











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
La familia Rubiaceae Juss. es la cuarta familia más diversa entre las angiospermas con 500 a 700 géneros y aproximadamente 13,800 especies (Standley 1918, Robbrecht 1988, Taylor 2001, Zappi et al. 2009, Lorence & Taylor 2012). Es una familia de distribución cosmopolita, caracterizada por la combinación única de caracteres tales como las hojas opuestas o verticiladas con el margen entero, la presencia de estípulas inter o intrapeciolares, las flores gamosépalas y gamopétalas y el ovario ínfero (Standley 1918, Robbrecht 1988, Taylor 2001, Borhidi 2006, 2012, Lorence & Taylor 2012).
Las rubiáceas fueron divididas en subfamilias por Schumann (1891), quien propuso dos grandes grupos: Cinchonoideae y Coffeoideae. Posteriormente, Verdcourt (1958) reconoció tres subfamilias: Cinchonoideae, Rubioideae y Guettardoideae. Robbrecht (1988) estableció uno de los principales esquemas de clasificación para la familia, donde se diagnostican cuatro subfamilias: Antirrheoideae, Cinchonoideae, Ixoroideae y Rubioideae. En la era molecular, algunos autores reconocen únicamente dos subfamilias: Cinchonoideae y Rubioideae (Robbrecht & Manen 2006). El esquema de clasificación más aceptado divide a Rubiaceae en tres subfamilias: Cinchonoideae, Ixoroideae y Rubioideae (Bremer et al. 1995, 1999, Bremer & Eriksson 2009, Kainulainen et al. 2013). La variación morfológica en el grupo también es evidente a nivel genérico y específico, lo que dificulta su caracterización y conlleva a discrepancias en la precisión de su diversidad.
En México las rubiáceas figuran entre las familias con mayor número de géneros (Figura 1) y especies en inventarios regionales y nacionales, pero hay grandes contrastes en la diversidad total reportada. Entre los esfuerzos por enlistar las Rubiaceae en México destacan: Lorence (1990) mencionó 77 géneros y 533 especies; Rzedowski (1991) estimó la existencia de 80 géneros y 510 especies; Borhidi & Diego-Pérez (2002) registraron 85 géneros y 540 especies; Villaseñor (2003) reportó 93 géneros y 593 especies y Villaseñor (2016) 107 géneros y 707 especies. Mención aparte merecen los tratamientos para la familia de Attila Borhidi (2006), titulados Las Rubiáceas de México donde además de enlistar la diversidad de la familia se incluyen descripciones y claves de identificación para géneros y especies; en su primera edición, Borhidi (2006) reconoció 103 géneros y 582 especies, mientras que, en la segunda edición, Borhidi (2012) incluyó 113 géneros y 666 especies.

Figura 1 Diversidad genérica de Rubiaceae en México. A. Arachnothryx. B. Bouvardia. C. Chiococca. D. Coccocypselum. E. Coutaportla. F. Crusea. G. Deppea. H. Exostema. I. Faramea. J. Hamelia. K. Hillia. L. Hoffmannia. M. Houstonia. N. Manettia. O. Notopleura. P. Palicourea. Q. Posoqueria. R. Psychotria. S. Randia. T. Renistipula. U. Richardia. V. Rogiera. W. Rovaeanthus. X. Sommera. Y. Spermacoce. Fotografías de A. Torres Montúfar
En síntesis, todos estos trabajos registran que la diversidad de Rubiaceae en México va de 77 a 113 géneros y de 510 a 707 especies, las diferencias invitan al análisis de los principales trabajos que tratan a la familia en el país con el objetivo de presentar un panorama general sobre las rubiáceas de México, dilucidar el estado de su conocimiento, generar una lista depurada a partir de propuestas concisas que resuelven problemas taxonómicos y nomenclaturales que provocan inconsistencias en el conteo actual de las Rubiaceae en el país, y presentar un patrón general de su diversidad y endemismo.
Material y métodos
Se creó una base de datos en Microsoft Access® que incluye la lista de especies de los trabajos de Borhidi (2012): Rubiáceas de México (AB, de ahora en adelante) y de Villaseñor (2016): Checklist of the native vascular plants of Mexico (JLVR, de ahora en adelante). A cada género y especie se le asignó la subfamilia y la tribu a la que pertenecen, de acuerdo con literatura reciente (Bremer & Manen 2000, Manns & Bremer 2010, Kainulainen et al. 2013). Del presente trabajo se excluyeron los listados florísticos de Lorence (1990), Rzedowski (1991), Borhidi & Diego-Pérez (2002) y Villaseñor (2003), por presentar información poco actualizada. De igual forma no incluimos el tratamiento de Rubiaceae en Flora Mesoamericana (Lorence & Taylor 2012), dado que solo incluye los taxones de Tabasco, Chiapas, Campeche, Quintana Roo y Yucatán, en cambio este tratamiento se usó para tomar decisiones taxonómicas con respecto a sinonimia y taxones aceptados.
Con la base de datos obtenida se compararon las especies registradas en ambos trabajos (AB y JLVR) y se obtuvo una lista final depurada mediante la consulta de literatura especializada y reciente, que permitió definir los sinónimos nomenclaturales y tomar decisiones taxonómicas controversiales a nivel genérico. Para las especies dudosas se consultó la existencia de ejemplares de referencia en herbarios o bases de datos de diferentes colecciones nacionales e internacionales (MEXU, ENCB, UAMIZ, CHAPA, CHIP, CIB, OAX, SERO, IEB, UIB, FCME, MO, K, F, MI, US, XAL). Los datos de distribución por estado se obtuvieron de lo citado por AB o JLVR y complementado con lo registrado en bases de datos de los herbarios ya mencionados.
Se realizó una curva de acumulación de especies histórica donde para cada especie en México se tabuló el año en que se citó por primera vez, sea por descripción original, mención en tratamiento taxonómico, listados florísticos o notas sobre su distribución. El número de taxones por año se graficó en Microsoft Excel.
El esfuerzo de colecta se analizó mediante el mapeo de todas las colectas georreferenciadas para Rubiaceae en México, disponibles en las bases de datos en línea (www.gbif.org, www.tropicos.org, www.conabio.gob.mx/remib). Con el software QGis 2.8 (Quantum GIS 2018) se sobrepusieron los puntos de colecta en cuadrantes de 0.5 × 0.5, 1 × 1 y 2 × 2 grados, ya que representan diferentes escalas de resolución. Con ello se calculó el porcentaje de superficie territorial que contiene colectas para la familia.
Resultados
Riqueza de Rubiaceae en México. El listado conjunto y depurado de las Rubiaceae de México que se presenta en este trabajo, incluye 111 géneros y 724 especies (Tabla S1). Se excluyen los taxones infraespecíficos y se adicionan los taxones descritos recientemente (2017-2020). Las razones y propuestas que permitieron resolver los problemas taxonómicos a nivel genérico para tener una sola lista de especies se detallan en el apartado de discusión.
En el tratamiento de las Rubiáceas de México, AB registra 114 géneros, 658 especies y 65 taxones infraespecíficos, mientras que el listado florístico de JLVR se enlistan 108 géneros y 707 especies (Material Suplementario 1). Las diferencias puntuales entre el número de especies de ambos trabajos son debidas a: i) 21 especies descritas entre 2012 y 2016 no incluidas en AB (Tabla 1); ii) 38 especies con al menos un ejemplar de respaldo en herbario, no consideradas en alguno de los trabajos (Tabla 2); iii) 13 taxones exóticos no incluidos en JLVR (Tabla 3); iv) 23 sinónimos taxonómicos (Tabla 4); v) 18 sinónimos nomenclaturales (Tabla 5).
Tabla 1 Especies de Rubiaceae descritas entre 2012 y 2016.
| Taxon | Año de descripción | |
|---|---|---|
| 1 | Arachnothryx angustispica | 2015 |
| 2 | Arachnothryx jaliscensis | 2013 |
| 3 | Arachnothryx pumae | 2013 |
| 4 | Borreria myrtilloides | 2013 |
| 5 | Bouvardia pascualii | 2012 |
| 6 | Bouvardia sinaloae | 2013 |
| 7 | Bouvardia tenuis | 2013 |
| 8 | Chomelia anisophylla | 2015 |
| 9 | Bouvardia juarezana | 2014 |
| 10 | Bouvardia lancifolia | 2013 |
| 11 | Guettarda coatzacoalcensis | 2013 |
| 12 | Guettarda elongata | 2015 |
| 13 | Guettarda sotonunezii | 2014 |
| 14 | Hoffmannia altipetens | 2013 |
| 15 | Hoffmannia rzedowskiana | 2014 |
| 16 | Mitracarpus micranthus | 2015 |
| 17 | Psychotria inegi | 2015 |
| 18 | Randia colimensis | 2013 |
| 19 | Randia dionisi | 2013 |
| 20 | Randia lorenceana | 2014 |
| 21 | Randia sepium | 2015 |
Tabla 2 Especies con ejemplares de referencia no incluidas en alguno de los trabajos de Rubiaceae en México.
| Especie | Herbario | |
|---|---|---|
| 1 | Alseis hondurensis1 | MO |
| 2 | Arachnothryx linguiloba2 | MEXU |
| 3 | Arachnothryx skutchii1 | MO |
| 4 | Borreria ocymoides1 | MEXU, MO |
| 5 | Borreria prostrata2 | BM, NY |
| 6 | Bouvardia diversiflora2 | MEXU |
| 7 | Coccocypselum herbaceum1 | MEXU, MO |
| 8 | Coussarea imitans1 | MEXU, MO |
| 9 | Chomelia tenuiflora1 | MO |
| 10 | Deppea amaranthoides2 | MEXU |
| 11 | Deppea densiflora2 | MEXU |
| 12 | Deppea grandiflora2 | MEXU, MO, PTBG |
| 13 | Deppea hamelioides2 | A, BM, F, US |
| 14 | Deppea hoffmannioides2 | MEXU |
| 15 | Galium hintoniorum1 | MO |
| 16 | Geophila cordifolia2 | MEXU |
| 17 | Geophila macropoda1 | MEXU, MO |
| 18 | Guettarda dealbata1 | BR |
| 19 | Guettarda deamii1 | MEXU, MO |
| 20 | Hillia maxonii1 | MEXU |
| 21 | Hoffmannia ghiesbreghtii2 | MEXU |
| 22 | Ixora acuminata1 | MEXU |
| 23 | Mitracarpus rhadinophyllus2 | MEXU, UC |
| 24 | Oldenlandia drymarioides2 | US |
| 25 | Palicourea buchtienii2 | MEXU |
| 26 | Psychotria aguilarii1 | F, MO, CICY |
| 27 | Psychotria cyanococca1 | MEXU |
| 28 | Psychotria gracilenta1 | MO |
| 29 | Psychotria micrantha1 | MEXU, MO |
| 30 | Randia nodifolia2 | INEGI |
| 31 | Rogiera edwardsii1 | F, MO |
| 32 | Rogiera oaxascensis2 | MEXU, MO |
| 33 | Ronabea latifolia2 | MEXU |
| 34 | Sabicea panamensis2 | MEXU |
| 35 | Spermacoce alata1 | MEXU |
| 36 | Stenaria rupicola2 | MEXU |
| 37 | Stenaria sanchezii2 | PTBG, ARIZ |
| 38 | Tessiera lithospermoides2 | PR |
Tabla 3 Taxones exóticos de Rubiaceae en México, no incluidos en el trabajo de Villaseñor (2016).
| Especie | Origen | |
|---|---|---|
| 1 | Cinchona officinalis | Sudamérica |
| 2 | Cinchona pubescens | Sudamérica |
| 3 | Coffea arabica | África |
| 4 | Coffea liberica | África |
| 5 | Dentella repens | Asia |
| 6 | Gardenia augusta | Asia |
| 7 | Ixora coccinea | Asia |
| 8 | Ixora finlaysoniana | Asia |
| 9 | Morinda citrifolia | Asia |
| 10 | Oldenlandia lancifolia | África |
| 11 | Richardia humistrata | Sudamérica |
| 12 | Sherardia arvensis | Europa-Asia |
| 13 | Uncaria tomentosa | Sudamérica |
Tabla 4 Sinonimia taxonómica en los trabajos de Rubiaceae de México.
| Borhidi (2012) | Villaseñor (2016) | |
|---|---|---|
| 1 | Arachnothryx aetheocalymna | *Arachnothryx linguiformis |
| 2 | Arachnothryx sooiana | *Arachnothryx gracilispica |
| 3 | Arachnothryx tabascensis | *Arachnothryx buddleioides |
| 4 | *Diodia apiculata | Diodia aspera |
| 5 | Didymaea microflosculosa | *Didymaea mexicana |
| 6 | Hintonia standleyana | *Hintonia latiflora |
| 7 | Hoffmannia calycosa | *Hoffmannia psychotriifolia |
| 8 | *Hoffmannia longipetiolata | Hoffmannia tonduzii |
| 9 | Hoffmannia teruae | *Hoffmannia psychotriifolia |
| 10 | Hoffmannia uniflora | *Hoffmannia excelsa |
| 11 | Morinda asperula | *Morinda panamensis |
| 12 | Palicourea axillaris | Psychotria aubletiana |
| 13 | Palicourea tomentosa | Psychotria poeppigiana |
| 14 | *Psychotria costivenia | Psychotria miradorensis |
| 15 | *Psychotria cyanococca | Psychotria dispersa |
| 16 | *Psychotria cyanococca | Psychotria pittieri |
| 17 | *Psychotria hoffmannseggiana | Palicourea furcata |
| 18 | *Psychotria pilosa | Psychotria calopogon |
| 19 | Randia nelsonii | *Randia thurberi |
| 20 | Rogiera aprica | *Rogiera cordata |
| 21 | Rogiera ehrenbergii | *Rogiera gratissima |
| 22 | Simira multiflora | *Simira salvadorensis |
| 23 | Simira vestita | *Simira salvadorensis |
* Taxón aceptado en el presente trabajo con base en Flora Mesoamericana (Taylor & Lorence 2012).
Tabla 5 Sinonimia nomenclatural en los trabajos de Rubiaceae de México.
| Borhidi (2012) | Villaseñor (2016) | |
|---|---|---|
| 1 | Borreria nesiotica | Spermacoce nesiotica |
| 2 | Borreria ovalifolia | Spermacoce ovalifolia |
| 3 | Cephalanthus occidentalis subsp. salicifolius | Cephalanthus salicifolius |
| 4 | Houstonia humifusa | Hedyotis humifusa |
| 5 | Houstonia rubra | Hedyotis rubra |
| 6 | Houstonia sharpii | Hedyotis sharpii |
| 7 | Houstonia spellenbergii | Hedyotis spellenbergii |
| 8 | Houstonia teretifolia | Hedyotis teretifolia |
| 9 | Houstonia wrightii | Hedyotis wrightii |
| 10 | Neomartensia breviflora | Martensianthus breviflorus |
| 11 | Neomartensia macdougallii | Martensianthus macdougallii |
| 12 | Neomartensia micrantha | Martensianthus micranthus |
| 13 | Oldenlandia drymarioides | Hedyotis drymarioides |
| 14 | Stenaria mullerae | Hedyotis mullerae |
| 15 | Stenaria nigricans var. nigricans | Hedyotis nigricans |
| 16 | Stenaria umbratilis subsp. umbratilis | Hedyotis umbratilis |
| 17 | Terrellianthus serpyllaceus | Arcytophyllum serpyllaceum |
| 18 | Tessiera hexasepala | Diphragmus hexasepalus |
Asimismo, existen 65 taxones infraespecíficos mencionados en AB la mayoría de los cuales carecen de equivalencia en el trabajo de JLVR. Aquellos que sí son incluidos se debe a que corresponden a la variedad típica o bien son elevados al rango taxonómico de especie (Tabla 6).
Tabla 6 Taxones infraespecíficos incluidos en Borhidi (2012).
| 1 | *Arachnothryx capitellata subsp. capitellata | 40 | *Palicourea galeottiana subsp. galeottiana |
| 2 | Arachnothryx capitellata subsp. pringlei | 41 | Palicourea galeottiana subsp. persearum |
| 3 | *Cephalanthus occidentalis subsp. occidentalis | 42 | Palicourea simiarum subsp. chiapensis |
| 4 | *Cephalanthus occidentalis subsp. salicifolius | 43 | *Palicourea simiarum subsp. simiarum |
| 5 | Chione venosa subsp. mexicana | 44 | Psychotria costivenia subsp. altorum |
| 6 | *Chione venosa subsp. venosa | 45 | *Psychotria costivenia subsp. costivenia |
| 7 | *Crusea coccinea var coccinea | 46 | Psychotria horizontalis subsp. glaucescens |
| 8 | Crusea coccinea var breviloba | 47 | Psychotria horizontalis subsp. glaucina |
| 9 | Crusea hispida subsp. grandiflora | 48 | *Psychotria horizontalis subsp. horizontalis |
| 10 | *Crusea hispida subsp. hispida | 49 | Psychotria panamensis var ixtlanensis |
| 11 | Crusea wrightii subsp. angustifolia | 50 | *Psychotria panamensis var panamensis |
| 12 | *Crusea wrightii subsp. wrightii | 51 | Randia aculeata fo mitis |
| 13 | *Edithea floribunda subsp. floribunda | 52 | *Randia aculeata var aculeata |
| 14 | Edithea floribunda subsp. leuconeura | 53 | Randia aculeata var dasyaclada |
| 15 | Galium fuscum subsp. altiplanicum | 54 | Richardia tricocca subsp. tetracocca |
| 16 | *Galium fuscum subsp. fuscum | 55 | *Richardia tricocca subsp. tricocca |
| 17 | Galium fuscum subsp. guerrericum | 56 | *Rogiera cordata var cordata |
| 18 | *Galium fuscum subsp. hypadenium | 57 | Rogiera cordata var longisepala |
| 19 | Galium mexicanum subsp. asperrimum | 58 | Stenaria nigricans var breviflora |
| 20 | Galium mexicanum subsp. flexicum | 59 | Stenaria nigricans var gypsophila |
| 21 | *Galium mexicanum subsp. mexicanum | 60 | *Stenaria nigricans var nigricans |
| 22 | Galium moranii subsp. aculeolatum | 61 | Stenaria umbratilis subsp. brevipedicellata |
| 23 | Galium moranii subsp. moranii | 62 | *Stenaria umbratilis subsp. umbratilis |
| 24 | Galium sphagnophilum var mazocarpum | 63 | *Syringantha coulteri var coulteri |
| 25 | *Galium sphagnophilum var sphagnophilum | 64 | Syringantha coulteri var loranthoides |
| 26 | *Genipa americana var americana | 65 | Tournefortiopsis crispiflora subsp. cobanensis |
| 27 | Genipa americana var caruto | ||
| 28 | *Guettarda combsii subsp. combsii | ||
| 29 | Guettarda combsii subsp. seleriana | ||
| 30 | Hamelia patens var glabra | ||
| 31 | *Hamelia patens var patens | ||
| 32 | *Hoffmannia macrosiphon var macrosiphon | ||
| 33 | Hoffmannia macrosiphon var multiflora | ||
| 34 | Houstonia acerosa subsp. polypremoides | ||
| 35 | Houstonia acerosa subsp. tamaulipana | ||
| 36 | Houstonia palmeri subsp. muzquiziana | ||
| 37 | Machaonia erythrocarpa subsp. hondurensis | ||
| 38 | Machaonia erythrocarpa subsp. parvifolia | ||
| 39 | Notopleura guadalupensis subsp. tetrapyrena |
*Taxones con equivalencia al rango de especie o variedad típica con equivalencia en Villaseñor (2016)
De ambos trabajos se excluyeron 20 nombres de taxones debido a que su presencia en México es dudosa por carecer de un ejemplar de herbario que lo corrobore (Tabla 7). Del trabajo de JLVR se excluyen cuatro taxones por sinonimia nomenclatural duplicada en ese mismo trabajo: Cigarrilla mexicana (Zucc. & Mart. ex DC.) Aiello /Nernstia mexicana (Zucc. & Mart. ex DC.) Urb., Neomartensia breviflora (Borhidi & Salas-Mor.) Borhidi & Lozada-Pérez/Martensianthus breviflorus (Borhidi & Salas-Mor.) Borhidi, Neomartensia macdougallii (Lorence) Borhidi & Lozada-Pérez /Martensianthus macdougallii (Lorence) Borhidi & Lozada-Pérez y Neomartensia micrantha (Borhidi) Borhidi & Lozada-Pérez/Martensianthus micranthus (Borhidi) Borhidi.
Tabla 7 Taxones dudosos de Rubiaceae en México.
| 1 | Arachnothryx bertieroides2 |
| 2 | Coussarea impetiolaris2 |
| 3 | Chiococca steyermarkii2 |
| 4 | Bouvardia nodiflora2 |
| 5 | Gonzalagunia rudis2 |
| 6 | Guettarda foliacea2 |
| 7 | Hamelia magnifolia2 |
| 8 | Hedyotis greenei2 |
| 9 | Hillia triflora2 |
| 10 | Hoffmannia amplexifolia2 |
| 11 | Hoffmannia sessilifolia1 |
| 12 | Ixora floribunda1 |
| 13 | Notopleura epiphytica2 |
| 14 | Notopleura macrophylla2 |
| 15 | Notopleura siggersiana2 |
| 16 | Pittoniotis trichantha2 |
| 17 | Randia mayana1 |
| 18 | Tocoyena pittieri1 |
Diversidad y endemismo. Las tres subfamilias reconocidas en Rubiaceae se encuentran presentes en México (Tabla 8). Destaca por su diversidad genérica la subfamilia Cinchonoideae con 48 géneros (49 géneros en AB y 51 en JLVR), seguida de Rubioideae con 41 géneros (42 en AB y 44 en JLVR) e Ixoroideae con 19 (17 en AB y 19 en JLVR). A nivel de especie, destaca la subfamilia Rubioideae con 352 especies (321 en AB y 358 en JLVR) seguida de Cinchonoideae con 276 (247 en AB y 260 en JLVR) e Ixoroideae con 99 (90 en AB y 91 en JLVR). A nivel de tribu, en México se encuentran presentes 29 tribus, la más diversa es Spermacoceae, seguida por Hamelieae, Chiococceae y Guettardeae; en estas tribus se concentra alrededor del 50 % de la diversidad genérica y específica de Rubiaceae en México.
Tabla 8 Diversidad por subfamilia y tribu de la familia Rubiaceae en México.
| Sub-familia | Tribu | Borhidi (2012) | Villaseñor (2016) | Listado propuesto en este trabajo |
||||
|---|---|---|---|---|---|---|---|---|
| Géneros | Especies | Géneros | Especies | Géneros | Especies | |||
| 1 | Cinchonoideae | Calycophylleae | 1/0/0 | 1//0/0 | 1/0/- | 1/0/- | 1/0/0 | 1//0/0 |
| 2 | Chiococceae | 10/2/0 | 27/9/0 | 11/2/- | 28/14/- | 9/2/0 | 28/14/0 | |
| 3 | Cinchoneae | 1/0/1 | 2/0/2 | - | - | 1/0/1 | 2/0/2 | |
| 4 | Guettardeae | 10/0/0 | 103/62/0 | 10/0/- | 117/67/- | 9/0/0 | 119/70/0 | |
| 5 | Hamelieae | 14/7/0 | 95/53/0 | 13/7/- | 92/53/- | 14/7/0 | 100/57/0 | |
| 6 | Hillieae | 3/0/0 | 6/1/0 | 3/0/- | 9/2/- | 3/0/0 | 9/2/0 | |
| 7 | Incertae sedis | 1/0/0 | 1/0/0 | 1/0/- | 1/0/- | 3/0/0 | 3/0/0 | |
| 8 | Isertieae | 1/0/0 | 1/0/0 | 1/0/- | 1/0/- | 1/0/0 | 1/0/0 | |
| 9 | Naucleeae | 2/0/1 | 2/0/1 | 1/0/- | 2/0 | 2/0/1 | 3/0/1 | |
| 10 | Rondeletieae | 7/2/0 | 8/2/0 | 7/2/- | 8/2/- | 5/2/0 | 6/2/0 | |
| 11 | Strumpfieae | 1/0/0 | 1/0/0 | 1/0/- | 1/0/- | 1/0/0 | 1/0/0 | |
| 12 | Ixoroideae | Augusteae | 1/0/0 | 1/0/0 | 1/0/- | 1/0/- | 1/0/0 | 1/0/0 |
| 13 | Bertiereae | 1/0/0 | 1/0/0 | 1/0/- | 1/0/- | 1/0/0 | 1/0/0 | |
| 14 | Coffeeae | 1/0/1 | 2/0/2 | - | - | 1/0/1 | 2/0/2 | |
| 15 | Condamineeae | 5/0/0 | 15/6/0 | 6/0/- | 15/5/- | 5/0/0 | 14/5/1 | |
| 16 | Cordiereae | 3/0/0 | 4/1/0 | 3/0/- | 4/1/- | 3/0/0 | 4/1/0 | |
| 17 | Gardenieae | 4/0/1 | 58/41/1 | 2/0/- | 64/43/- | 4/0/1 | 67/45/2 | |
| 18 | Ixoreae | 1/0/0 | 3/0/2 | 1/0/- | 1/1/- | 1/0/0 | 4/1/2 | |
| 19 | Posoquerieae | 1/0/0 | 2/0/0 | 1/0/- | 2/0/- | 1/0/0 | 2/0/0 | |
| 20 | Sabiceeae | 1/0/0 | 3/1/0 | 1/0/- | 2/1/- | 1/0/0 | 3/1/0 | |
| 21 | Sipaneeae | 1/0/0 | 1/0/0 | 1/0/- | 1/0/- | 1/0/0 | 1/0/0 | |
| 22 | Rubioideae | Anthospermeae | 2/1/0 | 2/1/0 | 2/1/- | 2/1/- | 2/1/0 | 2/1/0 |
| 23 | Coussareae | 3/0/0 | 12/7/0 | 3/0/- | 14/8/- | 3/0/0 | 14/8/0 | |
| 24 | Mitchelleae | 1/0/0 | 1/0/0 | 1/0/- | 1/0/- | 1/0/0 | 1/0/0 | |
| 25 | Morindeae | 2/0/0 | 6/0/1 | 2/0/- | 4/0/- | 2/0/0 | 5/0/1 | |
| 26 | Paederieae | 1/0/0 | 1/1/0 | 1/0/- | 1/1/- | 1/0/0 | 1/1/0 | |
| 27 | Palicoureeae | 5/0/0 | 53/14/0 | 5/0/- | 54/11/- | 5/0/0 | 55/11/0 | |
| 28 | Psychotrieae | 2/0/0 | 32/8/0 | 1/0/- | 48/12/- | 2/0/0 | 45/11/0 | |
| 29 | Rubieae | 4/0/1 | 55/28/1 | 3/0/- | 56/33/- | 4/0/1 | 57/34/1 | |
| 30 | Spermacoceae | 24/5/0 | 159/103/3 | 24/4/- | 178/112/- | 23/5/0 | 169/97/3 | |
| Total | 114/17/5 | 658/338/13 | 108/16/- | 707/367/- | 111/17/5 | 724/363/13 | ||
Núm.= Número de Taxones/En= endémicas/Ex=Exóticas
Por su endemismo resalta la tribu Hamelieae con siete géneros endémicos, seguida por Spermacoceae con cinco géneros endémicos, Chiococceae y Rondeletieae con dos géneros endémicos, y, Anthospermeae con un único género endémico de México (Tabla 8). A nivel de especies destaca Gardenieae, Guettardeae y Spermacoceae (con más del 60 % de endemismo) (Tabla 8). Los taxones exóticos de Rubiaceae pertenecen a las tribus Cinchoneae, Naucleeae, Coffeae, Gardenieae y Rubieae (Tabla 8).
En el Tabla S2 se enlistan los 108-114 géneros reconocidos para México en este y en los trabajos previos. Los géneros más diversos son Arachnothryx con 65 especies y 72 % de endemismo (58 especies en AB y 60 en JLVR) Randia con 64 especies y 74 % de endemismo (55 especies en AB y 63 en JLVR) y Bouvardia con 58 especies y 88 % de endemismo (52 especies en AB y57 en JLVR). En total se registran 16 géneros endémicos en el país, nueve de ellos monotípicos, cinco géneros son exóticos en México y cuatro más presentan al menos una especie introducida. Además, México es el centro de diversidad de 10 géneros para los cuales concentra más del 65 % de su diversidad reportada: Deppea (96 %), Bouvardia (80 %), Randia (78 %), Crusea (75 %), Arachnothryx (72 %), Chiococca (70 %), Didymaea (66 %), Glossostipula (66 %), Renistipula (66 %) y Rogiera (65 %) (Tabla S2).
Dentro de México, el estado de Oaxaca destaca por su diversidad (56 % de las especies en México, 51 % de endémicas del país y 81 especies endémicas del estado), seguido de Chiapas (49 % de las especies en México, 48 % especies endémicas del país y 22 especies endémicas del estado), Veracruz (43 % de las especies en México, 30 % especies endémicas del país y 13 especies endémicas del estado) y Guerrero (25 % de las especies en México, 38 % especies endémicas del país y 12 especies endémicas del estado) (Tabla 9).
Tabla 9 Diversidad especies de Rubiaceae por estado en México.
| Estado | Núm | Endémicas del país | Endémicas del estado | |||
|---|---|---|---|---|---|---|
| Borhidi (2012) | Villaseñor (2016) | Listado actualizado |
||||
| 1 | Aguascalientes | 20 | 2 | 3 | 3 | 0 |
| 2 | Baja California | 19 | 7 | 8 | 8 | 5 |
| 3 | Baja California Sur | 33 | 15 | 19 | 18 | 11 |
| 4 | Campeche | 90 | 17 | 18 | 18 | 1 |
| 5 | Chiapas | 365 | 61 | 81 | 80 | 21 |
| 6 | Chihuahua | 45 | 14 | 14 | 14 | 1 |
| 7 | Ciudad de México | 30 | 10 | 9 | 10 | 0 |
| 8 | Coahuila | 39 | 11 | 11 | 11 | 4 |
| 9 | Colima | 96 | 32 | 34 | 34 | 2 |
| 10 | Durango | 49 | 18 | 16 | 16 | 0 |
| 11 | Guanajuato | 43 | 12 | 14 | 14 | 0 |
| 12 | Guerrero | 184 | 71 | 71 | 71 | 12 |
| 13 | Hidalgo | 93 | 34 | 35 | 34 | 0 |
| 14 | Jalisco | 141 | 48 | 52 | 52 | 4 |
| 15 | México | 81 | 29 | 31 | 31 | 1 |
| 16 | Michoacán | 112 | 41 | 41 | 41 | 1 |
| 17 | Morelos | 63 | 21 | 24 | 24 | 0 |
| 18 | Nayarit | 89 | 29 | 29 | 29 | 0 |
| 19 | Nuevo León | 53 | 17 | 17 | 17 | 3 |
| 20 | Oaxaca | 416 | 175 | 202 | 201 | 80 |
| 21 | Puebla | 129 | 37 | 34 | 34 | 1 |
| 22 | Querétaro | 87 | 30 | 31 | 31 | 0 |
| 23 | Quintana Roo | 74 | 12 | 15 | 15 | 0 |
| 24 | San Luis Potosí | 104 | 29 | 31 | 31 | 0 |
| 25 | Sinaloa | 76 | 28 | 31 | 31 | 4 |
| 26 | Sonora | 53 | 14 | 18 | 18 | 0 |
| 27 | Tabasco | 156 | 18 | 24 | 24 | 5 |
| 28 | Tamaulipas | 89 | 24 | 27 | 27 | 2 |
| 29 | Tlaxcala | 17 | 4 | 4 | 4 | 0 |
| 30 | Veracruz | 318 | 81 | 97 | 97 | 13 |
| 31 | Yucatán | 66 | 9 | 9 | 9 | 0 |
| 32 | Zacatecas | 37 | 9 | 10 | 10 | 0 |
Estado de su conocimiento. A través de los años se observa que la cantidad de especies de Rubiaceae ha ido en aumento y muestra un esfuerzo de colecta relativamente constante durante el lapso de la gráfica, aunque se distingue un aumento principalmente con los esfuerzos de exploración a principios del siglo XIX y revisión de colecciones científicas a principios del siglo XXI, alcanzando una aparente estabilidad en décadas recientes (Figura 2).
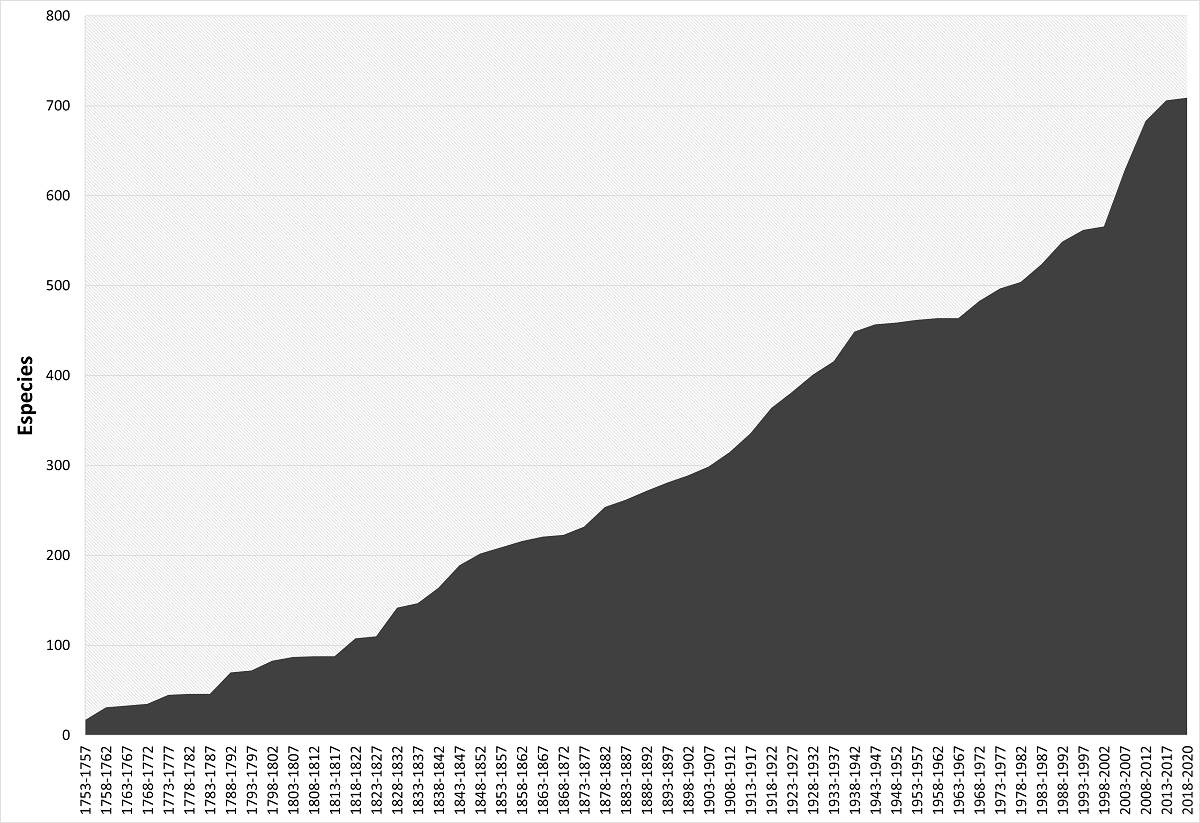
Figura 2 Curva de acumulación histórica de especies descritas de Rubiaceae para México con base en el año en que se describió la especie o fue reportada para el territorio en la literatura especializada
El esfuerzo de colecta de Rubiaceae indica que en la cuadrícula 2 × 2° (222.6 × 222.6 km, ca. 49,000 km2) el 99 % de los cuadrantes tiene al menos una colecta. En la cuadrícula de 1 × 1° (111.3 × 111.3 km, ca. 12,000 km2) el porcentaje de cuadrantes es de 88 %, mientras que en la resolución más fina de 0.5 × 0.5° (55.6 × 55.6 km, 3,000 km2) solamente el 57 % de los cuadrantes que dividen al territorio nacional cuentan con al menos una colecta de Rubiaceae georreferenciada (Figura 3).
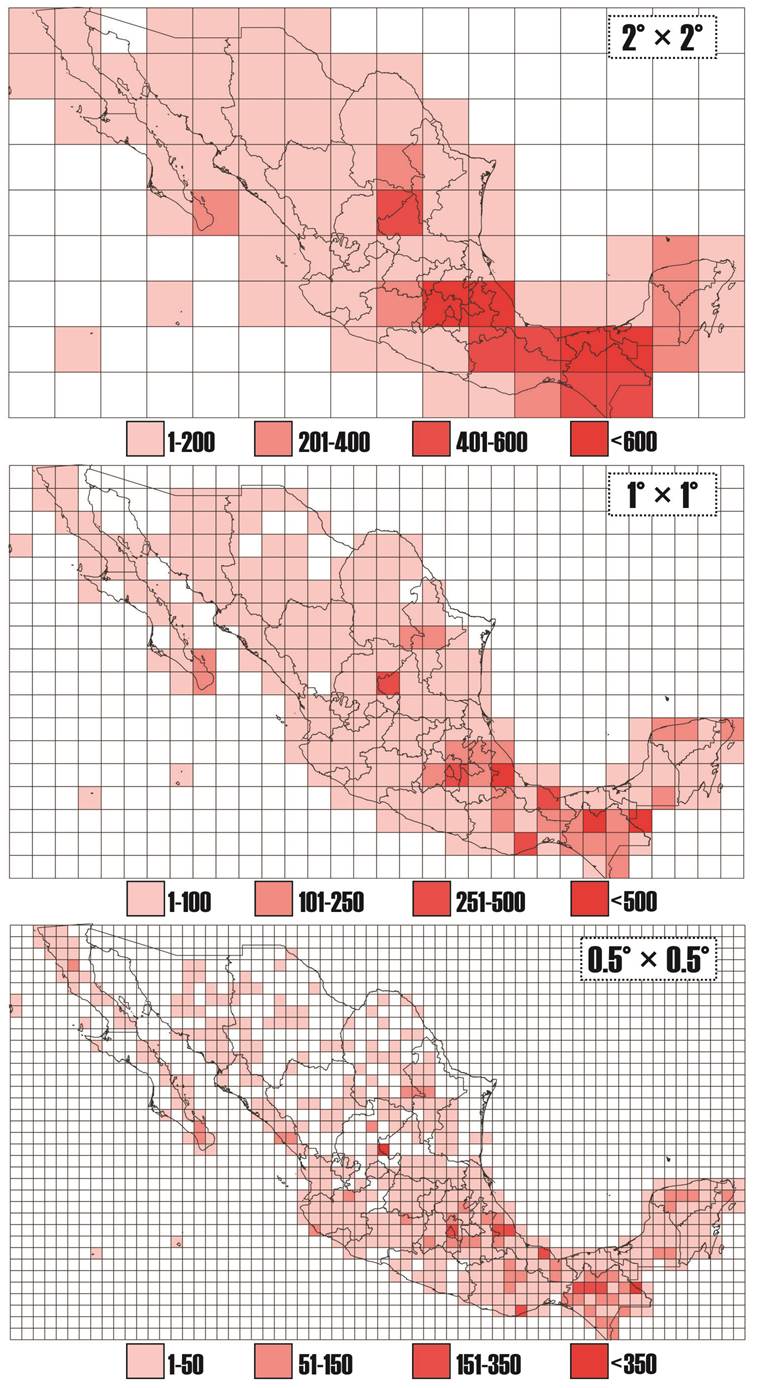
Figura 3 Mapas de esfuerzo de colecta de Rubiaceae en México a diferentes resoluciones: 2° × 2°, 1° × 1° y 0.5° × 0.5° elaboradas en el programa QGis (Quantum GIS 2018). Los colores representan el número de colectas georreferenciadas de Rubiaceae registradas en bases de datos en línea (www.gbif.org, www.tropicos.org, www.conabio.gob.mx/remib).
Discusión
Los dos principales trabajos que a la fecha engloban la diversidad de la familia Rubiaceae en México representan grandes esfuerzos individuales por registrar y conocer uno de los grupos más importantes en la diversidad vegetal mexicana. Entre ambos existen diferencias sustanciales en el número de géneros y especies. Son precisamente estas discrepancias donde se deben focalizar futuros estudios en la familia para México.
Nivel de género. La principal diferencia entre ambos trabajos es a nivel genérico. En AB se reconocen 108 géneros mientras que JLVR menciona 114 géneros. Es en este rubro que se denota la carencia de estudios filogenéticos y revisiones taxonómicas que se tiene de ciertos linajes neotropicales que a continuación de discuten:
Tribu Spermacoceae. Dentro de la tribu Spermacoceae, la más diversa de las tribus en México, existen varias controversias a nivel de géneros, la mayoría de los cuales aún se consideran no resueltas (Dessein 2003, Salas et al. 2011).
Borreria-Spermacoce.- Numerosos taxones han sido transferidos a Spermacoce L. de géneros morfológicamente afines como Borreria G. Mey., Diodia L. y Galianthe Griseb. (Delprete 2007, Salas et al. 2011). En particular la relación Borreria-Spermacoce es objeto de debate, donde algunos autores las consideran entidades separadas (Cabral et al. 2010, 2011) mientras que otros las sinonimizan en un mismo género (Adams & Taylor 2012). La diferencia morfológica principal entre ambos géneros radica en la dehiscencia de los frutos, Spermacoce presenta ambos mericarpios del fruto dehiscentes mientras que en Borreria solo un mericarpio abre para liberar las semillas (Cabral et al. 2011). La evidencia molecular no es conclusiva para separar ambos géneros ya que a la fecha no existe una filogenia molecular que incluya una buena representación de ambos géneros y de la tribu (Karehed et al. 2008, Groeninckx et al. 2009).
Tanto AB como JLVR aceptan ambos géneros, únicamente hay discordancia entre dos especies: Borreria/Spermacoce nesiotica y Borreria/Spermacoce ovalifolia. En el presente trabajo se aceptan ambas especies como pertenecientes al género Spermacoce, por las observaciones realizadas en los frutos de material depositado en MEXU. Se resalta entonces la necesidad de un estudio filogenético integral que evalué los límites genéricos en la tribu Spermacoceae.
Diphragmus-Tessiera.- En JLVR se tratan dos especies bajo el género Diphragmus C. Presl, mientras que en AB las considera como Tessiera DC. El género Tessiera fue descrito con dos especies de distribución marcadamente disyunta: Tessiera lithospermoides DC. de México y Tessiera lanigera DC. de Brasil (De Candolle 1830). De acuerdo con Salas & Cabral (2010), el tipo de T. lithospermoides fue colectado por Haenke en México y depositado en el herbario en Praga (PR), aunque no se menciona la localidad precisa ni fecha de colecta. Por otra parte, el género Diphragmus fue descrito por Presl (1844) y considerado un género monotípico endémico, con base en material también colectado por Haenke en México sin localidad precisa ni fecha de colecta. Salas & Cabral (2010) hacen un recuento histórico y concluyen que T. lithospermoides y Diphragmus scaber C. Presl son sinónimos nomenclaturales.
El género Diphragmus fue considerado como sinónimo de Staelia Cham. & Schltdl. por Standley (1931). Posteriormente, Borhidi & Lozada-Pérez (2006) compararon morfológicamente las especies de Staelia y como resultado revalidaron al género Diphragmus, pese a no consultar su material tipo. En ese mismo trabajo, Borhidi & Lozada-Pérez (2006) describieron Diphragmus hexasepalus Borhidi & Lozada-Pérez para México, propuesta incluida en la primera edición de Rubiáceas de México (Borhidi 2006), mientras que para la segunda edición ya se incluye Diphragmus como sinónimo de Tessiera sin mayor explicación (Borhidi 2012). Se reconoce entonces a Tessiera como un género endémico de México con dos especies: T. lithospermoides y Tessiera hexasepala (Borhidi & Lozada-Pérez) R.M. Salas & E.L. Cabral, dado que la especie brasileña es considerada parte de Staelia (Salas & Cabral 2010).
Complejo Hedyotis.- El género Hedyotis L. es considerado como un taxón controversial con numerosos cambios taxonómicos (Wikström et al. 2013). En particular los géneros presentes en México y en conflicto con Hedyotis: Arcytophyllum Schult. & Schult. f., Houstonia L., Oldenlandia L., Stenaria (Raf.) Terrell y Terrellianthus Borhidi.
En JLVR se aceptan los géneros Hedyotis y Houstonia, mientras que AB trata las especies de Hedyotis como sinónimos ya sea de Houstonia, Oldenlandia o Stenaria. De acuerdo con la filogenia molecular de Wikström et al. (2013) combinando cuatro marcadores plastidiales y nucleares, Hedyotis es un linaje asiático, mientras que las especies americanas corresponden a otros géneros. Con base en esto las especies consideradas como Hedyotis por JLVR son tratadas como Houstonia, Oldenlandia o Stenaria en este trabajo.
En particular, la historia taxonómica de Arcytophyllum-Terrellianthus también se relaciona con Hedyotis-Houstonia. La especie Arcytophyllum serpyllaceum (Schltdl.) Terrell se distribuye en México y en Guatemala (Terrell 1999), fue originalmente descrita en el género Hedyotis, transferida a Houstonia (Greenman 1897), a Arcytophyllum (Donnell-Smith 1893, Standley 1916), Hedyotis (Mena 1990, Terrell 1996) y nuevamente a Arcytophyllum (Terrell 1999).
En el análisis filogenético con evidencia molecular realizado por Andersson et al. (2002) el género Arcytophyllum resulta parafilético con respecto a Hedyotis y a Houstonia, mientras que A. serpyllaceum se encuentra como grupo hermano de Bouvardia ternifolia (Cav.) Schltdl., con base en esos resultados Borhidi & Stranczinger (2012) describieron el género Terrellianthus para incluir la especie A. serpyllaceum, retomando las mismas diferencias que describen Mena (1990) y Andersson et al. (2002).
En JLVR este taxón se trata bajo el género Arcytophyllum, mientras que en AB se le considera Terrellianthus. Si bien la filogenia molecular carece del muestreo adecuado, tanto de terminales como de caracteres, la especie A. serpyllaceum es un linaje diferente al resto de Arcytophyllum, por lo cual en el presente trabajo se adoptó la resolución de AB y Borhidi & Stranczinger (2012) de aceptar Terrellianthus como un taxón presente en México.
Tribu Hamelieae. Deppeopsis.- En JLVR no se reconoce el género Deppeopsis Borhidi & Stranczinger, descrito por Borhidi & Stranczinger (2012) con base en las cápsulas alargadas. Deppeopsis en JLVR es tratado como sinónimo de Deppea Schltdl. & Cham. tal como se hace en el tratamiento de Flora Mesoamericana (Lorence 2012). De acuerdo con la filogenia de Stranczinger et al. (2014), empleando un marcador del cloroplasto y uno nuclear, existe un linaje alejado del clado Deppea que corresponde a las especies de Deppeopsis, que se recuperan en una politomía donde es posible inferir que incluso con el aumento en el muestreo de terminales y marcadores moleculares, Deppeopsis nunca se anidará con Deppea, por lo cual en el presente trabajo reconocemos Deppeopsis como aceptado remarcando la necesidad de un estudio filogenético de este complejo genérico
Tribu Guettardeae. Tournefortiopsis-Guettarda.- En JLVR se incluye el taxón Guettarda crispiflora Vahl, mientras que en AB se trata bajo el nombre Tournefortiopsis crispiflora subsp. cobaensis (Donn. Sm.) Borhidi. De acuerdo con Achille et al. (2006), el género Guettarda L. resulta polifilético en un análisis empleando nrITS, en el cual dos especies de Guettarda (G. crispiflora y Guettarda hirsuta (Ruiz & Pav.) Pers.) se agrupan como hermanas del género Bobea Gaudich. La especie G. crispiflora está morfológicamente relacionada con la especie tipo de Tournefortiopsis Rusby: Tournefortiopsis reticulata Rusby, y por tanto los autores sugieren que podría revalidarse el género con un muestreo mayor de terminales y caracteres. Con base en este resultado parcial Borhidi (2008) revalida Tournefortiopsis con 10 especies, donde además sugiere que Tournefortiopsis crispiflora subsp. crispiflora (Vahl) Borhidi es endémica de Martinica y Montserrat, mientras que la subespecie cobanensis está presente en México.
En nuestro trabajo, decidimos no aceptar al género Tournefortiopsis debido a que su revalidación obedeció a resultados parciales con evidencia molecular procedente de un solo gen nuclear de múltiples copias, en la que además no se incluyó la especie tipo del género, por tanto, aquí se trata bajo el nombre de Guettarda crispiflora, a reserva de estudios filogenéticos particularmente enfocados en esta problemática.
Tribu Condamineeae. Sickingia-Simira.- En JLVR se aceptan los géneros Simira Aubl. y Sickingia Willd., mientras que en AB solo se trata Simira sensu Aublet (1775). De acuerdo con Bremekamp (1954), quien realizó una revisión taxonómica de Simira tinctoria Aubl., redujo a Sickingia como sinónimo de Simira, propuesta que fue retomada por diferentes autores (Steyermark 1972, Delprete & Nee 1997, Taylor & Lorence 2012), a pesar de la ausencia de estudios filogenéticos. En el presente trabajo se sigue está propuesta y sólo se reconoce al género Simira.
Géneros dudosos. Todas las especies incluidas en AB y JLVR son tratadas en Flora Mesoamericana, no obstante, existe controversia en la diversidad genérica donde algunas especies se tratan bajo otros géneros.
Guettardeae-Rondeletieae. Renistipula-Arachnothryx.- El género Renistipula Borhidi fue segregado de Arachnothryx Planch. por Borhidi et al. (2004). Esta segregación no fue aceptada en Flora Mesoamericana por Lorence (2012) en la cual se tratan como sinónimos. Estudios moleculares recientes son contradictorios, por un lado, Stranczinger et al. (2014) ubica a Renistipula en la tribu Hamelieae en lugar de Guettardeae, pero no se incluyeron otras muestras de Guettardeae y menos de Arachnothryx, además que Renistipula no presenta ninguna de las características morfológicas que definen Hamelieae como las inflorescencias ebracteadas, flores amarillas y la presencia de rafidios (Robbrecht 1988). Por otro lado, Torres-Montúfar (2018) encuentra ambos géneros formando un mismo clado sin resolución en Guettardeae, resultado que apoya su sinonimia. Debido a las similitudes morfológicas y a la evidencia molecular en el presente trabajo se reconoce únicamente a Arachnothryx.
Donnellyanthus-Arachnothryx y Rovaeanthus-Rogiera.- Los géneros Donnellyanthus Borhidi y Rovaeanthus Borhidi fueron segregados de Arachnothryx y Rogiera Planch., respectivamente (Borhidi et al. 2004, 2011). Esta segregación no fue aceptada en Flora Mesoamericana por Lorence (2012) en la cual se tratan como sinónimos. Estudios moleculares corroboran a Donnellyanthus y Rovaeanthus como géneros independientes y ubicados en la tribu Rondeletieae en lugar de Guettardeae (Manns & Bremer 2010, Torres-Montúfar 2018, Torres-Montúfar et al. 2018, 2020). Por la evidencia morfológica y molecular, en este trabajo se aceptan.
Tribu Chiococceae. Lorencea-Coutaportla.- El género Lorencea Borhidi fue descrito para incluir la especie Coutaportla guatemalensis (Standl.) Lorence con base en diversos caracteres florales (Borhidi 2003, 2018). Estos atributos fueron considerados como no contundentes debido al alto traslapamiento morfológico y por tanto Lorencea fue tratado como sinónimo de Coutaportla Urb. (Ochoterena 2012). De acuerdo con el resultado de Manns & Bremer (2010), Lorencea es recuperado en un clado sin resolución y con bajo soporte junto con numerosos géneros de Chiococceae incluida la especie Coutaportla ghiesbreghtiana (Baill.) Urb. El trabajo de Paudyal et al. (2014) recupera tres grupos en Chiococceae en un clado sin resolución, donde C. ghiesbreghtiana es un taxón flotante y Lorencea es hermano de Coutarea, sin especificar valores de soporte. Dada la disimilitud morfológica y la evidencia molecular contradictoria, en el presente trabajo se optó por reconocer a ambos géneros, a reserva de futuros estudios filogenéticos particularmente enfocados al grupo.
Solenandra-Exostema.- El género Solenandra Hook. f. fue descrito por Hooker (1873) y posteriormente considerado como sinónimo de Exostema (Pers.) Bonpl. (Schumann 1891, Standley 1921, Liogier 1962, Borhidi & Fernández-Zequeira 1989, McDowell & Bremer 1998, McDowell 2012). Filogenias moleculares que incluyen representantes del género Exostema coinciden en su carácter polifilético por lo que no se han definido fiablemente sus límites genéricos (Rova et al. 2002, Manns & Bremer 2010). Dada la similitud morfológica y la evidencia molecular insuficiente, en el presente trabajo se reconoció únicamente a Exostema a reserva de futuros estudios filogenéticos particularmente enfocados a estos dos géneros.
Tribu Hamelieae. Complejo Deppea.- Además de Deppeopsis, otros géneros morfológicamente relacionados con Deppea no son aceptados en el tratamiento para la familia en Flora Mesoamericana: Bellizinca Borhidi, Csapodya Borhidi y Edithea Standl., los cuales son reconocidos por AB y JLVR. De acuerdo con Lorence (2012), estos taxones se consideran como sinónimos de Deppea por su morfología vegetativa no obstante las claras diferencias en el tamaño y forma de la corola atribuibles a adaptaciones a la polinización por colibríes (Lorence & Dwyer 1988). La evidencia molecular apoya la conclusión de Lorence (2012), ya que los tres géneros se recuperan en un clado sin resolución con las especies de Deppea (Manns & Bremer 2010, Stranczinger et al. 2014). En el presente trabajo se reconocerán estos tres géneros por las evidentes diferencias morfológicas florales, aunado a que los trabajos filogenéticos carecen de una adecuada representación de taxones o un pobre muestreo de caracteres, por lo cual no es robusta la conclusión de sinonimizarlos todos en un solo género.
Sinónimos taxonómicos. Otras diferencias entre el trabajo de AB y JLVR, radican en los sinónimos taxonómicos. En total se incluyen 23 nombres que son tratados como sinónimos en una u otra obra. La mayoría de esta sinonimia se ha propuesto en el tratamiento regional de Rubiaceae de Flora Mesoamericana (2012) por taxónomos especialistas en la familia, en este trabajo se incluyen solo los nombres de las especies reconocidas en Flora Mesoamericana.
Pocos géneros de Rubiaceae en México tienen una revisión taxonómica. La cual impacta en la diversidad de los grupos, tal es el caso de Rogiera donde Torres-Montúfar et al. (2021) reducen a sinonimia dos de las especies reconocidas por AB y JLVR: Rogiera tabascensis Borhidi y Rogiera oaxacensis Borhidi & K. Velasco. Es lógico pensar que la profundización del conocimiento taxonómico permitirá dilucidar la sinonimia y también describir nuevos taxones de la familia en México.
Redundancias taxonómicas. Neomartensia-Martesianthus y Cigarrilla-Nernstia.- El trabajo de JLVR presenta algunas inconsistencias que llevan claramente a sobreestimar la diversidad genérica y específica de Rubiaceae en México, ejemplo de esto es la duplicidad de taxones que poseen el mismo tipo, es decir sinónimos nomenclaturales.
De acuerdo con Villaseñor (2016) en México se encuentra presente Nernstia mexicana (Zucc. & Mart. ex DC.) Urb., descrita en 1923, y Cigarrilla mexicana (Zucc. & Mart. ex DC.) Aiello publicada en 1923, ambos nombres tienen como basónimo a Coutarea mexicana Zucc. & Mart. ex DC. descrito con base en la colecta realizada por Martius en 1829.
De igual manera ocurre con los géneros Neomartensia Borhidi & Lozada-Pérez y Martensianthus Borhidi & Lozada-Pérez, para los cuales JLVR menciona tres especies, pero que poseen el mismo tipo nomenclatural: Neomartensia breviflora=Martesianthus breviflorus (Tipo: Zárate-Marcos 348), Neomartensia macdougallii=Martesianthus macdougalli (Tipo: MacDougall s.n.) y Neomartensia micrantha=Martesianthus micranthus (Tipo: Ramírez 731B). El género Neomartensia fue descrito por Borhidi & Lozada-Pérez (2010), pero es un homónimo posterior de un género de algas descrito en 1996; los mismos autores describieron un año después el género Martensianthus para corregir este error taxonómico (Borhidi & Lozada-Pérez 2011).
Nivel de especie e infraespecíficos. Las diferencias en contenido de especies se explican en su mayoría a la descripción de nuevos taxones en el periodo entre ambas publicaciones. Si se excluyen las especies descritas entre 2012 y 2016, los taxones introducidos y las redundancias taxonómicas en JLVR, encontramos que los números de especies nativas de México aun así son muy disímiles entre ambos trabajos: AB con 645 especies y JLVR con 684 especies.
Los 65 taxones infraespecíficos enlistados por AB, subespecies o variedades, tienen correspondencia con especies enlistadas en JLVR. Únicamente existe una discrepancia de un taxón: Cephalanthus occidentalis var. salicifolius (Bonpl.) A. Gray y Cephalanthus salicifolius Bonpl.
La especie C. salicifolius fue descrita por Bonpland en 1809, posteriormente, Asa Gray en 1844 la consideró como una variedad de Cephalanthus occidentalis L. con base en la forma de las hojas. En nuestro trabajo se acepta la propuesta de Standley & Williams (1975), quienes en el tratamiento de Rubiaceae de Flora de Guatemala descartan que sea una variedad y la consideran en el rango de especie.
En nuestro listado de Rubiaceae no incluimos taxones infraespecíficos, a reserva de revisiones y tratamientos a nivel genérico, así como de la evaluación de su rango taxonómico.
Información incompleta y taxones dudosos. Ambos trabajos son un esfuerzo monumental por incrementar el conocimiento botánico en México, no obstante, no están exentos de algunos vacíos de información, el más relevante es que en ninguno de estos trabajos se menciona la consulta de colecciones biológicas. El tratamiento de AB presenta descripciones de las especies y claves de identificación, sin incluir un apartado de ejemplares examinados de herbario. El trabajo de JLVR es un listado que usa bases de datos de colecciones biológicas y bibliografía, por lo cual no especifica en qué herbario se puede localizar el espécimen/ejemplar de cada especie, aunque detalla muy bien la distribución por estado de las especies.
Entre ambos se enlistan un total de 18 especies para las cuales no existe un ejemplar que corrobore su presencia en México cuya presencia puede considerarse muy probable en México, debido a que se desarrollan en países de Centroamérica en la región Mesoamericana o bien en Estados Unidos como parte del desierto de Sonora, regiones fitogeográficas que se extienden a través de la frontera hacia México. Por lo tanto, no se descarta su existencia en el país a reserva de exploración o revisión de colecciones biológicas más restringidas a la flora de los estados.
Algunos taxones tienen injerencia directa en la diversidad genérica reportada. Por ejemplo, el considerar Pittoniotis trichrantha Griseb. como presente en México incrementa el número de géneros reportado por JLVR en el país, para esta especie hay registros desde Guatemala hasta Colombia. La especie Tocoyena pittieri está registrada desde Colombia hasta Honduras, AB la cita para México, lo cual incrementa la diversidad genérica reportada. Para ambos taxones no existe un ejemplar físico citado en colecciones biológicas consultadas para este trabajo, pero optamos por considerarlas como potencialmente distribuidos en México ya que varias especies de Rubiaceae presentan patrones de distribución similares (p. ej. Chomelia tenuiflora Benth. o Randia loniceroides Dwyer & Lorence).
Otros nombres dudosos presentes en JLVR son “Bouvardia nodiflora Bohr. & I. García” y “Rogiera oaxacana” los cuales no se han encontrado en ninguna publicación de Attila Borhidi en Acta Botánica Hungarica, revista en la que publica todos sus taxones nuevos, y tampoco se encuentra en bases de datos de los herbarios consultados, por lo cual no se consideran estos nombres en el presente listado.
Taxones exóticos. Entre las especies de Rubiaceae reconocidas por Borhidi (2012), se cuentan 13 especies en 10 géneros, que son consideradas introducidas o exóticas, algunas ya como como especies naturalizadas o escapadas de cultivo. De estas, las más comunes son el café (Coffea arabica L.), ampliamente cultivada y consumida en el país e Ixora coccinea L., planta ornamental apreciada en jardines de regiones tropicales y subtropicales en México. Para las especies naturalizadas mencionadas se requieren estudios que ayuden a conocer mejor su comportamiento como malezas y su influencia en las comunidades vegetales donde se les encuentra.
Diversidad por estado. Los estados con mayor riqueza de especies de Rubiaceae contienen también una gran diversidad florística general (Chiapas, Oaxaca o Veracruz), por ejemplo, para Apocynaceae (Juárez-Jaimes et al. 2007, Alvarado-Cárdenas et al. 2020), Cucurbitaceae (Lira et al. 2002), Solanaceae (Martínez et al. 2017) o Asteraceae (Villaseñor 2018).
A nivel de especie, Oaxaca, Chiapas y Veracruz acumulan 577 especies con más del 50 % de nivel de endemismo, es decir que casi el 80 % de la diversidad de Rubiaceae en México se concentra en estos tres estados. La razón de la alta diversidad en estos estados es multifactorial, estas entidades presentan intrincada fisiografía que se traduce en diversos tipos de vegetación: bosque tropical caducifolio, bosque tropical perennifolio, bosque templado, bosque mesófilo de montaña y matorral xerófilo (Domínguez-Vázquez et al. 2002, Morrone 2005, García-Mendoza & Meave 2011, Luna-Vega et al. 2013), para Rubiaceae es particularmente importante la amplia extensión de vegetación con afinidad tropical dado que son centros de diversidad de la familia (Robbrecht 1988). Otro factor determinante puede ser el sesgo en los estudios botánicos e inventarios florísticos, JLVR señala que estos estados han sido objeto de numerosas exploraciones botánicas históricas y contemporáneas.
Conservación. A pesar de la diversidad, el alto grado de endemismo y la importancia ecológica de la familia, pocas especies se encuentran bajo alguna categoría de protección. La pérdida del hábitat es un factor evidente que amenaza su sobrevivencia.
Solo una especie aparece en el Apéndice I de la CITES (2020) bajo la categoría de especie en peligro de extinción. Esta pertenece al género monotípico Balmea Martínez que se distribuye en la vertiente del Pacífico desde México hasta el Salvador; la especie B. stormiae Martínez se menciona que se usa como árbol de navidad localmente y por eso su extracción y comercialización está restringido.
En la Norma Oficial Mexicana, NOM-059-SEMARNAT-2018 (SEMARNAT 2018), que tiene por objeto identificar las especies de flora y fauna silvestres en riesgo, se incluyen 20 especies, de las cuales 14 son endémicas a México y cinco de ellas se distribuyen en la frontera con Guatemala y compartidas con este país. De esta lista resalta Galium pilosum Aiton para la cual no existe registro de su presencia en México en ningún trabajo de Rubiaceae. Es una especie conocida del norte de Estados Unidos, por lo cual su presencia en la NOM-059 es cuestionable.
Para algunos géneros de Rubiaceae es necesario hacer esfuerzos de exploración, dado que son linajes posiblemente extintos por ser únicamente conocidos de la colecta tipo realizada más de un siglo atrás: Habroneuron Standl., Placocarpa Hook. f. y Stylosiphonia Brandegee.
El género monotípico mexicano Habroneuron, fue colectado por Hartweg en 1839 en Oaxaca, presumiblemente en la región de la Chinantla, Oaxaca. La pertenencia de este género a una tribu de Rubiaceae aún es desconocida dada la carencia de frutos en el material tipo (Torres-Montúfar et al. 2020). Recientes esfuerzos de exploración en la zona de la Chinantla no la reportan; sin embargo, estos trabajos sólo abarcan unas porciones de esta región, por lo que nuevas exploraciones son requeridas para la búsqueda de este taxón.
El género monotípico Placocarpa (Chiococceae, Cinchonoideae) fue colectado por Botteri en 1855 en la región de Orizaba, Veracruz. Sin tener una localidad precisa, y dado la presión demográfica en algunas zonas del centro de Veracruz (Williams-Linera et al. 2002, Bonilla-Moheno & Aide 2020), es posible que este taxón esté extinto.
El género Stylosiphonia fue descrito a partir de una colecta de Purpus en 1913, en el estado de Chiapas, sin precisar la localidad y nunca más ha sido recolectado. La pertenencia de este género a una tribu de Rubiaceae aún es desconocida dada la carencia de frutos en el material tipo (Torres-Montúfar et al. 2020). En el estado de Chiapas existe una gran presión ambiental y deforestación (Ochoa-Gaona & González-Espinoza 2000, Cayuela et al. 2006), por lo que es dudosa la supervivencia de este taxón. Sin embargo, en el estado aún existen zonas de difícil acceso no colectadas, lo cual abre la posibilidad de encontrarlo en el futuro.
Un caso de indudable éxito de nueva exploración ocurrió con el género monotípico y endémico de México Eizia Standl. (Hamelieae, Cinchonoideae). Eizia mexicana Standl. se conocía solo de la localidad tipo y de la colecta de Eizi Matuda en 1938, hasta que el grupo de botánicos de la Universidad de Chiapas (UNICACH) la redescubrió en 2015 en otra localidad cercana (Martínez-Camilo et al. 2015), demostrando que aún se requiere exploración en el estado.
Esfuerzo de colecta. Los esfuerzos de colecta de Rubiaceae indican que entre más pequeña sea la escala de la cuadrícula menor porcentaje de territorio posee una colecta registrada y viceversa a mayor escala prácticamente todo el territorio tiene una colecta de Rubiaceae (Figura 3). La ausencia de colectas de la familia en los diversos tamaños de cuadrícula no necesariamente indica que todavía faltan numerosas regiones de México por explorar, algunas corresponden a zonas donde el uso de suelo es urbano o agropecuario y por ello no hay colectas de plantas en general (resultados no mostrados) o bien pueden ser áreas estudiadas donde efectivamente se carece de representantes de Rubiaceae, no están correctamente georreferenciadas o no hay georreferencias disponibles. Es de resaltar que las áreas con menor porcentaje de colecta de Rubiaceae se ubican principalmente en las zonas áridas de México, donde la diversidad de la familia es menor, aunque también corresponden a algunas de las zonas menos exploradas y con menor cantidad de inventarios (Villaseñor 2016) en el territorio nacional.
La exploración y revisión de colecciones científicas ha derivado en que seis nuevas especies de Rubiaceae en México se hayan descrito desde 2016, mismas que se anexan al listado final de la familia en México (Tabla S1): Omiltemia guerrerensis Lozada-Pérez & J. Rojas Gut. (Lozada-Pérez & Rojas-Gutiérrez 2016), Hoffmannia uxpanapense Lozada-Pérez & J. Rojas Gut. (Castillo-Campos & Bautista-Bello 2017), Chiococca grandiflora Lorence & T. Van Devender (Lorence et al. 2018) Arachnothryx axillaris Torr.-Montúfar & Ochot.-Booth, Arachnothryx flores-olverae Torr.-Montúfar & Ochot.-Booth (Torres-Montúfar & Ochoterena 2018) y Rogiera metztitlensis Torr.-Montúfar, E. Solano, Morales-Gard. & Ochot.-Booth (Torres-Montúfar et al. 2019), todas ellas endémicas a México.
Con todo lo anterior, la familia Rubiaceae resalta como un linaje de gran importancia en la flora de México por sus niveles de riqueza y endemismo. El tratamiento de las Rubiáceas de México y la lista actualizada de las plantas vasculares son obras que nos acercan a la gran diversidad de la familia, por su parte nuestro trabajo pretende dar un panorama más preciso de las Rubiaceae de México y que los datos aquí discutidos guíen los futuros esfuerzos taxonómicos para incrementar el entendimiento de la familia, desde la exploración de nuevos territorios, la consulta y curación de colecciones biológicas, hasta focalizar los grupos que requieren ya sea tratamientos, o bien, estudios filogenéticos que ayuden a esclarecer los problemas taxonómicos y acercarnos aún más a conocer la diversidad de las rubiáceas en México.
Material suplementario
El material suplementario de este artículo puede ser consultado aquí: https://doi.org/10.17129/botsci.2847

