











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkEn los ecosistemas de montaña, la altitud, el microambiente y el clima juegan un rol importante en la distribución y estructura de las comunidades vegetales. Particularmente, el bosque mesófilo de montaña (BMM) (Rzedowski 1978), conocido también como bosque de niebla en México, y en inglés como “Tropical Montane Cloud Forest” (Hamilton et al. 1995), el cual alberga elementos de distinta afinidad biogeográfica, representa una gran oportunidad para evaluar las relaciones entre microambiente, clima, cambios en la estructura de la vegetación y su composición florística. Esta última ya ha sido documentada previamente a nivel mundial (Vázquez-García 1995, Hamilton et al. 1995, Bruijnzeel et al. 2010) y en México-Centroamérica (Lieberman et al. 1996, Alcántara-Ayala & Luna-Vega 1997, Catalán-Heverástico et al. 2003, CONABIO 2010, Villaseñor 2010, Williams-Linera et al. 2013). La riqueza florística y rasgos estructurales de los bosques de niebla han sido relacionados con la altitud, y se han encontrado tendencias contrastantes, tales como unimodales, de disminución o aumento lineal monotónico (Tang & Ohsawa 1997, Rahbek 2005, Homeier et al. 2010, Salas-Morales & Meave 2012, Krömer et al. 2013, Williams-Linera et al. 2013). Estas tendencias varían entre regiones y entre las escalas consideradas para el análisis de los intervalos altitudinales, es decir, también han sido encontradas en áreas extensas donde se estudió toda la vegetación en transectos con presencia de bosques tropicales caducifolios hasta bosques subalpinos, y no solamente el BMM (Kitayama 1992, Vázquez-García & Givnish 1998, Reich et al. 2010, Hemp 2010, Toledo-Garibaldi & Williams-Linera 2014).
En el sureste de Asia algunos investigadores han separado los bosques de niebla (el término BMM solo es utilizado en México) en bosques de niebla de baja y alta montaña (lower montane cloud forest y upper montane cloud forest) y bosques de niebla dominados por coníferas (Ohsawa 1991, 1995), en relación con la franja latitudinal y altitudinal en la que estén distribuidos. Además, varios autores han tratado de distinguir límites discretos (Grubb 1977, Hemp 2010, Martin et al. 2011, Bach & Gradstein 2011), aunque otros argumentan que los cambios estructurales y en composición de especies ocurren continuamente a través del gradiente (Lieberman et al. 1996). En contraste, en México no se ha aceptado el enfoque de separar el BMM en fajas altitudinales distintas, aunque en algunos estudios se han reconocido subtipos de BMM y distintas especies dominantes (Alcántara et al. 2002, Mejía-Domínguez et al. 2004, Williams-Linera et al. 2013). Un ejemplo de los distintos ensambles, componentes dominantes y subtipos de BMM son las ecotonías en su límite superior de distribución altitudinal que forman con los bosques de Abies (abeto, oyamel o pinabete). Estas ecotonías suelen presentarse en el occidente y sur de México, incluso hasta Guatemala (Ohsawa 1995, Cuevas-Guzmán et al. 2011, Guerrero-Hernández et al. 2014); sin embargo, ninguna investigación ha examinado específicamente la transición entre BMM y el bosque de Abies, y la relación de ambas comunidades vegetales con variables microambientales y climáticas a lo largo de gradientes altitudinales.
La porción occidental de Jalisco alberga archipiélagos de bosque mesófilo de montaña, los cuales, al igual que en la vertiente del Golfo de México, tienen una alta riqueza de especies arbóreas raras y con distribución restringida (Vázquez-García 1995, Jardel-Pelaez et al. 1996, Vázquez-García et al. 2000, Sánchez-Rodríguez et al. 2003, Guerrero-Hernández et al. 2014). A pesar de esto, la mayoría de estos bosques no figuran entre los hotspots de biodiversidad en la WWF, excepto Sierra de Manantlán (Cuevas-Guzmán et al. 1997). Por otro lado, probablemente los bosques de Abies ocupan mayores extensiones que los BMM, pero son relictos de una distribución que fue aún más amplia durante el Mioceno tardío (Graham 1999), y en la actualidad se distribuyen mayormente en zonas altas y frías de la Faja Volcánica Trans-Mexicana. Al igual que otros taxa de origen templado, el género Abies ha estado expuesto a una dinámica de “islas en el cielo” como respuesta a fluctuaciones climáticas (Mastretta-Yanes et al. 2015). Dada la distribución insular y restringida que exhiben estas comunidades vegetales, es necesario generar información de la estructura y dinámica vegetal para implementar estrategias de manejo y conservación tanto de los BMM como de los bosques de Abies en el occidente de México.
En esta investigación se exploran tendencias en composición de especies leñosas, cambios en la estructura de la vegetación, microambiente y clima en relación con la altitud en dos localidades del occidente de México. Las hipótesis que se plantearon son que en el área de estudio (1) con el incremento de la altitud, hay un cambio progresivo y continuo en la distribución de las especies dominantes, estructura de la vegetación y una disminución monotónica de la riqueza de especies leñosas; (2) el reemplazo del bosque mesófilo de montaña por bosque de oyamel está positivamente asociado con la altitud, disminución de temperatura y mayor disponibilidad de agua en los meses más fríos y secos del año. Nuestros objetivos específicos fueron (1) analizar, en dos gradientes altitudinales, cambios en la estructura de la vegetación y composición de especies leñosas, con base en área basal, altura, abundancia y riqueza; (2) examinar las relaciones entre variables climáticas, factores microambientales y la estructura de la vegetación.
Materiales y métodos
Área de estudio. El área de estudio comprende dos localidades con bosque mesófilo de montaña y bosque de Abies, que se encuentran en la Sierra de Jolapa, en el occidente del estado de Jalisco, México (Figura 1). Se delimita entre las coordenadas 20° 45’ 15” y 20° 38’ 07” N y 104° 50’ 29” y 104° 42’ 47” O (DATUM WGS 84), en los municipios de Mascota y San Sebastián del Oeste. Se registraron dos intervalos altitudinales, el primero abarca de los 1,500 a los 2,450 m en la localidad de La Bufa (B), y el segundo de los 1,750 a los 2,420 m en la localidad de Laguna Juanacatlán (J). La mayor parte de esta superficie está localizada dentro del Área de Protección de Recursos Naturales Río Ameca-Sierra de Vallejo. Se ubica en la cuenca hidrológica Ameca-Ixtapa, en la provincia morfotectónica de la Faja Volcánica Trans-Mexicana, en una zona donde converge con algunos afloramientos ígneos intrusivos de la Sierra Madre del Sur (Gómez-Tuena et al. 2005). De acuerdo con las divisiones florísticas de México propuestas por Rzedowski (1978), la zona se localiza en la provincia de las Serranías Meridionales y éstas a su vez en la Región Mesoamericana de Montaña. El clima, de acuerdo a la clasificación de Köppen modificada por García (2004), es templado subhúmedo con verano fresco y largo, con lluvias en verano con un porcentaje de lluvia invernal < 5 %, el más húmedo de los subhúmedos [C (w2) (w)]. Tiene una temperatura media anual que va de los 18 °C, en las partes más bajas a 13.2 °C en las zonas con mayor altitud; y una precipitación anual de 1,183 a 1,256 mm (Hijmans et al. 2005).

Figura 1 Localización del área de estudio. Dos localidades de estudio, La Bufa y Laguna Juanacatlán. Cuadros con relleno rojo son los sitios de estudio, etiquetados con letra B (Bufa) o J (Laguna Juanacatlán) y un número correspondiente a su altitud en metros.
Diseño de muestreo. Se realizó un muestreo dirigido a los BMM y a los bosques de Abies mejor conservados, sin disturbios antrópicos evidentes, mediante recorridos de campos en dos localidades. El método fue el utilizado por Vázquez-García & Givnish (1998), con unidades de muestreo de 0.1 ha, las cuales se ubicaron de la siguiente manera: en 32 parcelas de 60 × 48 m, cada una dividida a su vez en 20 subparcelas de 12 × 12 m, se seleccionaron de manera estratificada y aleatoria 10 de éstas (5 a cada lado de una línea que dividió la parcela mayor en 2 partes de 24 × 60 m). En el centro de cada una de las 10 subparcelas seleccionadas se colocó una estaca, a partir de la cual se trazó un círculo de 5.64 m de radio para obtener un área de 100 m² por subparcela, y en su conjunto de 0.1 ha por unidad de muestreo. En cada círculo de 100 m² se identificó, registró y midió el diámetro a la altura del pecho (DAP) y la altura, de cada uno de los individuos de las especies leñosas con diámetro ≥ 2.5 cm. Adicionalmente, en cada uno de los 32 sitios se registraron datos de variables edáficas, geográficas y topográficas. Las variables geográficas y topográficas recopiladas fueron coordenadas de latitud y longitud, altitud, exposición, pendiente (Tabla 1) y posición topográfica. Se eligieron al menos tres sitios de muestreo por cada intervalo altitudinal de 200 m de amplitud; al final, el total de unidades de muestreo fue de 17 en La Bufa y 15 en Laguna Juanacatlán. Los ejemplares no identificados en campo se colectaron para ser determinados y herborizados en el herbario IBUG. Esta metodología también ha sido usada en varios estudios de rangos altitudinales de la vegetación, estructura de BMM y bosques de Abies del occidente de México (Vázquez-García et al. 2000, Cuevas-Guzmán 2002, Sánchez-Rodríguez et al. 2003, Sahagún-Godínez 2004, Cuevas-Guzmán et al. 2011). Las variables edáficas se determinaron mediante análisis fisicoquímico de una muestra mixta de la capa superior del suelo (a una profundidad de 30 cm) por cada sitio de estudio (32 muestras en total). Se determinó la textura del suelo con hidrómetro de Bouyoucos, el pH con un potenciómetro Orion research 301 Analog Bench, la cantidad de materia orgánica de acuerdo al método de Walkley & Black (1934), la capacidad de intercambio de cationes (con el procedimiento de acetato de amonio), nitrógeno inorgánico, P, K, Ca, Na y Mg, todos con base en los métodos sugeridos en la NOM-021-SEMARNAT-2000 (SEMARNAT 2002).
Tabla 1 Características de los sitios de estudio a lo largo de dos gradientes altitudinales en el occidente de Jalisco, México
| Sitio | N Latitud | W Longitud | Alt (m snm) | Pend (°) | Exp (°) | Pa (mm) | T m anual (°) | PTS (mm) | PTF (mm) |
|---|---|---|---|---|---|---|---|---|---|
| La Bufa | |||||||||
| 1 | 20° 45' 15.6'' | 104° 50' 14.8'' | 1534 | 16 | 314 | 1256 | 18 | 29 | 64 |
| 2 | 20° 45' 10.4'' | 104° 50' 8.2'' | 1581 | 22 | 294 | 1256 | 18 | 29 | 64 |
| 3 | 20° 44' 59.6'' | 104° 49' 55.6'' | 1630 | 25 | 298 | 1228 | 17.1 | 30 | 73 |
| 4 | 20° 44' 59.9'' | 104° 49' 38.7'' | 1680 | 14 | 293 | 1228 | 17.1 | 30 | 73 |
| 5 | 20° 44' 53.6'' | 104° 49' 51.9'' | 1720 | 34 | 270 | 1228 | 16.6 | 30 | 73 |
| 6 | 20° 44' 55.7'' | 104° 49' 30'' | 1755 | 25 | 327 | 1228 | 16.6 | 29 | 73 |
| 7 | 20° 44' 52.7'' | 104° 49' 22.9'' | 1814 | 15.5 | 285 | 1215 | 15.1 | 29 | 79 |
| 8 | 20° 44' 47.2'' | 104° 49' 18'' | 1860 | 35 | 315 | 1215 | 15.1 | 29 | 79 |
| 9 | 20° 44' 42.2" | 104° 49' 13.1'' | 1937 | 19 | 315 | 1215 | 15.1 | 29 | 79 |
| 10 | 20° 44' 39.2'' | 104° 49' 11.6'' | 2020 | 33 | 315 | 1215 | 15.1 | 29 | 79 |
| 11 | 20° 44' 33.3'' | 104° 49' 9.1'' | 2108 | 29 | 315 | 1215 | 15.1 | 29 | 79 |
| 12 | 20° 44' 17.2'' | 104° 49' 35.2'' | 2170 | 32 | 270 | 1220 | 14.7 | 29 | 82 |
| 13 | 20° 43' 50.3'' | 104° 49' 47.6'' | 2265 | 35 | 358 | 1217 | 13.2 | 32 | 89 |
| 14 | 20° 44' 2.2'' | 104° 48' 52.4'' | 2282 | 20 | 10 | 1215 | 14.5 | 29 | 79 |
| 15 | 20° 43' 47.5'' | 104° 49' 43.1'' | 2345 | 31 | 341 | 1217 | 13.2 | 32 | 89 |
| 16 | 20° 43' 51.7'' | 104° 49' 38.9'' | 2374 | 37 | 315 | 1217 | 13.2 | 32 | 89 |
| 17 | 20° 43' 43.6'' | 104° 49' 37.7'' | 2447 | 29 | 329 | 1217 | 13.2 | 32 | 89 |
| Laguna Juanacatlán | |||||||||
| 1 | 20° 38 '24.1'' | 104° 46' 6'' | 1780 | 5 | 20 | 1217 | 16.5 | 29 | 72 |
| 2 | 20° 38' 22.1'' | 104° 45' 55.2'' | 1810 | 20 | 270 | 1215 | 16.3 | 29 | 73 |
| 3 | 20° 38' 1.6'' | 104° 45' 43.6'' | 1887 | 8 | 22 | 1215 | 16.3 | 29 | 73 |
| 4 | 20° 37' 50.5'' | 104° 45' 26.5'' | 1923 | 17 | 296 | 1215 | 16.1 | 29 | 74 |
| 5 | 20° 37' 30.5'' | 104° 44' 34'' | 1975 | 22 | 322 | 1211 | 15.7 | 28 | 75 |
| 6 | 20° 37' 15.5'' | 104° 43' 11.3'' | 2100 | 28 | 317 | 1200 | 15.1 | 29 | 77 |
| 7 | 20° 36' 45.8'' | 104°41'59'' | 2170 | 32 | 292 | 1188 | 14.3 | 30 | 80 |
| 8 | 20° 36' 58.9" | 104°41'42.3'' | 2256 | 31 | 297 | 1188 | 14.3 | 30 | 80 |
| 9 | 20° 36' 58.1 | 104°41'50.4'' | 2280 | 21 | 280 | 1188 | 14.3 | 30 | 80 |
| 10 | 20° 36' 57.2'' | 104°41'42.1'' | 2298 | 18 | 250 | 1188 | 14.3 | 30 | 80 |
| 11 | 20° 36' 33.2 | 104°42'5'' | 2353 | 20 | 280 | 1193 | 14.6 | 30 | 79 |
| 12 | 20° 37' 48.4'' | 104°42'2.2'' | 2360 | 28 | 280 | 1187 | 13.8 | 30 | 83 |
| 13 | 20° 37' 36.6 | 104°42'3.7'' | 2374 | 10 | 160 | 1187 | 13.8 | 30 | 83 |
| 14 | 20° 37' 48.1'' | 104°41'48.4'' | 2401 | 12 | 225 | 1183 | 13.9 | 30 | 81 |
| 15 | 20° 37' 46.9'' | 104°41'43.1'' | 2413 | 12 | 100 | 1183 | 13.9 | 30 | 81 |
Coordenadas geográficas, altitud (Alt), pendiente (Pend), exposición (Exp), precipitación anual (Pa), temperatura media anual (T m anual), precipitación del trimestre más seco del año (PTS) y precipitación del trimestre más frío del año (PTF).
Análisis de Datos. La diversidad de Shannon-Wiener y el estimador de riqueza ACE (Abundance Coverage Estimator) se calcularon con el programa EstimateS 9.0 (Colwell 2013). Después, se ajustaron ecuaciones lineales y polinomiales con las variables estructurales, diversidad Shannon y riqueza (variables dependientes) en relación con la altitud (variable independiente). Para evaluar las tendencias en casos con distribución de errores tipo Poisson (conteos de individuos y riqueza de especies) se utilizaron modelos lineales generalizados. El mejor modelo fue seleccionado con el criterio de información de Akaike corregido (AICc) (Burnham & Anderson 2002) utilizando el programa R project 2.5.1 (R Core Team 2007).
La información climática incluyó 19 variables bioclimáticas descargadas de la base de datos de WorldClim (Hijmans et al. 2005) para cada sitio de estudio. La precipitación del trimestre más seco del año y la precipitación del trimestre más frío del año fueron seleccionadas utilizando análisis de regresión y correlación de Pearson, para evitar colinealidad. Las variables bioclimáticas altamente correlacionadas fueron excluidas del análisis de ordenación. Solo dos variables bioclimáticas más (temperatura media anual y precipitación anual) fueron incluidas en este estudio, ya que las temperaturas y precipitaciones del resto de los trimestres estaban altamente correlacionadas.
Para analizar la relación entre la abundancia de especies leñosas y la ordenación de los sitios con las variables ambientales a lo largo del gradiente altitudinal, se utilizó el Análisis de Escalamiento Multidimensional No Métrico (en inglés Non-metric Multidimensional Scaling Analysis, NMDS) del programa PC-ORD 6 (McCune & Grace 2002) con 24 variables ambientales. La clasificación de los sitios de muestreo se realizó con un Análisis Jerárquico de Agrupamiento (Cluster Analysis), con una matriz que incluyó 32 sitios y 103 especies. Este análisis fue implementado utilizando la medida de distancia Sørensen (Bray-Curtis) y el método de ligamiento Beta Flexible (beta = -0.25). Las diferencias en la composición de especies entre los grupos obtenidos del Análisis de Agrupamiento fueron evaluadas estadísticamente mediante el Procedimiento de Permutación Multi-Respuesta (MRPP por sus siglas en inglés) (McCune & Grace 2002). La prueba estadística (T) describe la separación entre grupos (valores negativos reflejan separaciones mayores) y la probabilidad corregida dentro de un grupo (Δ). Cuando todas las especies dentro de un grupo son idénticas, Δ alcanza su valor mayor (Δ = 1). Cuando la heterogeneidad entre grupos iguala el nivel esperado al azar entonces (Δ = 0) y cuando existe mayor heterogeneidad dentro de los grupos que el nivel esperado al azar entonces (Δ < 0). En el Procedimiento de Permutación Multi-Respuesta el valor de p es dado para cada prueba de comparación grupal. Para identificar en forma objetiva y cuantitativa a las especies indicadoras de cada uno de los grupos del dendrograma del Análisis de Agrupamiento, se corrió el Análisis de Especies Indicadoras (ISA, por sus siglas en inglés) con el método de Tichý & Chytrý (2006). Estos análisis multivariados se corrieron usando la distancia Sorensen. El Análisis de Especies Indicadoras muestra las especies más exclusivas en cada uno de los grupos detectados en el análisis de agrupamiento, ya que utiliza un valor resultante del producto de la abundancia relativa y la frecuencia relativa de las especies en cada grupo seleccionado previamente. Este análisis arroja el valor indicativo máximo para cada grupo y la significancia estadística utilizando la técnica de Monte Carlo basado en 4,999 aleatorizaciones. Las pruebas estadísticas de ordenación NMDS, clasificación, MRPP e ISA fueron ejecutadas con el programa computacional PC-ORD 6.0 (McCune & Grace 2002).
Resultados
Composición florística leñosa y estructura de la vegetación. A lo largo de casi 1,000 m de intervalo altitudinal, en las dos localidades, fueron registrados un total de 3,801 individuos en 32 sitios (3.2 ha), los cuales pertenecen a 103 especies (incluyendo seis morfoespecies), 63 géneros y 42 familias (Apéndice 1). Las familias mejor representadas en número de especies fueron: Fagaceae (15), Pinaceae (8) y Asteraceae (6). Se resalta que Quercus (15) y Pinus (7) fueron los géneros con más riqueza, mientras que la mayoría de los géneros tuvieron una especie. Las especies más frecuentes (presentes en la mayoría de los sitios) fueron Abies jaliscana (23 sitios), Arbutus xalapensis (19), Quercus obtusata (16), Clethra hartwegii (12), Quercus castanea (12), Rumfordia floribunda (12), Styrax argenteus (12), Roldana angulifolia (11) y Ternstroemia lineata (11). Estas especies representaron, en conjunto, el 44.4 % del número total de individuos.
La estructura de la vegetación en el gradiente altitudinal de Laguna Juanacatlán mostró una tendencia lineal positiva en área basal, altura máxima y altura media (Tabla 3, Figura 2B, C y E); mientras que la densidad (N), índice de Shannon-Wiener (H') y la riqueza de especies (S), tuvieron un patrón unimodal con un pico en el intervalo de los 2,200-2,300 (N) y a 2,100 m (Hˈ y S) de altitud respectivamente (Figura 2A, D y F). En contraste, La Bufa no tuvo tendencias de cambio significativas en el índice de Shannon-Wiener y área basal (Figura 2C y D). Por otro lado, la densidad disminuyó significativamente con respecto a la altitud mientras que la altura media aumentó. La altura máxima tuvo un patrón unimodal con un pico en el intervalo de 1,900 a 2,100 msnm. (Figura 2E). Por último, la riqueza mostró una disminución monotónica en relación con el rango altitudinal (p = 0.0001) (Tabla 3, Figura 2F). Los resultados de los análisis fisicoquímicos del suelo se muestran en la Tabla 2.
Tabla 2 Valores resultantes de los análisis fisicoquímicos del suelo de los sitios de estudio a lo largo de dos gradientes altitudinales de bosque mesófilo de montaña y bosque de Abies en el occidente de Jalisco, México
| Sitio | M.O. (%) | C.I.C. (meq/100 g) | Textura | pH | N inorg. (ppm) | P (ppm) | K (meq/100 g) | Na (meq/100 g) | Ca (meq/100 g) | Mg (meq/100 g) |
|---|---|---|---|---|---|---|---|---|---|---|
| La Bufa | ||||||||||
| 1 | 12.8 | 21.3 | Areno Francoso | 5.3 | 48 | 4 | 1.3 | 1.5 | 12.5 | 2.9 |
| 2 | 22.4 | 60.3 | Franco arenoso | 5.8 | 126 | 0.5 | 1.0 | 0.0 | 13.6 | 4.3 |
| 3 | 22.4 | 16.7 | Franco arenoso | 7.6 | 47 | 1 | 1.5 | 1.5 | 9.0 | 2.8 |
| 4 | 6.7 | 41.5 | Francoso | 7.9 | 51 | 0.1 | 1.0 | 0.0 | 3.8 | 2.5 |
| 5 | 13.9 | 48 | Franco Arenoso | 7.6 | 211 | 1.2 | 1.5 | 0.2 | 25.4 | 5.3 |
| 6 | 17.9 | 41.9 | Francoso | 7.2 | 37 | 19 | 1.1 | 0.1 | 14.2 | 2.8 |
| 7 | 8.3 | 21.3 | Franco limoso | 6.9 | 37 | 2 | 0.8 | 0.6 | 9.6 | 4.4 |
| 8 | 16.9 | 66.8 | Franco Arenoso | 7.0 | 243 | 1.2 | 1.5 | 0.2 | 34.6 | 4.6 |
| 9 | 7.6 | 46.8 | Franco arenoso | 9.4 | 149 | 2.5 | 1.5 | 0.3 | 15.5 | 1.7 |
| 10 | 12.6 | 50.2 | Areno francoso | 6.7 | 121 | 2.6 | 0.8 | 0.2 | 17.0 | 2.1 |
| 11 | 19.9 | 51.4 | Areno francoso | 7.6 | 79 | 2.2 | 0.7 | 0.1 | 10.0 | 1.7 |
| 12 | 25.5 | 56 | Franco Arenoso | 7.2 | 210 | 25 | 1.4 | 0.2 | 29.7 | 2.7 |
| 13 | 14.7 | 42 | Franco arenoso | 6.7 | 79 | 2.4 | 1.3 | 0.2 | 16.5 | 2.4 |
| 14 | 19.3 | 67.4 | Franco Arenoso | 7.1 | 210 | 1 | 1.9 | 0.2 | 37.2 | 3.1 |
| 15 | 14.5 | 61.6 | Areno Francoso | 6.7 | 103 | 2.3 | 0.7 | 0.1 | 17.9 | 2.1 |
| 16 | 16.3 | 57.5 | Areno Francoso | 7.2 | 185 | 2.2 | 1.1 | 0.1 | 16.1 | 1.7 |
| 17 | 17.6 | 53.6 | Franco arenoso | 6.0 | 112 | 11.1 | 1.3 | 0.2 | 11.3 | 2.3 |
| Laguna Juanacatlán | ||||||||||
| 1 | 7.9 | 16.7 | Areno francoso | 7.2 | 75 | 3.8 | 0.3 | 0.1 | 5.8 | 0.8 |
| 2 | 2.1 | 18.1 | Franco arcillosoarenoso | 7.1 | 140 | 0.6 | 0.6 | 0.2 | 3.2 | 4.9 |
| 3 | 6.8 | 30.4 | Areno francoso | 7.6 | 84 | 2.6 | 2.2 | 0.1 | 3.9 | 1.2 |
| 4 | 4.1 | 18.1 | Franco arenoso | 7.6 | 70 | 7.2 | 0.5 | 0.0 | 7.2 | 1.0 |
| 5 | 25 | 23.3 | Areno francoso | 8.4 | 219 | 8.1 | 0.5 | 0.1 | 8.6 | 1.6 |
| 6 | 13.5 | 37 | Franco arenoso | 7.3 | 350 | 1.3 | 0.9 | 0.4 | 4.3 | 1.5 |
| 7 | 2.6 | 30.5 | Franco arenoso | 6.0 | 187 | 23 | 1.7 | 0.2 | 17.7 | 3.4 |
| 8 | 3.1 | 29.9 | Franco arenoso | 5.7 | 149 | 11 | 1.0 | 0.1 | 5.8 | 2.5 |
| 9 | 2 | 32.4 | Franco arcilloso | 6.4 | 145 | 0.5 | 1.1 | 0.3 | 10.7 | 2.7 |
| 10 | 7.5 | 44 | Franco arenoso | 8.1 | 154 | 1 | 1.1 | 0.2 | 11.2 | 1.6 |
| 11 | 13 | 70.3 | Franco arenoso | 6.8 | 126 | 1 | 1.3 | 0.2 | 37.6 | 4.0 |
| 12 | 23 | 73.7 | Franco arenoso | 6.9 | 149 | 1.4 | 1.3 | 0.0 | 42.3 | 3.7 |
| 13 | 4.1 | 29 | Franco | 5.7 | 149 | 0.8 | 1.7 | 0.3 | 7.7 | 1.6 |
| 14 | 15.7 | 55.8 | Franco arenoso | 6.8 | 173 | 0.8 | 1.0 | 0.4 | 12.3 | 1.8 |
| 15 | 15.3 | 49.9 | Areno francoso | 6.6 | 75 | 1 | 1 | 0.1 | 11 | 2 |
Abreviaturas: M.O. (materia orgánica), C.I.C. (capacidad de intercambio catiónico), pH (potencial de hidógeno, acidez), N inorg. (nitrógeno inorgánico), P (fósforo), K (potasio), Na (sodio), Ca (calcio), Mg (Magnesio), ppm (partes por millón), meq (miliequivalentes).
Tabla 3 Resultados del ajuste de modelos lineales (lm) y modelos lineales generalizados (glm) a las variables de estructura y diversidad de especies leñosas en 17 sitios de un gradiente altitudinal en La Bufa y 15 sitios en Laguna Juanacatlán, en función de la altitud. AICc = Criterio de Información Akaike corregido.
| Variables de estructura y diversidad | Modelo mínimo adecuado | AICc | Intercepto (a) | Pendiente (b) | Constante cuadrática (c) | r 2 o χ2 | Valor de p |
| LA BUFA | |||||||
| DENSIDAD | Lineal lm | 178.4 < 180.0 | 273.46 | -0.07925 | 0.29 | 0.0262* | |
| ÁREA BASAL | Lineal lm | 69.3 < 72.6 | 3.15 | 0.00077 | 0.02 | 0.5585 n.s. | |
| ALTURA MEDIA | Lineal lm | 81.8 < 84.4 | 1.14 | 0.00429 | 0.26 | 0.0367* | |
| ALTURA MÁXIMA | Polinomial cuadrático lm | 124.3 < 125.9 | 219.2 | 0.2479 | 0.000059 | 0.38 | 0.0341* |
| RIQUEZA (S) | Lineal glm (dist. Poisson) | 91.9 < 94.7 | 4.35 | -0.00081 | χ2 = 9.07 | 0.0001*** | |
| DIVERSIDAD (H´) | Lineal lm | 18.9 < 21.5 | 3.26 | -0.00057 | 0.2 | 0.0725 n.s. | |
| JUANACATLÁN | |||||||
| DENSIDAD | Polinomial cuadrático glm (dist. Poisson) | 274.9 < 331.2 | -23.2 | 0.02571 | -0.0000051 | χ2 = 168.5 | <0.0001*** |
| ÁREA BASAL | Lineal lm | 54.2 < 57.7 | -12.1 | 0.00812 | 0.72 | <0.0001*** | |
| ALTURA MEDIA | Lineal lm | 64.2 < 67.1 | -0.55 | 0.00518 | 0.35 | 0.02128* | |
| ALTURA MÁXIMA | Lineal lm | 118.9 < 122.1 | -46.1 | 0.0404 | 0.45 | 0.0058** | |
| RIQUEZA (S) | Polinomial cuadrático glm (dist. Poisson) | 78.7 < 81.2 | -23.1 | 0.024 | -0.0000055 | χ2 = 5.82 | 0.01714 * |
| DIVERSIDAD (H´) | Polinomial cuadrático lm | 4.4 < 14.4 | -29.4 | -0.0299 | -0.000007 | 0.6 | 0.0039** |
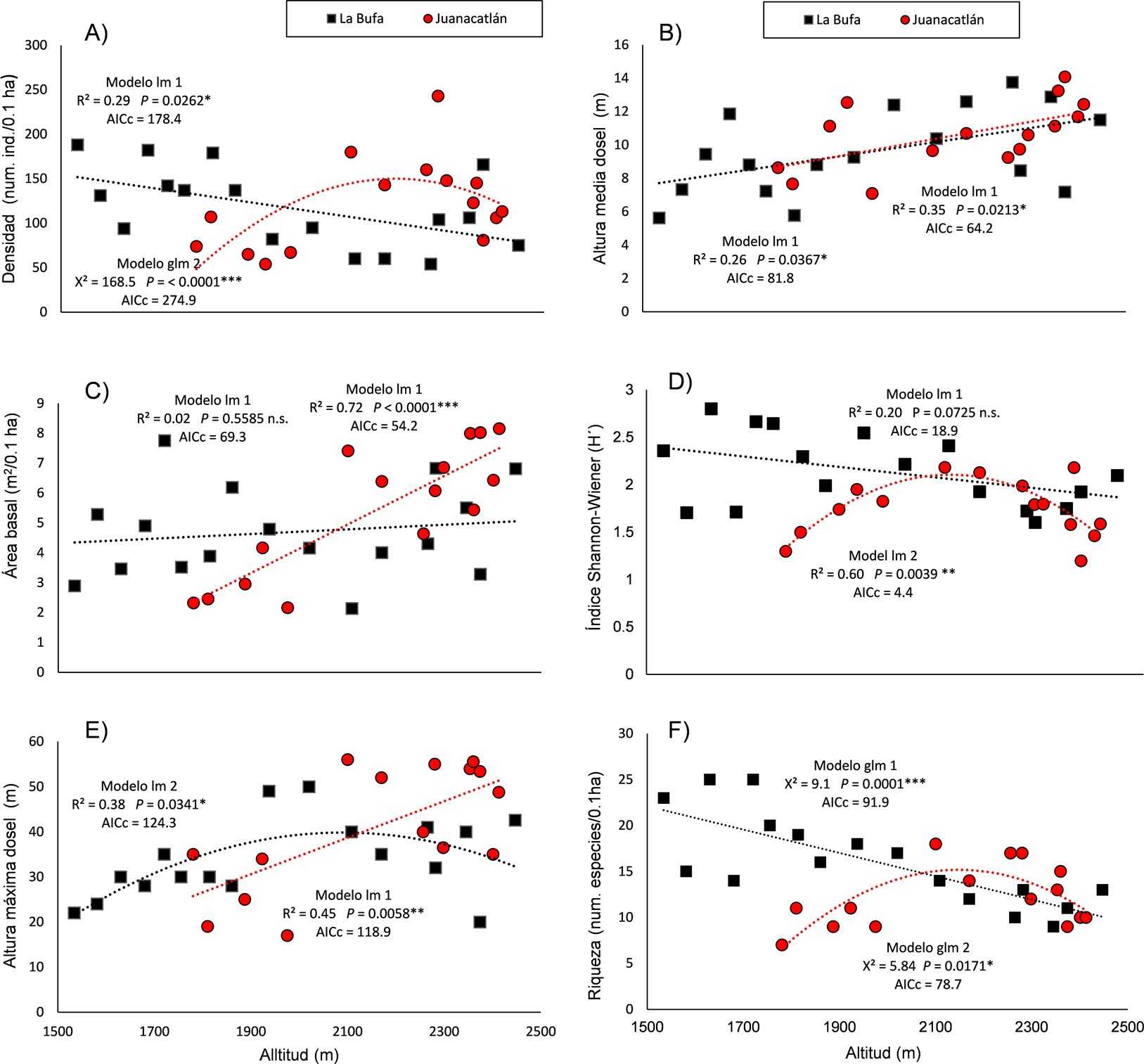
Figura 2 Modelos mínimos adecuados, ajustados a la densidad, área basal, altura promedio, altura máxima, riqueza observada y a los valores del índice de Shannon-Wiener (H') de plantas leñosas en 32 sitios de BMM-bosque de Abies en dos localidades del occidente de México; La Bufa (cuadros) y Laguna Juanacatlán (círculos), en función de la altitud. Modelo lineal (lm), modelo lineal generalizado (glm), modelo 1 (función lineal y = a + bx), modelo 2 (función Polinomial cuadrática y = a + bx + cx²). AICc (Criterio de Información Akaike corregido).
Ordenación, clasificación e interrelaciones clima-microambiente-vegetación. La ordenación indirecta NMDS de los 32 sitios (17 de La Bufa y 15 de Laguna Juanacatlán) con base en la abundancia de especies leñosas, resultó en una solución final de dos dimensiones o ejes (stress final: 16.04, 250 iteraciones) (Figura 3). La proporción de varianza acumulada fue r 2 = 0.65 (las proporciones de varianza representadas por el eje 1 y el eje 2, fueron 0.41 y 0.24 respectivamente). La prueba de Monte Carlo indicó que la probabilidad de que un estrés final similar pudiera haber sido obtenido al azar fue de p = 0.004.
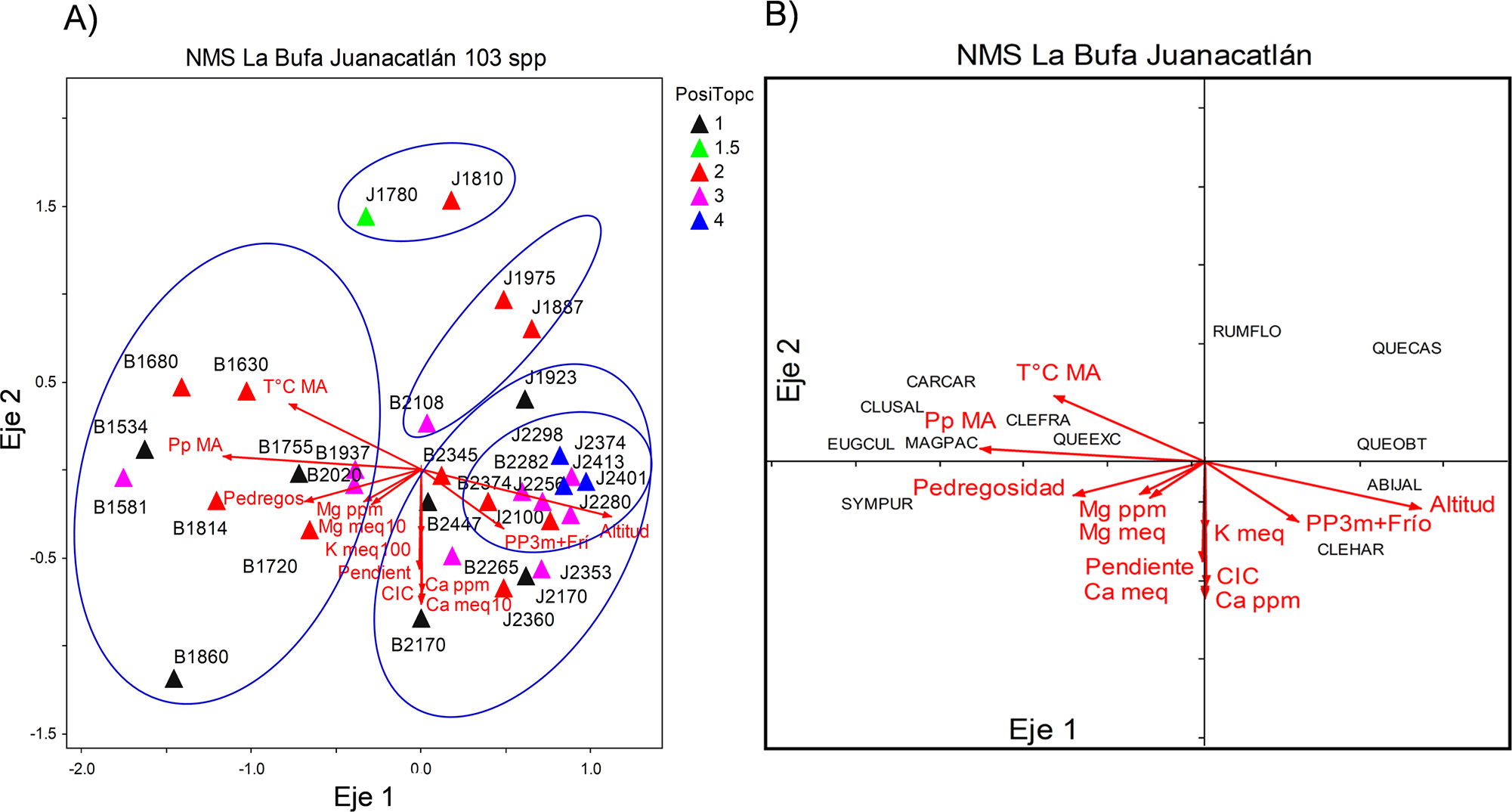
Figura 3 A) Análisis de Escalamiento No Métrico Multidimensional (NMDS) para los 32 sitios y 103 especies leñosas a lo largo del gradiente altitudinal en el occidente de Jalisco, en dos localidades, La Bufa y Laguna Juanacatlán; (T°C MA) Temperatura media anual, (Pp) Precipitación anual, (Pp3m+Frío) Precipitación del trimestre más frío del año, (Ca ppm) Contenido de Calcio en el suelo, (Mg ppm) Contenido de Magnesio en el suelo, (Ca meq) Catión de Calcio, (K meq) Catión de Potasio, (Mg meq) Catión de Magnesio. PosiTopo=Posición topográfica: (1) arroyo, (1.5) planicie baja, (2) ladera baja, (3) ladera media y (4) ladera alta. B) NMDS mostrando las 12 especies indicadoras, acrónimos: (ABIJAL) Abies jaliscana; (CARCAR) Carpinus caroliniana; (CLEFRA) Clethra fragans; (CLEHAR) Clethra hartwegii; (CLUSAL) Clusia salvinii; (EUGCUL) Eugenia culminicola; (MAGPAC) Magnolia pacifica; (QUECAS) Quercus castanea; (QUECEN) Quercus centenaria; (QUEOBT) Quercus obtusata; (RUMFLO) Rumfordia floribunda; (SYMPUR) Symplococarpon purpusii.
Las variables ambientales y climáticas que obtuvieron mayor relación con alguno de los dos ejes fueron, para el eje uno: altitud (r 2 = 0.64), precipitación anual (r 2 = 0.67), temperatura media anual (r 2 = 0.45), pedregosidad (r 2 = 0.39) y precipitación del trimestre más frío (r 2 = 0.28). Para el eje dos: pendiente topográfica (r 2 = 0.33), temperatura media anual (r 2 = 0.21), capacidad de intercambio catiónico del suelo (r 2 = 0.40), contenido de calcio del suelo (r 2 = 0.43), y de los cationes K (r 2 = 0.21) y Ca (r 2 = 0.44) (Figura 3).
El NMDS separó las parcelas localizadas en altitudes menores de aquellas localizadas en altitudes mayores a lo largo del eje 1 (Figuras 3A, B). Con valores más negativos para el eje 1 se encuentran los sitios de más baja altitud, los cuales están dominados por Carpinus caroliniana, Magnolia pacifica y Quercus centenaria; y con valores positivos se distribuyen los sitios de mayor altitud, dominados por Abies jaliscana, Clethra hartwegii y Q. obtusata. Además, a lo largo del eje 2 los ensambles fueron separados en: bosques dominados por encino-pino (sitios J1780, J1810, J1887, J1975 y B2108), bosques mixtos de alta montaña (sitios J1923, B2170, J2170, B2265, B2345, J2353, J2360 y B2447) y bosques dominados por Abies (sitios J2100, J2256, J2280, B2282, J2298, B2374, J2374, J2401 y J2413) inmersos entre los bosques mixtos de alta montaña. La presencia de cinco grupos de comunidades fue confirmada con su segregación a lo largo de rangos altitudinales, con un Análisis de Agrupamiento Jerárquico Aglomerativo (Hierarchical Agglomerative Cluster Analysis) (Figura 4). El Análisis de Agrupamiento resultó en un dendrograma en el que se distinguen los cinco grupos al realizar un corte vertical al nivel de una información restante del 35 %. El primer grupo incluye sitios de La Bufa (B) localizados a baja altitud (de los 1,534 m a los 2,020 m), el segundo grupo incluyó dos sitios de Juanacatlán (J) de baja altitud (de los 1,887 y 1,975 m) y un sitio de B (2,108 m de altitud); el tercer grupo incluye sitios de B de mayor altitud (entre 2,170 y 2,447 m) y entre 1,923 y 2,360 m en J; el cuarto grupo comprende a los dos sitios de J de más baja altitud (1,780 y 1,810 m); y el quinto grupo comprende a los sitios restantes de J de mayor altitud y a dos sitios de B de 2,282 y 2,374 m. Esta agrupación fue corroborada por el Procedimiento de Permutación Multirespuesta (MRPP), el cual proporcionó información máxima para hacer las separaciones de grupos, la homogeneidad dentro de los cinco grupos fue mayor que la esperada al azar (T = -12.59, A = 0.22, p = 0.00000).

Figura 4 Dendrograma resultante del análisis de agrupamiento de los sitios de estudio entre 1,534 y 2,447 m de altitud en bosque mesófilo de montaña y su transición con el bosque de Abies en el occidente de Jalisco, México. Se distinguen cinco grupos al realizar un corte vertical con una información restante del 35 %; La Bufa (B) y Laguna Juanacatlán (J).
El ISA identificó 12 especies como indicadoras significativas de los grupos (p < 0.05, Tabla 4). Para el grupo 1 fueron: Carpinus caroliniana, Clethra fragans, Clusia salvinii, Eugenia culminicola, Magnolia pacifica, Quercus centenaria y Symplococarpon purpusii; para el grupo 2 fueron: Quercus castanea y Rumfordia floribunda; para el grupo 3 fue solamente Clethra hartwegii; para el grupo 4 no hubo especies con valor indicador significativo; y para el grupo 5 las especies indicadoras fueron Abies jaliscana y Quercus obtusata. También resultaron otras 26 especies que tuvieron valor indicador máximo mayor a 0.50, pero que fueron marginalmente no significativos.
Tabla 4 Especies indicadoras de los cinco grupos determinadas por el Análisis de Especies Indicadoras (ISA)
| Especie | VIM | P | Especie | VIM | P |
| Grupo 1 | |||||
| Carpinus caroliniana | 0.74 | 0.032 | Grupo 2 | ||
| Clethra fragans | 0.87 | 0.000 | Pinus oocarpa | 0.55 | 0.074 |
| Clusia salvinii | 0.67 | 0.024 | Quercus castanea | 0.56 | 0.044 |
| Conostegia volcanilis | 0.51 | 0.071 | Rumfordia floribunda | 0.56 | 0.050 |
| Eugenia culminicola | 0.81 | 0.000 | Grupo 3 | ||
| Eugenia sp. | 0.59 | 0.058 | Clethra hartwegii | 0.56 | 0.001 |
| Euphorbia schlechtendalii | 0.51 | 0.076 | Meliosma dentata | 0.55 | 0.056 |
| Glossostipula concinna | 0.51 | 0.076 | Grupo 4 | ||
| Inga hintonii | 0.59 | 0.060 | Acaciella sp. | 0.67 | 0.059 |
| Magnolia pacifica | 0.74 | 0.032 | Clethra rosei | 0.67 | 0.062 |
| Miconia albicans | 0.41 | 0.510 | Quercus elliptica | 0.56 | 0.131 |
| Parathesis villosa | 0.51 | 0.072 | Quercus magnoliifolia | 0.56 | 0.134 |
| Picramnia guerrerensis | 0.51 | 0.076 | Quercus resinosa | 0.67 | 0.062 |
| Prunus cortapico | 0.51 | 0.080 | Quercus scytophylla | 0.60 | 0.118 |
| Quercus centenaria | 0.76 | 0.000 | Solanum brachystachys | 0.56 | 0.120 |
| Sommera grandis | 0.51 | 0.076 | Vaccinium stenophyllum | 0.667 | 0.0618 |
| Symplococarpon purpusii | 0.87 | 0.000 | Grupo 5 | ||
| Symplocos novogaliciana | 0.51 | 0.072 | Abies jaliscana | 0.34 | 0.045 |
| Synardisia sp. | 0.51 | 0.076 | Quercus obtusata | 0.56 | 0.001 |
| Verbesina fastigiata | 0.51 | 0.079 | |||
| Vitis sp. | 0.51 | 0.074 | |||
| Xylosma ciliatifolia | 0.52 | 0.074 | |||
| Xylosma flexuosa | 0.59 | 0.057 |
VIM= Valor Indicador Máximo
P = Valor de la probabilidad
De los cinco grupos discriminados por el análisis de agrupamiento se definieron cuatro tipos de bosque nombrados con base en dos criterios, el primero por sus principales especies dominantes y el segundo por el uso de la nomenclatura de Ohsawa (1991, 1995) para bosques de niebla (Tropical Montane Cloud Forests):
Bosque mesófilo de baja montaña (Lower montane cloud forest zone; grupo 1), 1,500-2,020 m snm. Este es el rango más amplio de bosque, aquí se mezclan especies perennifolias con caducifolias. Los elementos perennifolios como Magnolia pacifica, Symplococarpon purpusii, Inga hintonii, Clusia salvinii y Symplocos novogaliciana fueron encontrados en el estrato del dosel y subdosel. Las especies caducifolias por su parte fueron representadas principalmente por Carpinus caroliniana, Quercus centenaria, Quercus nixoniana y Ostrya virginiana. Las mayores alturas del dosel fueron dadas por Abies, Carpinus y Magnolia.
Bosque de Quercus-Pinus, 1,780-2,100 m snm. (grupos 2 y 4, parcelas J1,780, J1,810, J1,887, J 1,975, B2,108). Este es un fragmento del gradiente en el que normalmente debería haber BMM. Sin embargo, este bosque es dominado en el dosel por Quercus castanea, Q. coccolobifolia, Q. martinezii, Q. obtusata, Q. scytophylla y Pinus oocarpa. Entre los elementos mesófilos destacan Alnus acuminata, Oreopanax xalapensis, Ostrya virginiana, Persea hintonii y algunos individuos de Abies jaliscana. Aquí resaltan las especies heliófilas Baccharis salicifolia, Roldana angulifolia y Rumfordia floribundia con una alta abundancia.
Bosque mesófilo de alta montaña (Upper montane cloud forest zone; grupo 3 y una fracción del grupo 5), 1,920-2,450 m snm. Aquí el dosel superior es dominado ampliamente por Abies jaliscana, el cual alcanza alturas superiores a los 50 metros. A pesar de esto, existe un sotobosque bien desarrollado en diferentes estratos que está representado por los géneros Arbutus, Clethra, Cleyera, Ilex, Meliosma, Myrsine, Ostrya, Quercus, Symplocos, Styrax y Ternstroemia.
Bosque de Abies (Coniferous forest zone; una fracción del grupo 5, parcelas J2298, J2374, J2401, J2413). Este grupo fue exclusivamente dominado en todos los estratos por Abies jaliscana, aunque en el sotobosque pueden verse algunos individuos de tallas pequeñas de los géneros Pinus, Prunus, Quercus y Styrax. En esta franja altitudinal la mayor altura del estrato arbóreo fue de Abies, con 53 metros.
Discusión
En los bosques de las montañas tropicales y subtropicales de diferentes regiones del mundo se han encontrado patrones contrastantes en estructura de la vegetación, riqueza y composición florística a lo largo de gradientes altitudinales (Williams-Linera et al. 2013).
Composición florística leñosa en relación con la altitud. Las familias Fagaceae y Pinaceae fueron dominantes a lo largo de los dos gradientes altitudinales, especialmente en Laguna Juanacatlán, una tendencia similar a los bosques templados en la Sierra Nevada del Estado de México, Sierra de Manantlán y el centro de Veracruz (Sánchez-González & López-Mata 2003, Olvera-Vargas et al. 2010, Williams-Linera et al. 2013). La riqueza y diversidad de especies leñosas en Laguna Juanacatlán mostró una tendencia unimodal; es decir, aumenta al llegar a elevaciones medias y disminuye en las zonas más bajas y altas, con un pico en los 2,100 y una riqueza mínima en los 1,780 m de altitud. La tendencia unimodal también ha sido detectada en regiones templadas y tropicales (Tang & Ohsawa 1997, Sánchez-González & López-Mata 2003, Williams-Linera et al. 2013). En Cerro Grande, Sierra de Manantlán, aunque la diversidad de especies de plantas vasculares en general disminuye con la altitud, existe una tendencia unimodal en especies arbóreas, pero no es significativa (Vázquez-García & Givnish 1998). Por lo tanto, nuestros resultados sugieren que la precipitación invernal podría estar relacionada con el patrón unimodal en riqueza de especies leñosas, y estar contribuyendo a la presencia de bosque mesófilo de alta montaña y bosque de Abies; lo cual podría indicar mayor humedad proporcionada por niebla (no medida) en los primeros meses de la temporada seca, que en la zona de bosque de Quercus-Pinus con elementos mesófilos (1,780 m de altitud). En la Bufa hubo una disminución monotónica significativa en riqueza de especies, pero no significativa con el índice de Shannon. El patrón encontrado en La Bufa coincide con lo reportado por otros estudios, en los cuales fue observada una disminución de especies leñosas en las elevaciones más altas (Kitayama 1992, Lieberman et al. 1996, Aiba & Kitayama 1999, Behera & Kushwaha 2007, Homeier et al. 2010, Salas-Morales & Meave 2012). Otros autores han encontrado que la mayor riqueza de especies es explicada por variables relacionadas con alta precipitación y humedad en altitudes intermedias (López-Mata et al. 2012, Williams-Linera et al. 2013). Por el contrario, en la Sierra Madre del Sur, Oaxaca, la temperatura es la variable que explica, en su mayoría, el cambio de riqueza en relación con la altitud (Salas-Morales et al. 2015). La relación precipitación-riqueza de especies en bosques de niebla con coníferas también ha sido estudiada en El Himalaya, donde Abies y Tsuga dominan el rango de altitud más elevado y su presencia está positivamente asociada con el incremento de la precipitación anual, humedad relativa y la mayor riqueza de árboles (Wangda & Ohsawa 2010). Varios autores mencionan que un pico de diversidad en una altitud intermedia a lo largo de gradientes altitudinales podría ser el patron general (Lomolino 2001, Rahbek 2005), mientras que Williams-Linera et al. (2013) señalan que es importante considerar la escala cuando se comparan estudios, para entender los cambios de estructura y la riqueza de especies en el gradiente altitudinal.
Estructura de la vegetación en relación con la altitud. Los estudios sobre cambios estructurales en bosques húmedos a lo largo de gradientes altitudinales han generado resultados diversos. En investigaciones en el centro de Veracruz y La Chinantla, Oaxaca, no se encontraron cambios significativos en área basal y abundancia de árboles con el incremento de la altitud (Rincón 2007, Williams-Linera et al. 2013). En contraste, el gradiente altitudinal de Laguna Juanacatlán mostró incremento de área basal y abundancia en relación con la altitud, lo cual coincide con otras regiones estudiadas (Vázquez-García & Givnish 1998, Aiba & Kitayama 1999, Toledo-Garibaldi & Williams-Linera 2014). Por otro lado, La Bufa mostró una disminución significativa en abundancia a lo largo del gradiente altitudinal, mientras que en área basal no hubo un cambio significativo, esta última tendencia es similar a otras regiones que no exhiben una disminución o aumento significativo en términos de área basal, como el Monte Emei, China (Tang & Ohsawa 1997). La tendencia de área basal encontrada en La Bufa quizás se debe a que especies de grandes diámetros (mayores a 1 m), prosperan tanto en fajas altitudinales inferiores (Carpinus caroliniana y Magnolia pacifica) como en superiores (Abies jaliscana).
En distintas regiones del mundo se han encontrado varios patrones en la altura del dosel en relación con la altitud. Sin embargo, la tendencia mayormente reportada es que a mayor altitud hay una disminución de la altura máxima y media del dosel (Hamilton et al. 1995, Aiba & Kitayama 1999, Homeier et al. 2010, Krömer et al. 2013). En contraste, en ambos gradientes del presente estudio fue detectado un incremento significativo de la altura media del dosel a lo largo del gradiente altitudinal. El incremento en área basal y altura del dosel en Laguna Juanacatlán podría explicarse por la presencia de bosques mixtos y bosques casi monoespecíficos dominados por Abies jaliscana en altitudes mayores, los cuales presentan diámetros y alturas grandes. En general, la dominancia y presencia de las especies de oyamel o abeto está asociada con alta humedad y baja temperatura a lo largo del año (Rzedowski 1978, Ávila et al. 1994). En el rango altitudinal más elevado del Monte Emei en China, Abies fabri domina tanto el sotobosque como el dosel (Tang & Ohsawa 1997); sin embargo, este abeto no alcanza las tallas de los abetos mexicanos de la Faja Volcánica Trans-Mexicana, los cuales pueden alcanzar hasta 50 metros de altura (Cuevas-Guzmán et al. 2011, Guerrero-Hernández et al. 2014). La presencia de coníferas en zonas montañosas templado-frías de México-Centroamérica y el sureste de Asia, es un fenómeno que no ocurre con frecuencia en montañas tropicales, en las cuales se tiende a generalizar que la altura del dosel disminuye monotónicamente en relación con la altitud en los BMM (Hamilton et al. 1995, Scatena et al. 2010).
En el área de estudio, los sitios de mayor altitud mostraron menor precipitación anual que los de menor altitud (Tabla 1). Estos valores difieren de aquellos reportados en la estación climatológica de la Cumbre de Guadalupe (ubicada en la región noroeste de la Sierra de Cacoma, en la Sierra Madre del Sur, 50 km al sur de Juanacatlán), la cual muestra una precipitación anual de 2,003 mm a una altitud de 2,120 m (SMN 2016), en un lugar donde Abies jaliscana también forma comunidades vegetales casi monoespecíficas. Es posible que una menor evapotranspiración y una incidencia constante de niebla (variables no consideradas en WorldClim) son otros factores que podrían jugar un rol en mantener una humedad alta en los sitios en donde Abies es dominante, además de la disminución de la temperatura con el incremento de la altitud.
Ordenación, clasificación e interrelaciones clima-microambiente-vegetación. La altitud es la variable más fuerte en determinar tendencias en distribución y composición de especies leñosas en el BMM, debido a que modula otras variables tales como precipitación, temperatura y humedad. En este estudio, la ordenación NMDS reveló que la pedregosidad, la temperatura media anual más elevada y la mayor precipitación acumulada anual están asociadas con el bosque mesófilo de baja montaña (dominado por Carpinus caroliniana, Magnolia pacifica y Quercus centenaria). El bosque mesófilo de alta montaña y el bosque de Abies estuvieron positivamente asociados con la lluvia de invierno y el incremento altitudinal, lo que confirma la segunda hipótesis de este estudio. En contraste, Toledo-Garibaldi & Williams-Linera (2014) encontraron que en el oriente de México el BMM está asociado con la precipitación del trimestre más húmedo del año (PTH) mientras que el bosque de Abies con la altitud. Por otro lado, López-Mata et al. (2012) indicaron, para los bosques húmedos de montaña de México en general, una alta correlación entre riqueza, PTH, altitud, evapotranspiración anual, estacionalidad de la precipitación y carbono orgánico en el suelo.
La distribución de especies leñosas a lo largo de ambos transectos altitudinales apoya la hipótesis individualista de Gleason (1917, 1926), en la cual se postula que no hay grupos de especies bien definidos y con distribuciones similares, debido a que los centros y límites de la distribución de las especies están dispersos a lo largo de los gradientes altitudinales (Figura 5) (Whittaker 1975). Este enfoque fisonómico, permitió determinar que, para el caso del occidente de México, los límites entre bosque mesófilo de montaña y bosque de Abies no son abruptos, sino continuos, y se traslapan. Varios estudios se han enfocado en analizar solo el gradiente con muestreos en cañadas, crestas, cimas, elevaciones medias y tierras bajas, encontrando diferentes comunidades vegetales, comunidades discretas y discontinuidades (Vázquez-García & Givnish 1998, Ávila-Sánchez et al. 2010, Hemp 2010). Además, algunos autores han estudiado regiones grandes con condiciones climáticas y ambientales muy diferentes a lo largo de un amplio gradiente altitudinal, como en el centro de Veracruz (Williams-Linera et al. 2013, Toledo-Garibaldi & Williams-Linera 2014), lo cual explica los cambios un poco más abruptos y la alta heterogeneidad encontrada en los tipos de vegetación.
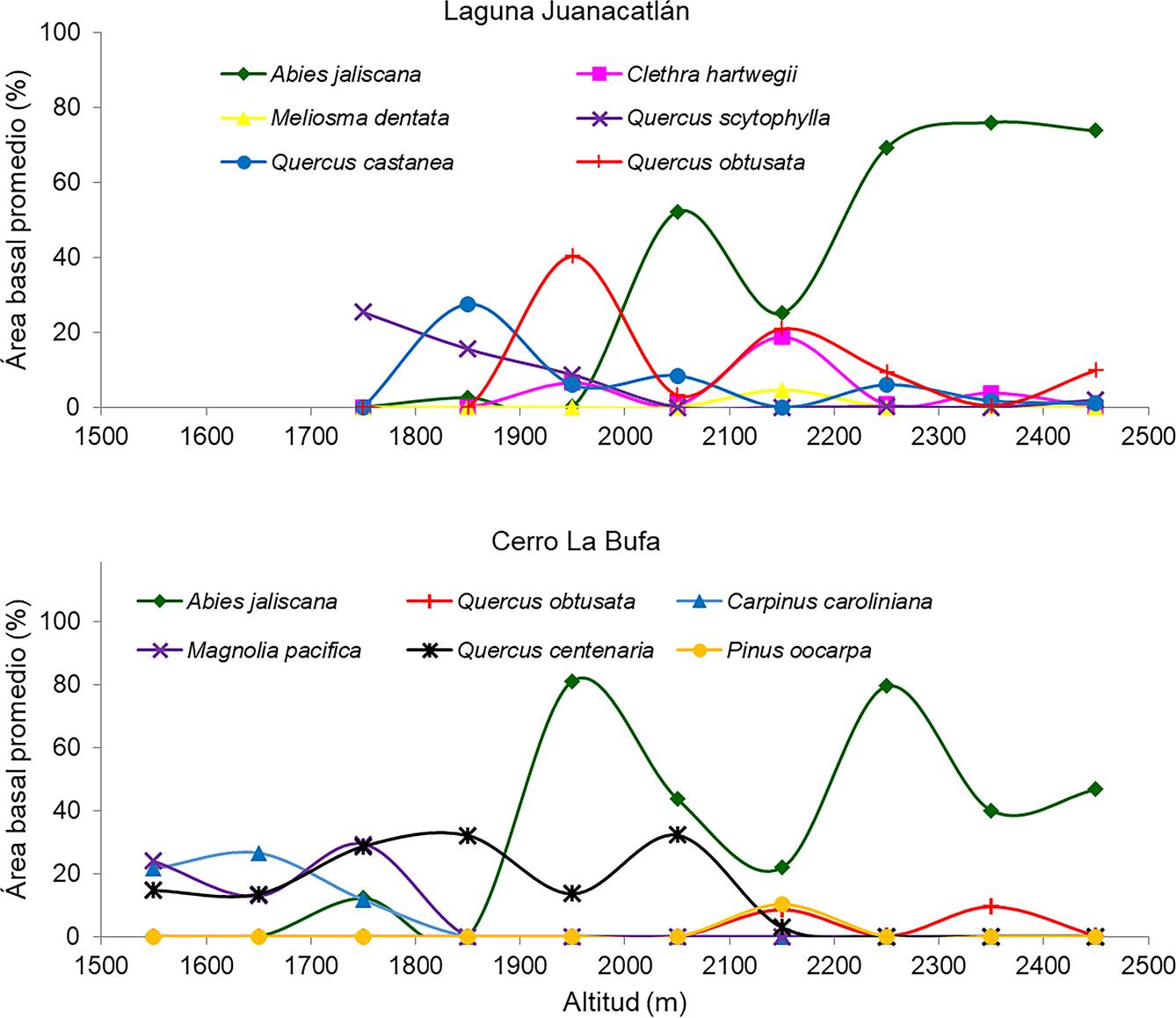
Figura 5 Especies dominantes en área basal, a lo largo de los dos gradientes altitudinales, La Bufa (B) y Laguna Juanacatlán (J).
Los sitios con menor pendiente y menor capacidad de intercambio catiónico en el suelo estuvieron dominados por especies relacionadas con ambientes menos húmedos o con la sucesión temprana, como es el caso de Rumfordia floribunda, Pinus oocarpa, Acaciella sp., Bocconia arborea, Clethra rosei, Quercus magnoliifolia, Quercus coccolobifolia y Quercus resinosa. Estas especies son componentes típicos del bosque de pino-encino y su presencia refleja la influencia tanto de un mayor disturbio antropogénico como de una menor humedad ambiental.
En resumen, a lo largo de los dos gradientes estudiados los patrones de riqueza, área basal y abundancia contrastan en ambas localidades; sin embargo, coinciden en que las mayores alturas de los individuos están dadas por Abies jaliscana. La ordenación y el análisis de agrupamiento, en este estudio, detectaron tres tipos de bosque que forman un continuo y son interpretados como bosque mesófilo de baja montaña, bosque mesófilo de alta montaña y bosque de Abies (desde los 1,500 a los 2,450 m de altitud). El género Abies es casi monodominante, en términos de área basal y abundancia, en cinco parcelas del rango altitudinal de 2,300 a 2,413 m. Los sitios con la menor altitud en Laguna Juanacatlán y un sitio de La Bufa formaron dos grupos distintos, los cuales fueron dominados por Quercus-Pinus. La distribución de estos grupos está ligada principalmente a las diferencias en altitud, precipitación anual e invernal, donde la niebla o precipitación horizontal es importante a considerar para futuros estudios; le siguen en importancia la pedregosidad, algunas variables edáficas y por último, el disturbio con la alta abundancia de especies heliófilas, como Rumfordia floribunda. El contraste de todos estos rasgos con otros bosques mesófilos de montaña de México-Centroamérica y del mundo, sugiere que para definir los cambios en estructura y composición de especies a lo largo de gradientes, se deben considerar todos los elementos con diferentes afinidades biogeográficas que forman zonas de transición o bosques mixtos; por ejemplo, la presencia de Abies y géneros típicos del bosque caducifolio templado como Carpinus, Cornus, Ostrya, Quercus y Tilia, que forman una transición entre el bosque mesófilo de montaña y el bosque de coníferas dominado por Abies.
Finalmente, los resultados indican un avance en la definición de los distintos ensambles en el bosque mesófilo de montaña. El bosque mesófilo de baja montaña, que incluye Carpinus, Conostegia, Inga, Magnolia y Symplococarpon es más diverso que el de alta montaña y el de Abies. El estudio muestra también que distintas especies dominantes están asociadas a diferentes condiciones climáticas y apoya dos tendencias: a) La riqueza de especies en La Bufa, disminuye linealmente con la altitud y se asocia con una mayor precipitación anual y mayor temperatura media anual; b) La riqueza de especies en Laguna Juanacatlán presenta un patrón unimodal, relacionado a una mayor precipitación en invierno; y en los sitios con mayor altitud a una menor temperatura. Estas tendencias apoyan de manera parcial la primera hipótesis de este estudio. La relación del bosque de Abies jaliscana con la precipitación del trimestre más frío (invierno), implica una alta fragilidad climática a la que se encuentra sujeto debido a las fluctuaciones climáticas actuales y pronosticadas a futuro, con posibles cambios en el régimen invernal (inviernos más secos).
En consecuencia, es necesario reforzar políticas de conservación de estos bosques, en especial en la vertiente del Océano Pacífico, donde también son muy heterogéneos (Vázquez-García et al. 2000, Sánchez-Rodríguez et al. 2003, Padilla-Velarde et al. 2008, Guerrero-Hernández et al. 2014), más escasos, discontinuos y relictuales, que los de la vertiente del Golfo de México (Vázquez-García et al. 1995, Vargas-Rodríguez et al. 2010, López-Mata et al. 2012). El realizar estudios acerca de la dinámica de comunidades contribuiría también a su adecuado manejo y conservación, ante la presión antropogénica actual.

