











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkPlant morphology is the expression of a set of characteristics in response to environmental conditions and therefore to the availability of resources. The shape of the plants is highly influenced by climatic and edaphic factors where they growth (Taiz & Zeiger 2006). These characteristics are important from the ecological point of view of the species and it is necessary to know them because they allow predicting their responses to climate change (Valverde et al. 2005).
It has been proven that environmental conditions such as temperature, relative humidity and altitudinal gradient influence the morphological characteristics of plants and their growth patterns (Viveros-Viveros et al. 2013). For example, for Turnera sidoides L. subsp. pinnatifida (Poir.) Arbo, Solís-Neffa (2010) conducted a study on the geographic patterns of morphological variation finding that morphotypes distributed over a wide geographical range occur in regions with different climatic regimes and deploy different habitat preferences. These authors showed that temperature and precipitation have an impact on these preferences, the largest leaf area in the Andean regions with greater precipitation and wider leaves in the warmer and wet areas; as well as the tendency to smaller leaves in cold and dry climates.
Water is one of the most influential factors in the plant morphology, its importance lies in the regulation of growth and its distribution (Folland et al. 2001). The annual precipitation patterns play a crucial role in the adaptation of the plants, in the strategies of water use and in the determination of the compositions of the vegetal communities. These patterns are related to plant phenotypes, soil characteristics and microclimates (Schwinning & Ehleringer 2001). For example, in Eucalyptus grandis W. Hill a positive and highly significant relationship was found between leaf area and greater water availability (Mejía-De Tafur et al. 2017). Although plant-water relationships have been an important part of ecophysiological research, the influence of variation in precipitation is poorly understood mainly due to the unpredictable nature of precipitation and the complexity of its function. Data on this topic have shown that the effects of rainfall variation on plant growth and ecosystem productivity can be complex and extensive and can differ widely between plants and ecosystems (Ogle & Reynolds 2004, Xu et al. 2007).
Light is another climatic factor that influence the growth and plant development, which plays a very important role, because the light received by plants influences the physiological and morphogenetic processes that determine the role of plants in the ecosystems (Taiz & Zeiger 2006). Particularly in arid and semi-arid ecosystems, the establishment of plants in open spaces with high availability of light is affected by water availability, for which plants can present morphological variation that allows them to adapt to these environments (Valladares et al. 2004). In semi-arid ecosystems, the plants limit the photosynthetic rate and its growth, presenting morphological and physiological adaptations to water deficit; these modifications include reduction of leaf area and productivity, deposition of waxes on leaves and stems, sunken stomata, abundant trichomes and, in some cases, the crassulacean acid metabolism (Passioura 1994, Lawlor & Tezara 2009).
Turnera diffusa Willd. is a small, deciduous, highly branched shrub, that measures between 30 and 200 cm of height; its leaves are lance-shaped with serrated edges and measure approximately 10 and 25 mm of length; it has small round fruits in the form of a capsule with a sweet smell and showy yellow flowers that appear during the summer (Alcaraz-Meléndez et al. 2007, Zhao et al. 2007, SEMARNAT 2008). It is supposed to originate in Brazil, but it is widely distributed in Mexico and Central America (Alcaraz-Meléndez & Véliz-Murillo 2006, Arbo & Espert 2009). In Mexico, it is located in Baja California, Baja California Sur, Campeche, Chiapas, Chihuahua, Coahuila, Colima, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, Quintana Roo, San Luis Potosí, Sinaloa, Sonora, Tamaulipas, Veracruz, Yucatán, and Zacatecas (Villaseñor 2016). It is generally associated with scrub, mainly xerophilous scrub and derived secondary vegetation, although there are also records in dry and humid forests, coastal dunes, and pastures, low forest and deciduous forest (Challenger & Soberón 2008). Turnera diffusa occurs in the arid and semi-arid zones of Tamaulipas, which are characterized by a hot climate, with an average annual precipitation of less than 900 mm, and usually the populations of this species are associated with thickets and thorny forest at altitudes ranging between 20 and 1,600 m a.s.l. (Holmgren et al. 1990).
There are two varieties of T. diffusa Willd, T. diffusa var. diffusa and T. diffusa var. aphrodisiaca (Ward) Urb. (Arbo 2000), which show differences in leaf size and trichome density (Baez-Parra et al. 2018). For Tamaulipas only, T. diffusa var. diffusa has been found (Arbo 2000).
The economic importance of T. diffusa lies in its industrial use as an infusion beverage, flavoring, liqueur ingredient and also has a medicinal use in traditional Mexican herbal medicine, since it is attributed to stimulating nervous, aphrodisiac and diuretic effects, mainly (Osuna-Leal & Meza-Sánchez 2000). Likewise, antimicrobial effects and possible hypoglycemic properties have also been reported (Zhao et al. 2007).
The morphological variability in cultivated and wild populations has been evaluated in different species, considering the number of branches and the height of the plant as main characteristics (Muñoz-Urías et al. 2008, Machahua 2010). It has been found that both wild and cultivated populations of T. diffusa present individuals between 30 and 200 cm of height (Alcaraz-Meléndez et al. 1994, Osuna-Leal & Meza-Sánchez 2000). However, despite its wide distribution, information of morphological parameters such as leaf area, flowering and seed production is null, which allows determining the morphological variability of the T. diffusa populations. It has been suggested that drought affects the growth and development of this species (Alcaraz-Meléndez & Véliz-Murillo 2006, Martínez de la Torre 2013).
We evaluated height, density of branches and leaf area of two populations of T. diffusa in the central zone of Tamaulipas, to address the hypothesis that water availability and temperature of the study sites affect growth of T. diffusa.
Materials and methods
Study area. The work was carried out in two sites in the central of Tamaulipas. Site 1 was in the municipality of Jaumave, 1 km south of the Padrón y Juárez “ejido” (23º 20 '33' 'N and 99º 25' 43 '' W), at an altitude of 930 m a.s.l. This site is characterized by a semi-dry semi-warm climate, with an average annual temperature of 20 °C and an average annual rainfall of 500 mm; the soil is of haplico xerosol type and predominant vegetation is submontane scrub; site 2, was located in the municipality of Güémez, located 4 km from the municipal seat in the “ejido” of the same name (23° 55' 58' 'N and 98° 58' 07 '' W) at 163 m a.s.l., presents a semi-warm subhumid temperate climate, with an average annual temperature of 24 ºC, an average annual precipitation of 700 mm, the predominant soil type is calcareous regosol and the vegetation present is grassland and submontane scrub with secondary vegetation (INEGI 2014).
Identification. Plants from both populations were identified by Dr. Arturo Mora Olivo, research scientist from Instituto de Ecología Aplicada of the Universidad Autónoma de Tamaulipas.
Climatic variables. The information of the climatic variables (temperature and precipitation) were obtained from the nearest meteorological station to each study site through the Comisión Nacional del Agua. With these data, the average annual temperature and accumulated annual precipitation, from September 2013 to August 2014, were calculated.
Sampling and soil analysis. A field sampling was done at the beginning of the work in October 2013 in each site. Ten collection points were randomly marked, at each point soil samples were taken from the first 20 cm, subsequently mixed and a composite, homogeneous and representative sample was obtained from each site. The soil samples were air dried, ground and sieved (2 mm), and stored in plastic bags until analysis in the laboratory. The pH, electrical conductivity, texture and nutrients of the soil (organic matter, organic carbon and nitrogen) were measured. The pH was determined by the potentiometric method (pH Meter 5500 Daigger) in a 2:1 ratio. The electrical conductivity was determined by means of a conductimeter (AB30 Accumet Basic, Fisher Scientific) on a sample of water or soil extract, based on the theory of electrolytic dissociation (Fernández-Linares et al. 2006), and the texture was determined by the size of soil particles, applying the Bouyucos method (Fernández-Linares et al. 2006). Organic matter, total nitrogen and total organic carbon were determined using the Walkley and Black method (Galantini et al. 1994).
Plant morphology. To determine the sample number, a preliminary sampling was carried out using the plant height as the response variable, which was used to define the sample of 15 transects, with a limit of the estimation error of less than 1.7 cm for Jaumave and 1.3 cm for Güémez. In both sites, the error limit was lesser than the standard deviation. Subsequently, random unrestricted sampling was carried out, applying the Gentry method (1988) to measure plant height, branch density, and leaf length and width in T. diffusa plants. The criterion used was to measure all the individuals of T. diffusa that were in the transect. In total, four samples were taken, two in each study site; in October 2013 and in April 2014. A total of 2,717 individuals considering all height classes were measured; 1,477 in Jaumave and 1,240 in Güémez.
Plant height and branch density. Plant height was measured from the ground level to the upper limit of the main branches, excluding the inflorescences. The branch density was determined as the number of branches per unit length of the stem. Both variables were determined according to Pérez-Harguindeguy et al. (2013).
Leaf area. All the 2,717 T. diffusa plants found in the evaluated transects in both sites, Jaumave and Güémez, were considered. A healthy single leaf of each individual was randomly chosen, and the length and width were measured to estimate leaf area. Leaf area was calculated as an ellipse according to De la Rosa-Manzano et al. (2014).
Exposure of plants to light. It was recorded if each plant was exposed to light or under the shade of a tree or shrub in the transects, in each study site.
Flowering. It was recorded if each plant of T. diffusa was in bloom in the transects, in each study site.
Statistical analysis. To compare the means of each morphological variable between populations, a one-way analysis of variance (ANOVA) was applied. In addition, a multivariate analysis with linear discriminant functions was performed, to determine the possible differences between populations studied considering the morphological variables (plant height, density of the branches and leaf area) as a whole. This analysis was used to assign cases to a group defined by a previous classification and recently, to evaluate a classification based on a certain number of attributes because this technique looks for discriminant functions that maximize the differences between populations. To measure the degree of linear relationship between precipitation and morphological variables (height, branch density and leaf area), a Pearson's simple correlation analysis was carried out for each month in which the measurements were carried out.
A chi-square test (χ2) was applied to evaluate flowering between study sites; as well to evaluate flowering in relation to the exposure of the plants in each study site. In addition, we considered three classes of plant height: 10-25 cm (seedlings), 26-40 cm (saplings and adults) and 41-95 cm (saplings and adults), and a chi-square test (χ2) was applied to evaluate the possible differences between the exposed plants and under shade for each height class in each study site. The statistical analyzes were done with the Statistica© 8.0 Program (Weiß 2007).
Results
Climatic variables of the sites. Annual rainfall and average annual temperature were lower in Jaumave (797 mm and 21 °C, respectively) than in Güémez (1681 mm and 25 °C, respectively). During the period evaluated (September 2013 - August 2014) in Jaumave, only two peaks of precipitation were recorded, in September 2013 (240 mm) and July-August 2014 (210 mm) and in Güémez were three very marked peaks, in September of 2013 (270 mm), January-March (220-230 mm), and July-August 2014 (215-210 mm). Regarding temperature, a decrease was observed in January 2014 (15 °C in Jaumave and 19 °C in Güémez) with a peak in July to August 2014, mainly in Güémez (30-33 °C) (Figure 1).
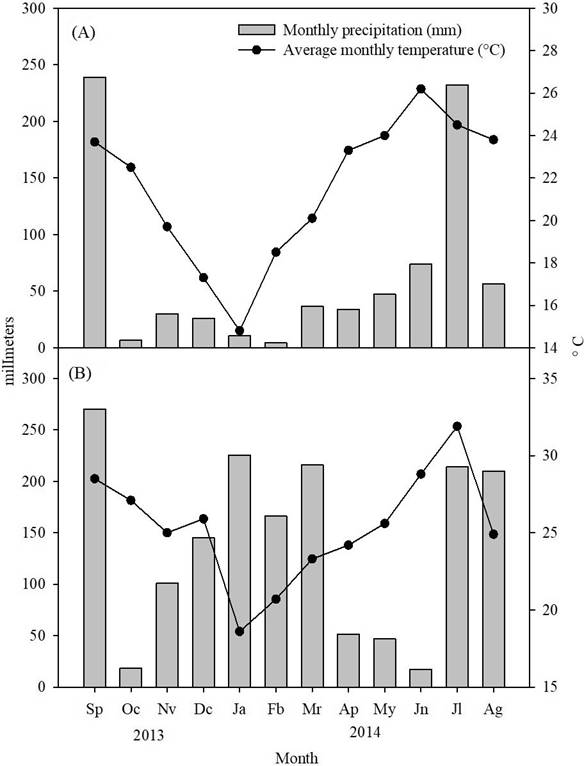
Figure 1 Register of monthly total precipitation and average monthly temperature in Jaumave (A) and Güémez (B), at Northeastern Mexico, during September 2013 to August 2014.
The texture, pH and electrical conductivity of the soil were very similar at the two sites; both soils showed characteristics of sediment trawl soils, not very permeable, moderately alkaline and not saline; however, nutrients, organic matter and carbon were higher in Jaumave compared to Güémez (Table 1).
Table 1 Physicochemical characteristics of the soil of Jaumave and Güémez, Tamaulipas.
| Soil physical-chemestry parameters | Jaumave | Güémez |
|---|---|---|
| pH | 8.1 | 8.24 |
| Texture | Silt loam | Sandy clay loam |
| Conductivity (µmhos/cm) | 0.95 | 1.52 |
| Organic Matter Content (%) | 9.56 | 6.89 |
| Organic Carbon (%) | 5.6 | 3.9 |
| Nitrogen (mg kg-1) | 0.32 | 0.39 |
Morphology of T. diffusa. We did not found differences in height between Jaumave and Güémez populations (24.5 ± 0.29 vs 24.1 ± 0.30 cm, respectively; F 1, 2715 = 0.8403, P > 0.01, Figure 2A). Branch density (F 1, 2715 = 4339.07, P < 0.01) and leaf area (F 1, 2715 = 685.68, P < 0.01) did show differences between populations, the values were higher in the plants of Güémez (13 ± 0.16 branches / plant and 5.59 ± 0.13 cm2, respectively) compared to those of Jaumave (3 ± 0.05 branches / plant and 2.34 ± 0.04 cm2, respectively; Figure 2B and C).

Figure 2 Morphological variables of Turnera diffusa in the populations of Jaumave and Güémez, at Northeastern Mexico. Each point indicates the mean ± standard error. Different letters between sites are statistically different means (P < 0.01).
We found significant differences in leaf length (F 1, 2715 = 652.7, P <0.01) and leaf width (F 1, 2715 = 705.4, P < 0.01) between Jaumave and Güémez populations. The longest and widest leaves were found in Güémez (1.81 ± 0.04 cm and 0.84 ± 0.02 cm, respectively), in contrast to those of Jaumave (1.22 ± 0.02 cm and 0.57 ± 0.01 cm, respectively; Figure 3A and B).

Figure 3 Leaf (A) width and (B) length of Turnera diffusa in the populations of Jaumave and Güémez, at Northeastern Mexico. Each point indicates the mean ± standard error. Different letters between sites are statistically different means (P < 0.01).
The analysis of linear discriminant functions showed that there are significant differences between the two populations of T. diffusa (Wilk's Lambda = 0.336, F 3, 2713 = 1,786.2, P < 0.01), branch density was the variable that contributed the greater variation and the relation of this variable was negative. The plant height contributed with the least variation between populations (Table 2).
Table 2 Analysis of linear discriminant functions of Turnera diffusa plants in two populations at Northeastern Mexico.
| Wilks' Lamb | Partial Lamb | F | P | Toler | 1-Toler | |
|---|---|---|---|---|---|---|
| Plant height | 0.3483 | 0.9694 | 98.669 | 0.0000 | 0.9402 | 0.0598 |
| Branch density | 0.7983 | 0.4210 | 3730.971 | 0.0000 | 0.9406 | 0.0593 |
| Leaf area | 0.3689 | 0.9111 | 264.651 | 0.0000 | 0.1695 | 0.0008 |
The Jaumave plants did present 2.4 % with characteristics associated with the plants of the Güémez population, whereas the Güémez plants did show 15.6 % with characteristics associated with the plants of the Jaumave population. However, 91.6 % of the total plants were classified according to the corresponding population (Table 3). Finally, the analysis of discriminant functions was validated, obtaining a Cohen Kappa value of 0.8291; therefore, it is considered a good model.
Table 3 Classification of Turnera diffusa plants according to functional linear discriminant analysis.
| % correctly classified | Jaumave population (density) | Güémez population (density) |
Total | |
|---|---|---|---|---|
| Jaumave population | 97.63033 | 1442 | 35 | 1,477 |
| Güémez population | 84.43549 | 194 | 1046 | 1,240 |
| Total | 91.60839 | 1636 | 1081 | 2,717 |
A strong linear association was found between precipitation and branch density (r = 0.85, P < 0.05) at each study site in October 2013, and moderate association (r = 0.72, P < 0.05) in April 2014. Precipitation and foliar area did show a moderate association in October 2013 (r = 0.44, P < 0.05) and April 2014 (r = 0.46, P <0.05), respectively (Table 4).
Table 4 Correlation coefficients of precipitation and morphological variables between the months in which the measurements were carried out.
| Sampling | Plant height r |
Branch density r |
Leaf área r |
|---|---|---|---|
| October 2013 | -0.03456 | 0.852347 | 0.440247 |
| April 2014 | 0.004106 | 0.724602 | 0.458145 |
Exposure of plants to light and flowering. The site was not related to flowering of T. diffusa (χ2 = 0.46, df = 1, n = 2,716, P = 0.49), nor with the shade condition (χ2 = 0.04, df = 1, n = 2,716, P = 0.83). The percentage of plants found in flowering was 63 % in Jaumave and 58 % in Güémez; and under shade was 56 % in Jaumave and 57 % in Güémez (Figure 4). There is a relation between the plants exposed and under shade with the plant height in the population of Jaumave (χ2 (1, N = 1403) = 83.72 P < 0.005), where the greatest number of plants was observed under shade in the strata of higher heights (26-40 and 41-95 cm) (χ2 (1, N = 1403) = 15.32 P < 0.005). This pattern is also observed in the population of Güémez, but it was not significant (χ2 (1, N = 1185) = 4.03 P > 0.005, Figure 5).
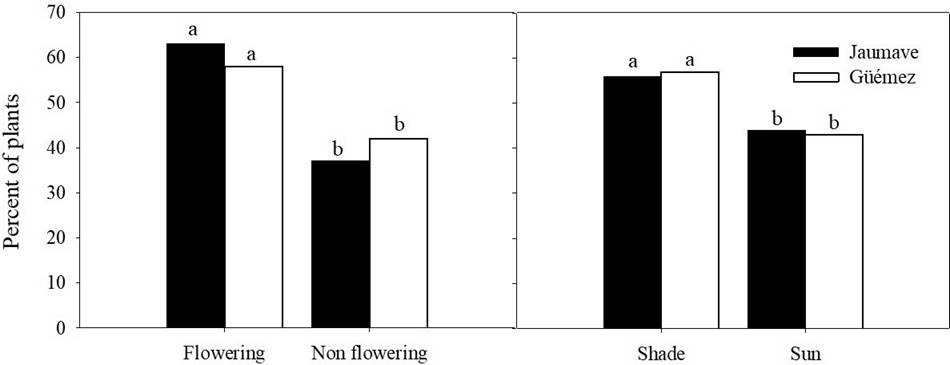
Figure 4 Relationship of the site with the flowering and the natural condition of sun and shade of Turnera diffusa plants with Jaumave and Güémez populations. Different letters indicate statistical differences (P < 0.01).

Discussion
The number of branches and the leaf area showed greater variability between the plants of Turnera diffusa of Jaumave and Güémez, the values of these variables were higher in Güémez. These differences are influenced by precipitation, since the monthly averages of precipitation were higher in Güémez compared to Jaumave, which was supported by the Pearson correlation analysis. It is widely known that water deficit is one of the main environmental factors that limit growth, photosynthesis and plant productivity (Shao et al. 2008). The number of branches was mainly responsible for the variation between populations, as in the Fabaceae Lotus tenuis Waldst. & Kit., where the number of branches and other morphological characteristics such as the length of the longest branch, the width / length ratio of the central leaflet and the dry weight of the aerial biomass, differed among the populations (Stoffella et al. 1998).
The reduction of the leaf area of T. diffusa at the site with the least rainfall (Jaumave) is influenced by the water deficit. Similarly, Solís-Neffa (2010) found in Turnera sidoides L. that the lower leaf area decreases under low water availability. It is suggested that in arid or semi-arid environments, such as the Ejido Padrón y Juarez in Jaumave, the reduction of the leaf surface is a mechanism of protection or tolerance adopted by the plants in order to avoid the loss of water by transpiration, in addition to other physiological responses such as the reduction of stomatal conductance, changes at the cellular and molecular level, root growth and changes in the assimilation of CO2 to reduce the photosynthetic rate (Medrano et al. 2007, Moreno 2009).
The analysis of discriminant functions showed significant differences for the height, the density of the branches and the leaf area. These results are similar to those found in Jatropha curcas L. in Perú, where the total height of the plant and the number of branches per plant among the different populations were significant differences, which was attributed to the variation in the environmental factors and the type of soil (Machahua 2010). Likewise, significant differences were registered between populations of J. curcas in India, with respect to the number of branches in relation to altitude, and for the effects of the edapho-climatic factors (Pant et al. 2006). Morphological variations have also been found among populations of Opuntia species (O. cantabrigiensis Lynch, O. leucotricha DC., O. rastrera F.A.C. Weber, O. robusta H.L. Wendl. ex Pfeiff. and O. streptacantha Lem.) in the southern Chihuahuan Desert; these species did show variation in the height of the stem and the number of spines (Muñoz-Urías et al. 2008). In other hand, Domínguez-Gómez et al. (2013) found little differences (1 cm) in the average height between two populations of T. diffusa with similar climatic characteristics in the arid region of Mexico, but the differences are only numerical because statistical analysis was not carried out. However, in other studies for different species it is suggested that morphological differences could be influenced by environmental conditions such as solar radiation and edaphic, as well as genetic factors (Silva et al. 2001, Hernández et al. 2007, Premoli et al. 2012).
The height of T. diffusa did show no significant differences between populations when was analyzed by itself; however, in the set of variables treated by the analysis of discriminant functions, it was significant. This analysis maximizes the differences between populations, and plant height was the variable that showed the least variation, so the probability that the differences of height of T. diffusa in Jaumave and Güémez is very low. The higher height of T. diffusa was found under natural shade conditions in Jaumave. The influence of light on the plant growth also has been found in T. trioniflora Sims plants that increased foliar area and decreased foliar thickness in response to shade (Kahar 2009). The species T. diffusa may need a plant that provides protection and offers specific microclimatic conditions for greater growth and development, as found for many arid and semiarid species (Steenbergh & Lowe 1969, Flores & Jurado 2003); for this reason, plants of this species are usually found under trees and between plants with thorns, in microenvironments formed under nurse plants (Alcaraz-Meléndez & Véliz-Murillo 2006). The relationship of height of the plant and exposure to light was not significant in the Güémez plants, this could be due that in this site the vegetation is denser and greater precipitation occurs (INEGI 2014).
Flowering is a condition that was not related to the exposure to light, similar to findings for Coffea arabica L. that did no show significant differences between the numbers of flowers considering shade treatments (Estívariz & Muschler 1997). However, it differs from other species, where different light conditions have a great influence on flowering, advancing or delaying it (Benvenuti et al. 1994, Valverde et al. 2009).
In conclusion, plants of T. diffusa from Jaumave and Güémez populations were morphologically different. Those of Jaumave had smaller number of branches and smaller leaves in comparison with the plants of Güémez; which was related to the greater precipitation that occur in Güémez in comparison with Jaumave. These differences are mainly marked by the density of the branches, which had a negative relationship with the height of the plants, so that, at a greater height of the plant, the density of the branches decreases.
These results contribute to understanding the growth plasticity of the useful species T. diffusa and could enhance the propagation of large numbers of cultivated individuals outside their habitats, promoting ex situ conservation. In addition, knowledge on the phenotypic plasticity of this species can help understand and predict their adaptive response to climate change and potential changes in plant distribution due to the loss of nurse plant species.

