











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkEcologists have searched for general trends among rare species that may reveal the underlying causes of rarity (Rabinowitz 1981, Gaston 1994, Gaston & Kunin 2012, Stebbins Jr 2013). Rare species have also been of great interest in the context of conservation biology, as many threatened species are naturally rare or have become rare as a result of anthropogenic processes (Gaston 2012). However, few common traits have been found among rare species (Esparza-Olguín 2004). Thus, rarity has long been an appealing but frustrating puzzle for ecologists and other biologists.
Among the features of a species’ biology that may determine its level of rarity are its reproductive traits, which affect its genetic diversity and structure, as well as important aspects of its population dynamics (Gaston1994, Esparza-Olguín 2004; Gong et al. 2015). In this study we address the reproductive biology of a rare plant, Furcraea parmentieri (Roezl) García-Mend (syn: Furcraea bendinghausii K.Koch), which is listed as threatened in the red list of the Mexican Environmental Agency (SEMARNAT 2010). Although F. parmentieri may be locally abundant, its geographic range is quite limited and it is found only in certain habitat types, which happen to have a high pressure of land-use change (Almeida-Leñero et al. 2014). Such features correspond to one of the types of rarity identified by Rabinowitz (1981) (i.e., locally abundant in a specific habitat type, but restricted geographically).
Furcraea parmentieri belongs to the Asparagaceae family, subfamily Agavoideae, a plant clade endemic to the Americas. Agavoideae consists of nine genera and ca. 300 species with a varied reproductive biology (Rocha et al. 2006). Plants in Agavoideae are either iteroparous (polycarpic) or semelparous (monocarpic); among the latter, rosettes produce a large reproductive stalk from their apical meristem and die after reproduction (though some may grow lateral offshoots) (Eguiarte et al. 2000, Rocha et al. 2006). The majority of species are self-compatible, and there is evidence of inbreeding depression in some genera (Eguiarte et al. 2000). However, there is self-incompatibility in the genus Hesperaloe (Pellmyr & Augenstein 1997) and in some Agave species that show pre-zygotic barriers. In such cases, as in other mostly hermaphroditic cross-pollinated groups (Sutherland 1987), fruit set is generally low.
The understanding of the reproductive biology of rare, vulnerable plant species is of paramount importance, given its central role in determining genetic diversity, which in turns shapes its evolutionary potential and affects its long-term persistence (Arizaga & Ezcurra2002, Newman et al. 2013). In hermaphroditic plants, fertilization may occur through either cross-pollination, which involves pollen transfer from the stamen of a flower in one plant to a different plant’s stigma; or self pollination, when pollen is transferred form a stamen to a stigma on the same plant, either within the same flower (autogamy) or to another flower on the same plant (geitonogamy) (González et al. 2016). Self-pollination may bring about relatively high fruit set with low reproductive costs; however, it does not translate in increased genetic diversity and may eventually lead to inbreeding depression (Shivanna 2016). On the other hand, cross-pollination may be associated with higher reproductive costs and lower fruit set, but represents an advantage in terms of increased genetic diversity and hence a higher potential to respond to the challenges of natural selection (González et al. 2016).
Although the Agavoidea subfamily is a well-studied group regarding reproductive biology (Eguiarte et al. 2000, Smith et al. 2012), very little is known about the genus Furcraea, despite it is one of its most diverse genera. It is known that most species in this genus are semelparous, but other details of their reproductive biology remain unclear. Interestingly, the two best studied genera within this sub-family (i.e., Agave and Yucca) show contrasting features in relation to their reproductive biology: Yucca species tend to be iteroparous while most Agave species are semelparous; Yucca species bear white flowers and present specialized cross-pollination system (McKain et al. 2016), whereas Agave may have different color flowers (from reddish to yellow, white, or greenish) and attract a wide array of pollinators including bees, wasps, hummingbirds, bats and moths, and are frequently self compatible (Rocha et al. 2006, Trejo-Salazar et al. 2015), which may result in high inbreeding depression (Eguiarte et al. 2000). Yucca flowers produce no or little nectar (5 to 8 µl per flower) with relatively low sugar concentration (ca. 11 %), while Agave flowers tend to have a much higher nectar production (up to 713 µl per flower, per night) with relatively higher sugar concentration (up to 26 %; Eguiarte et al. 2000, Rocha et al. 2006). Considering that the genus Furcraea is phylogenetically related to both Agave and Yucca, we hypothesize that the reproductive biology of F. parmentieri will share characteristics with both genera; however, we expect it to bear a closer resemblance to the Agave than to the Yucca group, as it is considered to be relatively closer to the former (Henández-Sandoval 1995, Good-Ávila et al. 2006).
The main aims of this study are: a) to describe the reproductive phenology of F. parmentieri at the population level and quantify the variation in flower morphology; b) to measure nectar production and identify the main flower visitors; c) to describe its reproductive system by evaluating fruit set and seed set under different pollination treatments; and d) to evaluate seed germination in progeny of different origins (geitonogamy, autogamy and cross pollination).
Materials and Methods
Study species. Furcraea parmentieri (synonym: Furcraea bendinghausii) belongs to the subfamily Agavoidea. This plant clade is endemic to the Americas and it has been suggested that Mexico is its centre of origin and diversification (Eguiarte et al. 2000). The genus Furcraea has ca. 25 species, most of which are native to Mexico (Rocha et al. 2006). Furcraea parmentieri belongs to the subgroup of furcraeas that occupy high altitude habitats (between 2,000 and 3,400 m; García-Mendoza 2000). This species is endemic to the Trans-Mexican Volcanic Belt (Figure 1), where it inhabits temperate forests at altitudes between 2,300 and 3,400 m), and establishes on soils of volcanic origin (García-Mendoza 2000). Given its limited geographic range and the high land-use change pressure to which its populations are subjected, this plant is listed as Threatened in the red list of the Mexican Environmental Agency (SEMARNAT 2010).

Figure 1 A) Mexico with its political division, showing the distribution of Furcraea parmentieri (black dots). The rectangle is the area amplified below. B) Central Mexico (area that corresponds with the rectangle marked in A), showing the distribution of F. parmentieri, as well as the different physiographic regions. Note that F. parmentieri is limited to high altitude areas. In light blue is the Trans-Mexican Volcanic Belt region; in pink is the South Sierra Madre; and in yellow is the Eastern Sierra Madre. The red cross indicates the study site.
Furcraea parmentieri is a long-lived, semelparous plant with arborescent, monopodial growth (longevity ca. 100 years, Hernández-Pedrero 2009). It has a simple trunk that may reach a height of 4 (-8) m, and 0.2 to 0.5 m in diameter (Figure 2). The trunk bears a large rosette on the top (2 – 2.5 m in diameter) with up to 150 fibrous erect lanceolate leaves (García-Mendoza 2000). Plants reproduce only once, after which they die. At reproduction, a large paniculate inflorescence emerges from the apical meristem between January and March. A fully developed inflorescence may measure up to 6 m in height and bears white campanulate flowers that remain open day and night between April and June (Figure 2). Fruits are capsules that ripen between October and December. Each fruit contains ca. 125 flat black seeds (9-11 × 6-8 mm), which are wind dispersed between January and February and germinate with the onset of the rainy season in June – July (Hernández-Pedrero & Valverde 2017). Thus, the complete reproductive period, from inflorescence production to seed germination, takes about a year and a half.

Figure 2 A) Adult individuals of Furcraea parmentieri at the study site. B) Setting up the pollination treatments on an inflorescence.
Furcraea parmentieri displays a mast seeding behavior (Herández-Pedrero 2009, García-Mendoza pers. com.). Most years only very few individuals reproduce, but there are years in which a large number of individuals synchronically start inflorescence production, giving place to a massive flowering event. There are no reports about the presence of a seed bank. Yet, our observations suggest that when seeds are dispersed and the rainy season begins, they either germinate or die, and do not accumulate in the soil.
Like many species in the Asparagaceae, F. parmentieri may produce bulbils in the inflorescence, i.e., vegetative structures that resemble small rosettes and may readily disperse and establish as vegetative propagules. It has been reported that in some F. parmentieti populations bulbil production is quite high, while capsule production is almost negligible, thus it is believed that population regeneration occurs mainly via vegetative propagation (García-Mendoza 2000). However, recent studies on population genetics confirm that seed germination and seedling establishment are common during its massive but occasional reproductive events (Bárceinas-Cruz 2011), although it is unknown whether fertilization depends on flower visitors or it may occur by wind or self-pollination.
Study site. This study was carried out at ‘El Xitle’ volcano, located in the south of Mexico City (19° 14’ 42’’ N, 99° 13’ 11’’ W), at an altitude of 2,800 – 3,100 m (Figure 1). The climate at this area is temperate, with summer rainfall (i.e., climate type Cb(w2)(w)ig, sensu García 2004). The average annual temperature is 14.7 °C. January is the coldest month (average temperature = 11.9 °C), and May is the warmest one (average temperature = 16.9 °C). Total annual rainfall is 910 mm, 92.2 % of which falls between May and October. The soil is derived from volcanic ash, with sandy and stony texture (Carrillo 1995). At this site, the F. parmentieri population covers a large area of steep slopes and relatively flatter land, where the vegetation is a pine-oak forests dominated by Pinus pseudostrobus, P. teocote, Quercus laurina, Q. rugosa and Arbutus xalapensis. The grass Muhlenbergia macroura dominates the understory, along with Verbesina virgata and some species in the genus Salvia (nomenclature follows Calderón de Rzedowski & Rzedowski 2005).
Field Methods. Phenology and flower morphology.- The study site was repeatedly visited from January 2007 to March 2008 in ca. fortnightly intervals (though more often during flowering and less often during fruit maturation). During this time we registered the different phenological events observed at the population level (presence vs. absence). With this information we constructed a yearly calendar in which we noted the reproductive structures present at any given moment.
Additionally, in March 2007 we tagged and followed 30 individual flower buds that were about to open (15 in each of two individuals) and visited them daily for 7 days to track their development. Observations were made between 16:00 and 21:00 hrs. For each flower, we measured daily the corolla aperture, and noted the stigma receptivity (humid stigmas were assumed to be receptive), and anther dehiscence.
To describe flower morphology, we collected six flowers in full anthesis from each of four individuals. Immediately after collection, we measured the diameter and height of the corolla (CD and CH in Figure 3), tepal length (TL), pistil and stamen length (PL and SL), flower length (FL), ovary length (OL), and internal diameter (ID) of the corolla (Figure 3). We obtained average measures for each individual and analyzed the variation between individuals through an ANOVA for each of the variables measured. We also carried out Spearman correlations to test whether there was any association between pairs of variables.

Figure 3 Flower traits measured in the flowers of Furcraea parmentieri. FL = flower length, OL = ovary length, SL = stamen length, PL = pistil length, TL = tepal length, CD = corolla diameter, ID = internal diameter, CH = corolla height.
Nectar production and flower visitors.- Each time we visited the studied F. parmentieri population during the flowering period (March to May 2007) we carried out informal observations of flower visitors. In addition, on the night of May 10 to the morning of May 11, 2007 we (3 observers) dedicated 4 hours to carefully record flower visitors in a total of 5 plants. Night observations were done from 21:00 to 22:00 and from 24:00 to 1:00 hrs, and morning observations were done from 7:00 to 8:00 and from 9:00 to 10:00 hrs. We recorded the frequency of visits in periods of 20 min per plant. Flower visitors could not be captured so their taxonomic identity could not be determined in detail.
In addition, on this same date we collected 12 flowers at midnight (3 flowers from each of 4 individuals) and another 12 flowers at 7:00 hrs next morning, and measured their nectar volume with a microcapillary pipette. The nectar sugar concentration was measured with a pocket refractometer (ERMA mod. 101190-32).
Pollination treatments.- Pollination treatments were set between April and May 2007. We selected 18 flowering F. parmentieri plants in the study population to establish 4 pollination treatments. Each treatment was applied to 5 flowers per individual; thus, there were 90 flowers per pollination treatment (18 × 5). When experimental flowers were selected, they were in the same developmental stage: they were barely open, with a humid, receptive stigma, and closed anthers. As described below, some treatments included flower bagging. To do so, we made small organdie bags, cutting circles of ca. 25 cm in diameter. These circles were stitched in their rims with thin copper wire and wrapped loosely around the experimental flower before softly tightening the wire at the base of the flower (Figure2 B). The four pollination treatments were as follows:
Cross pollination (CP) – Flowers were first emasculated (i.e., anthers were cut off) to prevent them from self pollinating, and then they were manually pollinated using a mix of pollen obtained from 15 flowering individuals located at the other extreme of the study population (ca. 500 to 1,000 m away). The pollen mix was placed on the stigma of the experimental flower making sure that it was well saturated, following which the flower was bagged as described above.
Geitonogamy (G) – Flowers were first emasculated, after which they were manually pollinated using a mix of pollen obtained from other flowers in the same plant. Flowers were bagged after pollination.
‘Autonomous’ pollination (AP) – This treatment consisted only of bagging the experimental flowers, without any previous manipulation. In this way, no potential pollinators could visit the flowers; thus, fruit set in this treatment would indicate that flowers can self-pollinate without the need of a pollen vector.
Open control (OC) – In this case we only selected and tagged flower buds on which no manipulation was done.
The study site was visited every two weeks from May to December to record fruit set in each of the 5 experimental flowers per treatment, per plant. As fruits ripened, they were collected and their seeds were counted. Seeds were separated in viable (i.e., of black color) and unviable (i.e., white); 4 groups of 20 viable seeds were weighted per fruit to obtain average seed weight.
The results of these experiments were evaluated in two moments in time: at fruit initiation (middle of May 2007), when flower ovaries started swelling suggesting that fertilization had occurred, and at the end of the fruit ripening season (end of November 2007), when fruits were collected and seeds counted. Thus, the response variables analyzed were i) the proportion of initiated fruits, ii) the proportion of fully ripened fruits (in both cases, in relation to the total number of flowers that conformed each treatment), iii) the number of viable seeds per fruit (black seeds were assumed to be viable), iv) seed set, which was calculated as the number of black seeds divided by the total number of seeds per fruit; and v) average seed weight of 20 seeds per fruit, per pollination treatment.
ANOVAs, followed by Tukey post hoc tests, were performed on these variables to test whether they differed between pollination treatments. As fruit-set and seed-set data are proportions, they were arcsin transformed for normality. Number of viable seeds per fruit was subjected to a log transformation.
Seed germination in progeny of different origins.- Seeds were obtained from the fruits collected in the pollination experiment described above. However, not all pollination treatments were included, as the ‘Autonomous’ pollination treatment did not produce any fruits.
Seeds were sown in Petri dishes with filter paper as substrate. Before sowing them, seed were disinfected by immersing them for 10 min in 5 % sodium hypochlorite. We sowed 4 Petri dishes, each with 20 seeds, for each pollination treatment, per experimental plant. The seed germination experiment consisted of a total of 60 Petri dishes and 1200 seeds (considering that not all treatments produced fruits in all plants). Petri dishes were placed in germination chambers with a constant temperature of 25±2 °C, ca.25 %relative humidity, and a 16:8 h photoperiod (hours of light: hours of dark). Seed germination was recorded daily for 15 days.
We obtained final germination percentage and the time required to reach 50 % germination (t50, as a means to evaluate germination velocity; Hatzig et al. 2015). Germination percentage was analyzed through a non-parametric Kruskal Wallis test in which the experimental factors were pollination treatment and parental plant. The variable t50 was analyzed through a two-way ANOVA with the same experimental factors.
Results
Phenology and flower morphology. The first signs of reproduction were observed in January and consisted in the start of the inflorescence stems on the apex of adult plants. By February the stem was noticeable in most reproductive plants and the developing of the inflorescence proper had started (Figure 4). The first open flowers were observed at the beginning of March and the flowering period lasted for three months, overlapping with fruit initiation in May. Thus, during the month of May, several phenological stages coexisted in the same inflorescence: flower buds, mature flowers (i.e., open, active), senile flowers and even initiating fruits. During the middle of May bulbil production started along inflorescence stems, and bulbils were present the rest of the year (Figure 4).
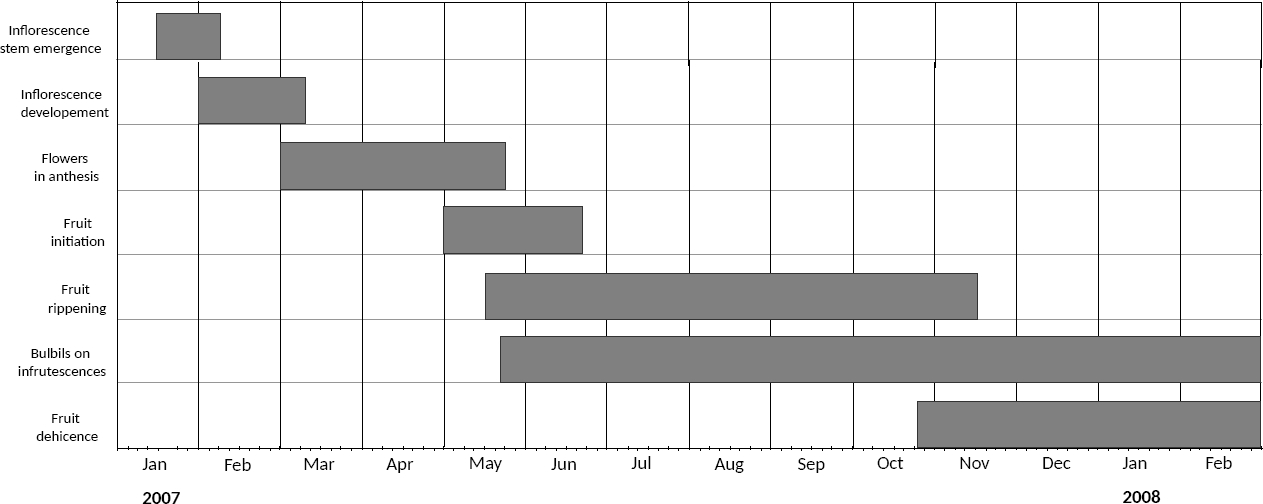
Figure 4 Reproductive phenology in the population of Furcraea parmentieri at El Xitle from January 2007 to February 2008. The grey bars mark the duration of each process.
Fruit ripening took several months, from May to November. The first dehiscent fruits were observed at the end of October, and by February of the following year most fruits had dispersed their seeds and plant die-out was apparent.
Furcraea parmentieri flowers were active for seven days and showed a slight protogyny: when the flower bud started to open, the pistil was receptive but anthers were closed. In the first day of activity of the flower, 41 % of anthers had opened and by the second this percentage increased to 97.2 % (Figure 5). During the second and the third day of the active flower, pistils were receptive and anthers were fully open, so in principle, self pollination could take place at least in terms of the timing of both processes. The corolla reaches its full aperture during the third day and by the fourth day pistils have started to lose their receptivity. On the seventh day, although the corolla is still slightly open, both pistils and anthers have dried out (Figure 5).
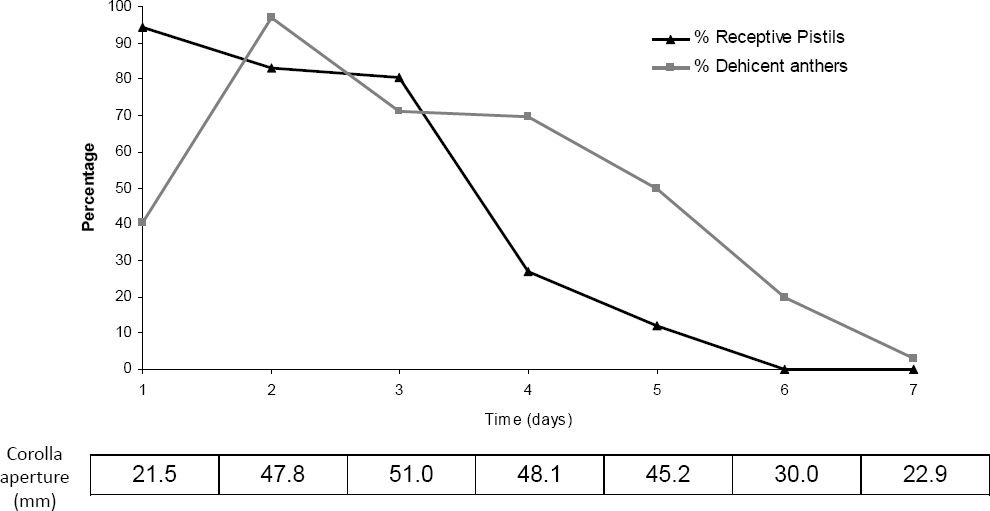
Figure 5 Percentage of receptive pistils and dehiscent anthers during the seven days of the life of a Furcraea parmentieri flower. Mean corolla aperture is shown below the graph for each day.
In relation to flower morphology, the only variables that did not differed significantly between individuals were the diameter and the height of the corolla, as most of the variation in these measures was observed within individuals. The rest of the flower measures differed significantly between individuals (Table 1).
Table 1 Mean and standard error (S.E.) of flower measures of several morphological variables obtained from 6 flowers from each of 4 individuals (total = 24 flowers) of Furcraea parmentieri. The P value shown in the right column refers to the significance level of the ANOVA performed to test the effect of parental plant. Initials in parenthesis next to each flower variable as in Figure 3.
| Flower morphological variables | Mean (mm) | S. E. (mm) | P |
|---|---|---|---|
| Corolla Diameter (CD) | 47.0 | 1.2 | 0.197 |
| Corolla height (CH) | 16.7 | 0.5 | 0.073 |
| Tepal length (TL) | 24.4 | 0.4 | 0.0002 |
| Pistil length (PL) | 18.1 | 0.2 | 0.0004 |
| Stamen length (SL) | 13.4 | 0.2 | 0.00003 |
| Flower length (FL) | 39.2 | 1.0 | 0.038 |
| Ovary length (OL) | 21.5 | 0.7 | 0.00002 |
| Internal diameter of the corolla (ID) | 12.8 | 0.1 | 0.0002 |
Several flower measures were correlated. The highest positive correlations were observed between ovary length and flower length (R= 0.793, P = 0.001), and internal flower diameter and tepal length (R= 0.672, P = 0.001). The highest negative correlations were found between corolla height and stamen length (R= -0.574, P = 0.003), and between internal flower diameter and stamen length (R= -0.540, P = 0.006). Pistil length and stamen length were the floral features that showed more significant correlations with other variables.
Nectar production and flower visitors. During the night observations the flower visitors were moths in the families Noctuidae, Geometridae, and Sphingidae. In the morning observations, two hummingbird species (Hylocarisleucotis andCynanthuslatirostris; identified by sight, comparing with the list of the local fauna), as well as bees, wasps and flies, visited F. parmentieri flowers (Table 2). In general, the frequency of flower visits was low, and in most cases the same individual visitor would approach the same group of flowers several times. The highest visit frequency was that of the hummingbirds, with almost 15 visits per plant in a period of 20 minutes. At night, the most active visitors were the Sphingidae moths (i.e., hawkmoths), with almost 6 visits per plant in 20 minutes (Table 2). Although it was impossible to make detailed observations about which flower parts were touched by these visitors, it was apparent that both hummingbirds and hawkmoths approached the flowers in a firm and unwavering manner and we consider that both groups may be effective pollinators.
Table 2 Flower visitors observed on 5 Furcraea parmentieri plants on May 10-11, 2007. Figures in the table are mean number of visits per 20 minutes, per plant. Insects were identified up to Family level. Hummingbirds belonged to two species: Hylocarisleucotis and Cynanthuslatirostris.
| Observation period (time of day) | ||||
|---|---|---|---|---|
| Flower visitors | 0700 – 0800 | 0900 – 1000 | 2100 – 2200 | 2400 – 0100 |
| Sphingidae (hawkmoths) | 0 | 0 | 5.8 | 0 |
| Geometridae (looper moths) | 0 | 0 | 0.8 | 1.0 |
| Noctuidae (owlet moths) | 0 | 0 | 0.2 | 0 |
| Apidae (bees) | 0 | 0.4 | 0.1 | 0 |
| Muscidae (flies) | 0 | 1.1 | 0 | 0 |
| Vespidae (wasps) | 0 | 0.1 | 0 | 0 |
| Hummingbirds | 14.9 | 1.7 | 0.8 | 0 |
| (Hylocarisleucotis and Cynanthuslatirostris) | ||||
From the 12 flowers collected at midnight, a total of 0.16µlofnectar were accumulated. The total amount of nectar accumulated during all the nigh in the 12 flowers collected in the early morning was over eight times higher (1.34 µl). The sugar concentration of the nectar was 26-28 %. Thus, F. parmentieri flowers show a very low nectar production, mainly during the night, and with a high in sugar concentration.
Pollination treatments. The cross pollination treatments resulted in a significantly higher proportion of initiated fruits compared to the other pollination treatments (F3,63 = 20.13; P < 0.0001;Figure 6), and also a significantly higher fruit set at the end of the experiment (F3,63 = 14.64; P < 0.0001;Figure 6). In the ‘autonomous’ pollination treatment only 1.1% of the fruits initiated, but fruit set was nil. The open control and the geinonogamy treatments had a similar percentage of initiated fruits (18-25 %) and a very similar fruit set (ca. 5 %).

Figure 6 Relative success of the different pollination treatments in terms of fruit set (proportion of fruits in relation to the number of flowers pollinated).
A total of 37 fruits were collected from experimental plants. Another group of seven fruits were collected at random from other plants in order to increase the sample size of the open control and present more robust results regarding seed set. In the end, we had 32 fruits originated from cross-pollination, 3 fruits produced by geitonogamy and 9 fruits from the open controls. There were no significant differences in the total number of seeds per fruit according to fruit origin (F2,41 = 0.27, P = 0.762), or parental plant (F7,22 = 1.76, P = 0.145). The average number of seeds per fruit was 207.5.
Pollination treatments did not have a significant effect on seed set (number of viable seeds divided by the total number of seeds per fruit), or seed weight (F 2,41 = 0.80; P = 0.457; and F 2,57 = 0.18, P = 0.83, respectively). Seed set ranged between 58 to 78 % (average = 67.7 %), and seed weight was, on average, 0.245 g per group of 20 seeds. However, the parental plant did have a significant effect on seed weight (F8,51 = 139.18, P <0.001). Some plants produced heavier seeds than others. The plant with the lightest seeds was, on average, 0.195 g per group of 20 seeds, while the plant with heaviest seeds was 0.335 g per group of 20 seeds.
Seed germination in progeny of different origins. Seed germination reached 98 %, on average. In 80 % of the Petri dishes seed germination was 100 %. The Kruskal-Wallis test showed that seed germination did not vary according to the pollination treatment from which they were originated (H(2,N=15) = 5.55, P = 0.06), nor to the parental plant (H(8,N=15) = 7.38, P = 0.5). In relation to germination velocity, we observed a significant effect of the pollination treatment (F2,57 = 3.75, P = 0.029), with seeds from the CP and G treatments reaching 50 % germination in ca. 6 days, and those from OC in ca. 7.4 days. The effect of parental plant was also significant on this variable (F8,51 = 30.83, P < 0.0001); some plants had rapidly germinating seeds (t50 = 4.5 days), while others took twice as long to reach 50 % germination.
Discussion
The flowers of Furcraea parmentieri are white, which is usually associated with nocturnal pollinators. However, nocturnal Lepidoptera (moths) and Chiroptera (bats) are attracted by strong flower odors (Rosas-Guerrero et al. 2014), while F. parmentieri flowers have a soft citrus-sweet scent (pers. obs.). This may explain the low abundance of nocturnal visitors on its flowers. In addition, we detected an extremely low nectar production (0.16-1.34 µl of nectarin a total of 12 flowers; nectar production being mainly nocturnal) with a relatively high concentration of sugars (26-28 %). A moderate nectar production with high concentration of sugars is usually associated with a pollination syndrome by hymenoptera (Perret et al. 2001) which may benefit from the nocturnal nectar production if they visit flowers early in the morning. Yet, only 2.7 % of diurnal visits were hymenoptera.
Unexpectedly, the most abundant visitors were hummingbirds (82 % of the recorded visits), which are usually attracted to tubular, bright-colored flowers (Johnson& Nicholson 2008). However, the floral display of F. parmentieri during mast seeding years is massive: there may be up to ca. 50 inflorescences per hectare, each reaching up to 10-12 m in height, and the mean number of flowers per inflorescence is ca. 17,500. Thus, F. parmentieri’s flowers may result attractive for hummingbirds merely because of its conspicuousness, and also because of the lack of alternative food resources for them during its flowering season. Whether they are effective pollinators is yet to be confirmed through more thorough observations. However, our data on flower morphology (i.e., flower, pistil and stamen length) and reports on the beak length of the two hummingbird species recorded (Arizmendi & Ornelas 1990,Zamudio Beltrán2010) suggest that they could be effective pollination vectors of F. parmentieri flowers, as they are likely to make contact with the different flower parts while trying to harvest the nectar. This is also the case for the few hawkmoths observed. However, it has been shown that hawkmoths do not rely exclusively on specialized plants as nectar sources, which may explain their low visitation rate to F. parmentieri flowers (Amorim et al. 2014). Clearly, further investigation on this area is needed in order to verify whether hummingbirds and hawkmoths actually transport pollen from one flower to another, and whether they do so between flowers of different individuals, as cross-pollination seems to be crucial.
Fruit set was almost identical between the open control and the geitonogamy treatment, which suggests that pollen transport occurs predominantly between flowers in the same inflorescence. Flower fertilization definitely depends on pollen transport by a vector, as the ‘autonomous’ pollination treatment had no success whatsoever. Nevertheless, whichever this vector is, it is not particularly effective or active in the study population. According to our observations, F. parmentieri flowers are not massively visited, albeit their conspicuousness; these observations are supported by the low fruit set recorded in the ‘open control’ (ca. 5 %), and in the population as a whole.
Although the pollination syndrome of F. parmentieri suggests a rather specific association with nocturnal insects, the actual relationship between its flowers and their pollinators must be of a generalist type, given the long and unpredictable periods between massive reproductive events. Most of the flower traits measured showed significant variation between individuals, thus, they are not genetically fixed perhaps due to diffuse selective pressures from different pollinator groups. On the other hand, the current flower traits may have evolved driven by interactions with flower visitors that have gone locally extinct; or else, they may be the result of phylogenetic restrictions (Herrera 1996,Silva-Montellano 2001).
The low fruit set found in the ‘open control’ supports the idea that there is a reproductive limitation in F. parmentieri due to the scarcity or low effectiveness of pollinators. Low natural fruit-set values have been recorded in other related species (Hesperaloe sp. with 0.5-4 %, Agave palmeri with 3.7 %, A. chrysantha with 5 %; Slauson2000, Rocha et al. 2006). However, in other cases higher fruit-set values have been found, such as in Yucca sp. (6-21 %; Eguiarte et al. 2000, Rentsch & Leebens-Mack 2014), Agave xylonacantha (63 %; Rocha et al. 2005), A. lechuguilla (40-50 %; Silva-Montellano 2001), A. cupreata (30 %; García-Meneses 2004), as well as A. celsii var. albicans (44 %), A. difformis (47 %) and A. striata (51 %) (Rocha et al. 2006). In F. parmentieri fruit set increased to 43 % when cross-fertilization was done manually. This implies that the genetic quality of the pollen reaching the stigma has a strong effect on fruit set. As in other members of the subfamily Agavoidea, there is self-incompatibility in F. parmentieri; although it has also been suggested that the low success of auto-pollination could be an early expression of inbreeding depression (Husband & Schemske 1996). Yet, later in the development of fruits and seeds no signs of inbreeding depression were found: the number of seeds per fruit, seed set, seed weight and seed germination were similar in all pollination treatments. Seed viability was high, as with other species in the same genus (e.g., 95 % in F. cahum, 92-100 % in F. longaeva, and 100 % in F. pubescens; García-Mendoza 2001, de la Rosa 2010). There were differences in average seed weight, but these were associated with the parental plant, and not with the pollination treatment. Thus, they could result either from genetic variation between plants, or from differences in resource availability between reproductive plants.
We estimate that the average number of seeds that a F. parmentieri individual may produce is ca. 21,000 (from the number of fruits produced by N = 22 plants), which is not such a large number compared with other Agavoidea species: Arizaga et al. (2000b) estimated that an Agave macroacantha plant produces ca. 20,000 seeds, while Gentry (1972) stated that an A. chrysoglossa individual produces ca. 750,000. In relation to seed set, F. parmentieri showed a relatively high value (63 %) compared to other Agavoidea species: 43 % in A. cupreata, 35 % in A. celsii var. albicans, 37 % in A. difformis, 31 % in A. striata, and ca. 60 % in A. macroacantha (Arizaga et al. 2000a, García-Meneses 2004, Rocha et al. 2005).
Furcraea parmentieri has an interesting life history. Plants are long lived, semelparous, and show massive reproductive events that are synchronous at the population and supra-population level (species-wide synchrony, sensu Keeley & Bond 1999). Despite the conspicuousness of reproduction, fruit set is relatively low because of high self-incompatibility and a low flower visitation rate, as is the case with other rare species (e.g., Gong et al. 2015). As stated before, we consider F. parmentieri as rare despite the fact that it may be locally abundant, because of its high habitat specificity and its limited geographic range. Although the relatively high local abundance and the massive reproduction should theoretically favor a high cross-pollination rate, in this case potential pollinators seem to be scarce, either because of the high altitude of the study site, or because of the proximity of the heavily urbanized Mexico City, which results in the low fruit set observed.
It is likely that the few pollinators that do visit reproductive F. parmentieri plants, predominantly transfer pollen between flowers in the same inflorescence (i.e., geitonogamy). However, reproductive stalks produce bulbils, in addition to flowers and fruits, which is not uncommon within the genus Furcraea (García-Mendoza 2000). Thus, even if sexual reproduction fails or turns out to be negligible, reproductive events may be followed by a high establishment of new individuals of sexual and/or vegetative origin. Bárceinas-Cruz (2011) studied the population genetics of this species using ISSR’s and found high levels of genetic variability both within and between populations. This suggests that at least some propagules of sexual origin – and derived from cross-fertilization – successfully establish within populations. The latter is supported by the results of Hernández-Pedrero & Valverde (2017) who studied seedling emergence and survival and found them to be high compared to other Agavoidea species (seedling emergence = 20-50 %; seedling survival after one year = 14 %). Thus, F. parmentieri populations appear to be healthy and thriving both demographically and genetically, despite the relatively low fruit set found.
As mentioned before, compared to the other Agavoidea species whose reproductive biology has been studied in detail, the genus Furcraea is phylogenetically closer to the Agave than to the Yucca group (Good-Ávila et al. 2006); thus, we had hypothesized that it would show a closer resemblance to the former in relation to its reproductive biology. The results of our study show that F. parmentieri shares some traits with Yucca (e.g., white flowers with a soft scent; strict self incompatibility), other traits with Agave (e.g., a generalist association with its flower visitors; semelparity; large and conspicuous inflorescences), while other traits are not shared with either (e.g., an extremely low nectar production, but with a high sugar concentration). Thus, regarding its reproductive biology, we cannot conclude that F. parmentieri shows a higher resemblance to one group compared to the other.
The geographic distribution of F. parmentieri is limited to relatively high altitudes in the trans-Mexican volcanic belt (Figure 1). At present, the remaining populations are restricted to the higher areas of this mountain range and are separated from one another by large expanses of urban or agricultural land. Habitat fragmentation, which is occurring at a high rate, accounts for the high genetic differentiation between populations (Bárceinas 2011). The environmental alterations anticipated due to climate change (Still et al. 2015), along with the high land-use change pressure on what remains of its natural habitat, will probably lead to the loss of the colonizable habitat of F. parmentieri within the present century. Therefore, it has been classified as threatened by the Mexican Environmental Agency (SEMARNAT 2010), and we believe this category barely does justice to the species’ situation. We encourage conservation planers to strengthen the policies of protected areas and to use this species in restoration and reforestation programs to ensure its future persistence.

