz y Puebla, en las regiones que abarcan la agrupación de los bosques tropicales con climas cálido-húmedos.
Por el contrario, las áreas de baja diversidad pero alto endemismo se concentran en la vertiente de sotavento en los estados de Coahuila, Nuevo León, Tamaulipas, San Luis Potosí, Querétaro e Hidalgo, con climas templados y semiáridos y las agrupaciones de matorral xerófilo, chaparral, vegetación gipsófila y halófila, tendencia que no es exclusiva de la SMOr sino de otras áreas montañosas del mundo (Orme et al. 2005).
Diversidad por agrupación de vegetación. La mayoría de las especies son exclusivas de una agrupación de vegetación o máximo dos (85 %), hecho que denota una alta especificidad por su hábitat como también sugieren otras investigaciones (Rzedowski 1991, Villaseñor 2004, Cruz-Cárdenas et al. 2013, Villaseñor 2016). Comparando las agrupaciones vegetales con Villaseñor (2016) la agrupación más rica fue el bosque templado con 2,906 especies, o aproximadamente 50 % de la flora de este ecosistema en México (5,823), seguido del matorral xerófilo con 2,083 especies o 45 % del total para México (4,614). Hay que destacar que ésta última agrupación cubre aproximadamente un 29 % de la vegetación primaria. Aunque abarcan un 0.3 y 3.2 % de la vegetación primaria, las comunidades de bosque mesófilo de montaña y bosque tropical albergan 1,626 y 1,623 especies respectivamente, y el 25 y el 40 % de las especies a nivel país (Luna-Vega et al. 2000).
En la SMOr hay dos agrupaciones de vegetación cuasi endémicas. La primera la denominamos matorral submontano siguiendo a Rzedowski (1978), que se caracteriza por ser un matorral del piedemonte de la SMOr que rara vez sobrepasa los 2,000 m snm, con especies con hojas más grandes que las de los matorrales xerófilos micrófilos, con especies adaptadas a climas subtropicales que pierden sus hojas durante la temporada seca del año y que se ubica entre el matorral espinoso tamaulipeco hacia la Llanura Costera del Golfo y los bosques de encino de las laderas más escarpadas de la SMOr, así como en los cañones estrechos y valles intermontanos en las sierras de Coahuila, Durango, Nuevo León, Tamaulipas, San Luis Potosí, Querétaro e Hidalgo (Estrada-Castillón et al. 2012). Este tipo de vegetación presentó una riqueza de 1,143 especies. La segunda agrupación es un archipiélago de matorrales de Pinus compuestos por poblaciones de pino enano (Pinus culminicola), endémico de las cimas de las montañas más elevadas de los estados de Nuevo León y Coahuila. Rzedowski (1978) lo nombró matorral de Pinus por la intrincada estructura, en esta investigación registramos junto con la pradera alpina, un total de 134 especies, la mayoría endémicas.
En contraste a lo anterior, cabe destacar que más del 56 % de la superficie de la SMOr está ocupada por vegetación secundaria y áreas transformadas por las actividades antropogénicas.
Diversidad por estado. La subprovincia fisiográfica del Carso Huasteco formada por la porción sur de la SMOr ubicada en los estados de Querétaro (2,803 especies) (Rzedowski et al. 2012), Hidalgo (1,851) (Luna-Vega et al. 2000), San Luis Potosí (1,659), Puebla (1,165) y en menor proporción Guanajuato (919) y Veracruz (686) concentra la mayor cantidad de especies. Esto se explica porque aquí se encuentran matorrales xerófilos en la vertiente occidental, bosques templados en las áreas elevadas, bosques tropicales caducifolios en los valles intermontanos y elementos de bosque mesófilo de montaña y de bosque tropical perennifolio en la vertiente oriental de la SMOr (Rzedowski 1963).
La segunda área de alta riqueza es la Subprovincia Fisiográfica denominada Gran Sierra Plegada entre los estados de Coahuila (2,707 especies), Nuevo León (2,402) y Tamaulipas (1,506), también con un amplio gradiente de elevación y a diferencia del Carso Huasteco hay presencia recurrente de heladas. En la vertiente occidental de esta región se encuentran varios tipos de vegetación como: matorral xerófilo, chaparral o encinar arbustivo y vegetación gipsófila y halófila; en las zonas medias se presenta el matorral submontano y en las partes altas de las montañas están los bosques templados, matorral de Pinus y pastizal alpino. Además, en esta subprovincia llega el límite boreal de los bosques tropicales subperennifolios en la Reserva de la Biosfera el Cielo (Valiente-Banuet et al. 1995). Cabe mencionar que la presencia de cañones es común en estos estados y han servido como refugio para las especies endémicas (Hinton & Hinton 1995, Villarreal-Quintanilla & Encina-Domínguez 2005, Villarreal-Quintanilla & Estrada-Castillón 2008, Estrada-Castillón et al. 2012).
Las áreas con menor riqueza de especies se ubican en los valles contiguos a la vertiente de sotavento, en los estados de Zacatecas (438) y Durango (295), aquí la SMOr tiene poca extensión sobre la subprovincia de las Sierras Transversales, donde la sombra orográfica y el efecto de la continentalidad propician condiciones de aridez y un amplio intervalo termal diario.
Afinidad de los géneros. De acuerdo con nuestros resultados, los géneros con distribución geográfica boreal se encuentran en las áreas de mayor elevación de las montañas, en donde predominan climas templados con lluvias estacionales y presencia recurrente de heladas (Hernández & Carrasco 2004, Suárez-Mota et al. 2017). Están ligados al bosque templado, chaparral, matorral de Pinus y pastizal alpino. En este grupo predominan géneros como Alnus, Amelanchier, Arctostaphylos, Berberis, Carya, Cupressus, Heuchera, Juglans, Juniperus, Penstemon, Picea, Pinus, Pseudotsuga, Quercus, Ranunculus, Taxus, Vaccinium y Viola, algunos de ellos con distribución que se extiende hasta las zonas subtropicales y templadas de África, China y Eurasia (Contreras-Medina et al. 2004, Villarreal-Quintanilla et al. 2009, García-Aranda et al. 2012).
Los géneros con distribución geográfica tropical se ubican en la vertiente oriental de la sierra, en la zona de influencia de los vientos alisios húmedos provenientes del Golfo de México, con climas cálido-húmedos y templado-húmedos (Hernández & Carrasco 2004, Suárez-Mota et al. 2017), con influencia continua de neblinas en las zonas medias de las montañas, geomorfología compleja con sierras, cañones, así como relieves kársticos como sótanos, mogotes calizos, dolinas, lapiaces, uvalas y poljes, éstos cuatro últimos localmente denominados “hoyas” que son valles aislados entre las montañas (Clements et al. 2006, Li et al. 2013, Feng et al. 2020). En este grupo se observa un aumento en los géneros de afinidad neotropical, pantropical, incluyendo algunas otras de distribución antillana, africana y asiática, así como una mezcla de especies boreales en la franja de bosques mesófilos de montaña, con riqueza de especies pertenecientes a géneros como Annona, Bauhinia, Begonia, Ceratozamia, Citharexylum, Desmodium, Dioscorea, Elaphoglossum, Ficus, Heliconia, Leucaena, Magnolia, Mimosa, Peperomia, Psychotria, Senna y Vachellia (Rzedowski 1996, Luna-Vega et al. 2000, Luna-Vega et al. 2001, Espinosa et al. 2004, Morrone 2005, Morrone et al. 2017, Villaseñor et al. 2020).
El elemento endémico mexicano con géneros como Agave, Ariocarpus, Coryphantha, Cylindropuntia, Dasylirion, Echeveria, Echinocereus, Hechtia, Hunnemannia, Kadenicarpus, Leucophyllum, Mammillaria, Pachyphytum, Perymenium, Rapicactus, Thelocactus, Turbinicarpus, Vauquelinia, Yucca y Zaluzania, se ubican principalmente en la vertiente de sotavento, con geoformas de valles intermontanos, cañones, mesetas y lomeríos ondulados, con clima árido y vegetación de chaparral, matorral xerófilo, pastizal, vegetación gipsófila y halófila, pero también está muy presente en los bosques tropicales caducifolios y en las áreas transicionales a los encinares (Santa Anna-Del Conde et al. 2009, Ruíz-Sánchez et al. 2012). El hecho anterior concuerda con lo encontrado por Villaseñor et al. (2020) en donde muchos de los linajes endémicos en la SMOr se compartieron estrechamente con los del Desierto Chihuahuense, denominado por ellos como Altiplano Norte y Altiplano Sur.
Los representantes de los géneros cosmopolitas se encuentran dispersos en todas las agrupaciones vegetales, pero especialmente en los bosques templados, matorrales xerófilos y áreas ribereñas de los principales ríos de la SMOr, incluyendo la vegetación acuática (Bonilla-Barbosa 2004). Destacan géneros como Abutilon, Argemone, Aristida, Asclepias, Asplenium, Azolla, Carex, Cyperus, Eleocharis, Erigeron, Euphorbia, Juncus, Ipomoea, Lemna, Plantago, Portulaca, Potamogeton, Selaginella, Schoenoplectus, Solanum y Utricularia.
Los géneros relacionados con del bioma global de los desiertos fue el menos rico, se distribuye principalmente en los matorrales de los lomeríos de Coahuila, Durango, Zacatecas y San Luis Potosí, Querétaro e Hidalgo, cuyos representantes se comparten con los desiertos de Sudamérica, Australia y Sudáfrica, destacando géneros como Allionia, Bahia, Bouchetia, Carlowrigthia, Castela, Chamaesaracha, Condalia, Cryptantha, Erioneuron, Fagonia, Flourensia, Gutierrezia, Halimalobos, Hermannia, Ipomopsis, Larrea, Malvella, Menodora, Nama, Parkinsonia, Thymophylla y Tiquila (Rzedowski 1973, Morrone 2005).
Nuestros resultados concuerdan con los de Villaseñor et al. (2020) quienes encontraron que la mayor parte de la Zona de Transición Mexicana, presenta géneros de afinidad neártica, seguida de la neotropical y finalmente la propia de esta región transicional, además de otra superficie más pequeña dentro de su polígono de la SMOr que tienen afinidades entremezcladas entre la neártica y la netropical. Los mismos autores señalan que para su polígono de la SMOr existen 240 géneros de afinidad neártica y 142 de afinidad neotropical.
Heterogeneidad fisiográfica. Diversos autores señalan que la heterogeneidad topográfica favorece la biodiversidad en las montañas del mundo (Ricklefs 1977, Richerson & Lum 1980, Burnett et al. 1998, Nichols et al. 1998, Badgley et al. 2017), ya que funciona como generadora de nuevos hábitats (Hoorn et al. 2013, Antonelli 2015, Zizka & Antonelli et al. 2018, Rahbek et al. 2019, Perrigo et al. 2020) y puede propiciar la diversificación de las especies. La SMOr comprende paisajes kársticos que generan mircro hábitats (Clements et al. 2006, Li et al. 2013, Feng et al. 2020), que ha propiciado la evolución de géneros de hábitos rupícolas, como: Agave, Brahea, Dasylirion, Dioon, Echeveria, Eucnide, Eutetras, Hechtia, Mammillaria, Nolina, Pinguicula, Sedum, Tillandsia, Verbesina y Villadia, en donde además se han generado múltiples mezclas edáficas que favorecen la diversificación (Figura 5) (During & Willems 1984, Brooks 1987, Van Santen & Linder 2020, Moreno et al. 2020). Estos hábitats son también importantes para grupos que ante la escasez de nutrientes presentan hábitos insectívoros, como Pinguicula adaptada a suelos calcáreos y yesosos, con escurrimientos de agua o con presencia de líquenes (Zamudio et al. 2019), así como el caso de Atriplex, Aztekium, Ephedra, Gaillardia, Geohintonia, Helianthela, Hemyphyllacus, Hunnemannia, Mammillaria, Marshalljhonstonia, Menodora, Nama, Perymenium, Selaginella, Strotheria, Suaeda, Thelesperma y Xylothamnia, muchas de ellas asociadas a suelos calichosos. Aunque la “hipótesis de geobiodiversidad” especula que el levantamiento orogénico, la heterogeneidad del relieve y la diversidad edafológica podrían ser los precursores fundamentales de la riqueza de plantas vasculares, al menos en la cordillera del Himalaya (Mosbrugger et al. 2018), otros estudios (Muellner‐Riehl et al. 2019) han confirmado que la geodiversidad, sumada a factores bióticos contemporáneos y a las fluctuaciones climáticas modernas e históricas, han sido precursores de la biodiversidad.

Figura 5 Las rocas de la provincia fisiográfica de la Sierra Madre Oriental son en su mayoría de origen sedimentario, lo que ha dado lugar a variados sustratos y geoformas que han permitido el crecimiento de los siguientes endemismos: A) conglomerados con Aztekium valdezii, B) calizas con escurrideros de agua y asociados a líquenes con Pinguicula simulans, C) yesos, localmente llamados calichales con Geohintonia mexicana y Aztekium hintonii, D) salinas con Suaeda palmeri, E) paredes kársticas que permanecen con niebla gran parte del año con Nolina robusta, F) sótanos kársticos con Agave muxii.
Diversidad climática. Los procesos orogénicos que formaron la provincia fisiográfica de la SMOr (Eguiluz-De Antuñano et al. 2000, Fitz-Díaz et al. 2018) originaron un ensamble de cinturones termales que la biota ha ido ocupando. Tomando como referencia a Körner et al. (2011), y el hecho de que en la SMOr existe un gradiente altitudinal superior a los 2,800 m snm, en ésta se presentan cinco de los siete cinturones termales que de menor a mayor elevación son: remanente montano sin congelación, remanente montano con congelación, montano inferior, montano superior y alpino inferior (están ausentes el cinturón alpino superior y el cinturón nival), estos se unifican en todas las montañas del mundo a partir de la “línea de árboles” que en la SMOr se ubica aproximadamente a los 3,600 m snm y es el límite entre el cinturón montano superior y el alpino inferior. Consecuentemente, a muy grandes rasgos la flora de la SMOr se distribuiría de la siguiente manera: plantas de afinidad tropical en el cinturón remanente montano sin congelación, especies de afinidad endémica y boreal en el cinturón remanente montano con congelación y cinturón montano inferior y plantas de afinidad exclusivamente boreal en el cinturón montano superior y alpino inferior. Su posición frente a los vientos alisios húmedos crea el efecto de sombra orográfica. Plantas adaptadas a climas lluviosos e isotermales se ensamblan en la vertiente oriental, mientras que especies pertenecientes a los desiertos se distribuyen en la vertiente occidental. Además, hay una diferencia en la radiación que recibe la vertiente norte con respecto a la sur; mientras que la norte es más húmeda y fresca, la sur es más soleada y seca, teniendo un efecto similar en otras regiones montañosas del planeta (Kutzbach et al. 1993, Raupach & Finnigan, 1997, Bennie et al. 2008, Lembrechts et al. 2019).
La SMOr como refugio ante cambios climáticos. Su posición latitudinal y orografía accidentada han sido importantes ante las fluctuaciones climáticas que ha experimentado esta parte del territorio mexicano (Flores-Villela & Goyenchea 2001), como por ejemplo durante el último glacial tardío, cuando las precipitaciones fueron más intensas y permitieron la presencia de bosques en zonas que ahora están ocupadas por matorrales de clima árido (Roy et al. 2020). Especies de climas húmedos pudieron refugiarse en el bosque mesófilo de montaña ante la creciente aridez durante el último máximo glacial (Ramírez‐Barahona & Eguiarte 2013). Los bosques pleistocénicos de las lagunas de lo que ahora es el bolsón de Mapimí, adyacente a la vertiente occidental de la SMOr, fueron refugio de especies de bosque de climas templados e incluso actualmente registramos elementos de distribución boreal como los géneros Draba, Picea y Pseudotsuga, aislados en la cima de las montañas o en áreas proclives a nevadas (Van Devender & Burgess 1985).
La Provincia Fisiográfica de la SMOr alberga más de la cuarta parte de la diversidad de plantas del territorio mexicano y una séptima parte de los endemismos del país. Su antigüedad, aunada a su complejo arreglo geológico y topográfico, propició la generación de nuevos hábitats y zonas de transición de biotas con una mezcla de afinidades destacando dos regiones: la septentrional y la meridional, como mencionan Espinosa et al. (2004), donde la mayoría de sus géneros son de distribución boreal, seguida de los de distribución tropical y finalmente los endémicos.
La vertiente occidental ha fungido como cuna de diversificación para ciertos géneros pertenecientes al elemento endémico mexicano que posteriormente migraron al Desierto Chihuahuense, sus áreas elevadas como áreas de distribución de elementos boreales y la vertiente oriental ha servido como refugio de géneros de afinidad tropical (Luna-Vega et al. 2001).
Las cifras de la riqueza florística de la SMOR presentadas en este manuscrito son preliminares, y con seguridad se incrementarán en la medida que se aumente el esfuerzo de recolecta en áreas remotas y de topografía complicada.
Material suplementario
El material suplementario de este artículo puede ser consultado aquí: https://doi.org/10.17129/botsci.2864

