











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIn arid and semi-arid areas where weather conditions are hard for cultivating pasture, grazing depends exclusively on the shrub production (Ramírez et al. 2008). Some advantages to use shrubs as forage for animals (von Maydel, 1996) are that, green leaves have high crude protein content throughout most of the year (Ramírez-Orduña 2003); they constitute habitat for wildlife and coverage to prevent soil erosion (González-Rodríguez & Cantú-Silva 2001).
The Tamaulipan Thornscrub is a vegetation of the semiarid zone from the coastal plains of the Gulf of Mexico to the southern shoreline of Texas, U.S.A. (Foroughbakhch et al. 2005, 2009). This ecosystem presents a high species richness and diversity of tree and shrub species (Heiseke & Foroughbakhch 1985, Alanís-Rodríguez et al. 2008, Jiménez-Pérez et al. 2013, Mora-Donjuán et al. 2013). Many of them are important in forestry and silvipastoral production (timber, posts, firewood, forage, etc.), which constitute the most common land use in the region (Foroughbakhch et al. 2009). During the last 350 years, extensive grazing has been practiced in these areas (Jiménez-Pérez et al. 2012), resulting in the loss of both quality and quantity of forage plant species followed by a reduction in the plant layer which covers and protects the soil (Eviner & Chapin 2003, Fahring 2003, González et al. 2009). This leads to decrease the vegetable production soil capacity, resulting in an abandonment of the area for the natural regeneration of vegetation (Pequeño-Ledezma 2013), which is a significant shortfall for the people who depend on this resource. That scenario can be reversed through a management plan involving livestock, forestry and agroforestry practices, in which foliar biomass production of the shrubland is an important parameter to consider. According to Ledesma et al. (2010), the foliar biomass is a fundamental variable in the tree structure, since it is directly related to primary productivity. The evaluation of this variable is affected by the tree size and requires more time and resources on big trees (Rodríguez-Ortiz et al. 2012).
However, plant production in most environments is limited by the scarcity of mineral nutrients (Dominguez-Gómez et al. 2013). Under natural conditions, the amount of nutrients available is always limited and external additions are scarce, so the plants need to recycle, reduce losses and maximize efficiency in the use of nutrients for maximum biomass production (Escudero & Mediavilla 2003). This information is key to properly develop conservation, restoration and sustainable management plans (Legendre et al. 2005), where the litterfall can be a fundamental tool.
The litterfall is plant material accumulating on the ground, and its decomposition plays an important role in maintaining the productivity of forest ecosystems, affecting the transfer of organic matter, carbon, nitrogen, phosphorus and other nutrients of vegetation to the soil (Pérez-Suárez et al. 2009). It is the main nutrients source of forest soil (Landsberg & Gower 1997, Vitousek & Sanford 1986) and represents about 80 % of nutrients returned to the soil by the detritus of the tree (Santa Regina et al. 2001). The quantification of litterfall accumulation and its nature are crucial to understanding nutrient cycling in forest ecosystems (Zapata et al. 2007). However, litterfall production and accumulation are influenced by climatic variables (temperature and precipitation), soil conditions and anthropogenic interventions (Pavón et al. 2005, Prause et al. 2003, Roig et al. 2005, Zaldívar-Jiménez et al. 2004), which make difficult its determination.
There is a tendency to relate the litterfall accumulation and biomass production. Therefore, the objectives of this study were to quantify the leaf litter fluctuations through the seasons of the year and the leaf biomass production, and determine the relationship between fallen leaves pattern and foliar biomass production in forest plantations of five species of the Tamaulipan thornscrub, in the northeastern Mexico, as a tool for an adequate management of this ecosystem.
Materials and methods
Study area. The study was conducted in the school shrub plantations of the Forest Sciences Faculty of the Autonomous University of Nuevo Leon (UANL) in the town of Linares, located within the coordinates 24° 47’ north latitude and 99° 32’ west longitude in a plain region between 430-450 m altitudes in the foothills of the Sierra Madre Oriental, Mexico (Foroughbakhch et al. 2014). The regional climate is defined as semi-arid and sub-humid [(A) C (Wo)] in the scheme of Köppen modified by García (2004), with two rainy seasons (summer and autumn) and a dry season between November and April. Mean annual temperature is 22.3 ºC with temperatures over 40 ºC during the summer, and frost from December to March. Mean annual precipitation is 749 mm (Foroughbakhch et al. 2001), with the precipitation ratio to free evaporation of 0.48 and precipitation to potential evaporation of 0.62 (Foroughbakhch et al. 2009).
Selection of the species. The five woody species with the higher values of ecological characteristics (Alanís-Rodríguez et al. 2008, Jiménez-Pérez et al. 2009) were selected for this investigation: Acacia berlandieri, Acacia wrightii, Ebenopsis ebano, Havardia pallens and Helietta parvifolia. The choice was made after careful consideration of their growth rate, potential uses and preferences by local populations (Table 1).
Table 1 Uses, forest measurements and ecological variables of five timber species of the Tamaulipan thorn scrub in the northeastern Mexican semiarid land.
| Family | Species | Uses | BD (cm) |
DBH (cm) |
Ht (m) |
C (m2/ind.) |
RA (%) |
RF (%) |
IV (%) |
|---|---|---|---|---|---|---|---|---|---|
| Mimosaceae | Acacia berlandieri Benth. | Handcrafts, Firewood, forage | 3.10 | 2.05 | 3.25 | 8.53 | 8.4 | 4.1 | 5.36 |
| Mimosaceae | Acacia wrightii Benth | Firewood, Charcoal, Handcrafts, Food (seeds), wood, forage. | 7.60 | 4.40 | 3.70 | 5.64 | 2.3 | 5.6 | 5.8 |
| Mimosaceae | Ebenopsis ebano (Berland.) Barneby & J.W.Grimes | Posts, Firewood, Charcoal, Wood (furniture), shade for animal protection, Food (fruit & pods). | 7.90 | 5.95 | 4.80 | 7.60 | 6.1 | 6.6 | 7.60 |
| Mimosaceae | Havardia pallens Britton & Rose. | Firewood, construction and manufacture of furniture. | 5.20 | 3.98 | 4.80 | 6.72 | 3.6 | 5.6 | 3.73 |
| Rutaceae | Helietta parvifolia (A.Gray ex Hemsl.) Benth. | Poles, shelves, wood, Charcoal, medicinal. | 4.40 | 3.05 | 5.00 | 6.78 | 8.7 | 8.2 | 8.80 |
BD. Basal Diameter; DBH. Diameter at breast height; Ht. Height; C. Coverage; ind. Individual; RA. Relative abuncance; RF. Relative frequency; IV. Importance value. Sources. Foroughbakhch et al. 2009, Velazco-Macías et al. 2011, Martínez-Hernández et al. 2013, Ngangyo et al. 2014a,b.
Sampling design. Three plots of 100 m2 (10 × 10 m) for each woody species containing 25 trees were installed on a 30 years old experimental plantations. The plantations were not subjected to any type of silvicultural treatment or use.
Foliar biomass estimation. The foliar biomass was determined by “Adelaide” method or “hand reference” method (Forouhbakhch et al. 1996), which is the most precise, practical and simplest method for measuring the forage leaf biomass of many shrub species (Forouhbakhch et al. 2009). The method consisted in selecting outside the study area, a branch called “reference unit”, as representative unit in form and foliar density, for each studied species. Then, the number of the branch units was estimated by counting on the four cardinal points of the crown of 10 trees per plot, resulting in a total of 150 shrub samples. Afterwards, the reference units were brought to the laboratory to separate the leaves and dry them in an oven at 65 °C to constant weight. The dry foliar biomass of each species was obtained by multiplied the number of reference units by the dry weight. The evaluation was conducted by seasons, from June 2013 to May 2014, in order to determine the seasonal production during a year.
Litterfall accumulation. The methodology used was proposed by Návar-Cháidez & Jurado-Ybarra (2009), which consisted of collecting fallen leaves by litter traps of 1 m2 (1 × 1 m), with mesh of 1 mm2 opening. Three traps per plot were randomly placed under the trees, and the collection was made every 15 days during the year (from May 2013 to April 2014). The litterfall was manually separated into leaves, branches, twigs, seeds and flowers; the leaves fraction was used for this study. The dry mass of the collected leaf litter was determined on a digital balance (Ohaus Corporation, model EP612C, Pine Brook, NJ, USA; 0.001 g of precision), after dried in an oven (Lab-Line, Instruments, Inc., model 3476, Imperial V, Melrose Park, IL, USA) at 70 °C for 72 hours. The annual accumulation of leaf litter was determined by summing the production of all samples taken, per replication and studied species.
Statistical analyses. To verify statistical differences in the foliar biomass production and leaf litter accumulation, a multifactor analysis of variance was applied (Quinn & Keough 2002), and the interactions of each of the two parameters were studied with species and seasons through simple regressions. The statistical analyzes of all data was performed using the statistical program Statgraphics Centurion XVI (STATGRAPHICS 2002). Relationships were adjusted to the averages of foliar biomass and leaf litter, through simple regressions and corresponding equations, by species and seasons.
Results
Foliar biomass production. The foliar biomass production presented significant differences (P < 0.05) among species as well as seasons (Table 2). The higher annual average production was registered by Ebenopsis ebano with 2687 kg ha-1 whereas the lower annual average was recorded by Acacia wrightii with 431 kg ha-1 (Figure 1).
Table 2 ANOVA for Foliar biomass by Season
| Source | Sum of Squares | Df | Mean Square | F-Ratio | P-Value |
|---|---|---|---|---|---|
| Between groups | 1,06625E7 | 3 | 3,55415E6 | 7,06 | 0,0004 |
| Within groups | 2,82085E7 | 56 | 503723, | ||
| Total (Corr.) | 3,8871E7 | 59 |
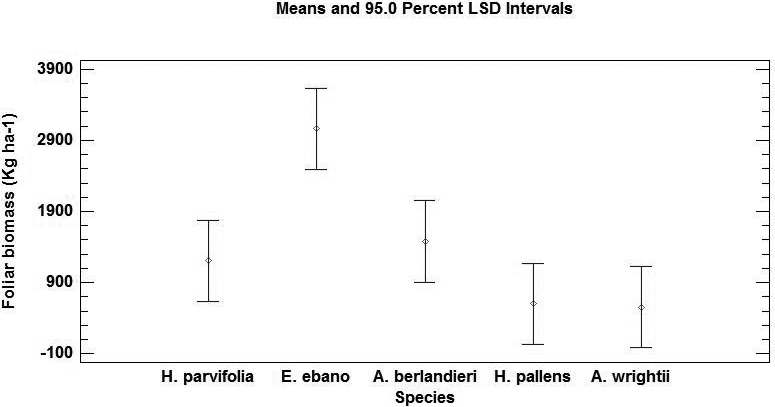
Figure 1 Annual average production of foliar biomass (Kg ha-1) and standard errors of five species in the semi-arid area of northeastern Mexico
Figure 2 presents the seasonal production, where summer showed the maximum value (12,253 kg ha-1), registered by Ebenopsis ebano, and winter showed the lower value (2,215 kg ha-1), recorded by Acacia wrightii.
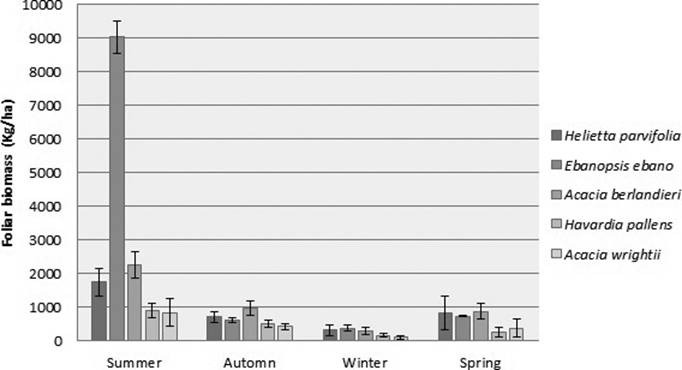
Figure 2 Seasonal foliar biomass production of five species of the semi-arid area of northeastern Mexico
The values were ranged from 103 to 371 kg ha-1 in winter, corresponding to Ebenopsis ebano and Acacia wrightii respectively; 248 – 879 kg ha-1 in spring, corresponding to Acacia berlandieri and Havardia pallens respectively; 834 – 9,029 kg ha-1 in summer, E. ebano and A. wrightii respectively; and 418 – 962 kg ha-1 in autumn, corresponding to A. berlandieri and A. wrightii respectively.
Litterfall accumulation. No statistically significant differences (P > 0.05) were detected among species. However, highly significant differences (P < 0.01) were observed among seasons of the year (Table 3). The annual average of leaf litter accumulation varied from 646 to 721 kg ha-1, values corresponding to Ebenopsis ebano and Acacia wrightii, respectively (Figure 3).
Table 3 ANOVA for Leaf litter by Season
| Source | Sum of Squares | Df | Mean Square | F-Ratio | P-Value |
|---|---|---|---|---|---|
| Between groups | 292499, | 3 | 97499,7 | 122,16 | 0,0000 |
| Within groups | 44694,8 | 56 | 798,121 | ||
| Total (Corr.) | 337194, | 59 |
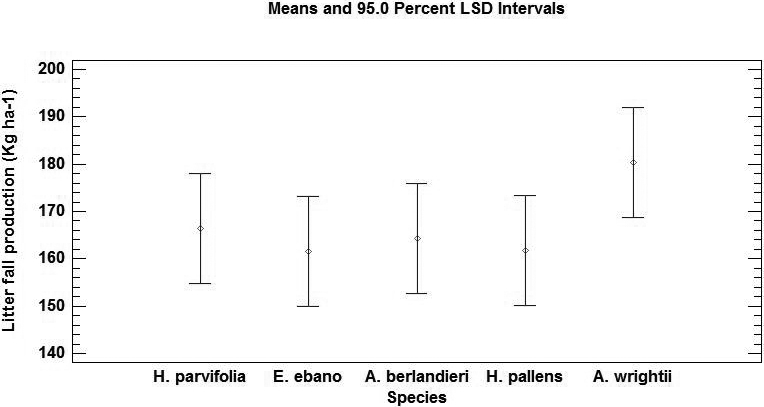
Figure 3 Annual average of litterfall deposition (Kg ha-1) and standard error of five species of the semiarid zone of northeastern Mexico
The greater amount of leaf litter accumulated was observed in winter by Acacia berlandieri (274 kg ha-1) and Acacia wrightii (296 kg ha-1), while the lower value was recorded in spring, by Ebenopsis ebano (97 kg ha-1). Summer and autumn presented the ranges from 109 to 163 kg ha-1, values corresponding to A. wrightii and Helietta parvifolia, respectively; and from 139 to 162 kg ha-1, values corresponding to A. berlandieri and Havardia pallens, respectively (Figure 4).
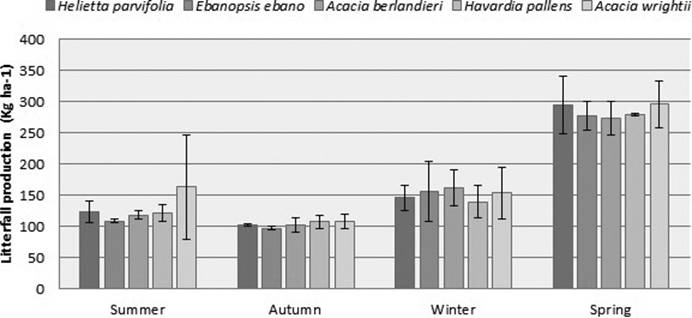
Figure 4 Seasonal rate of litterfall production (kg ha-1) and its standard deviation in five species of the semi-arid land of northeastern Mexico
Relationship between foliar biomass production and litterfall accumulation. Table 4 shows the results of fitting a linear model to describe the relationship between Leaf litter and foliar biomass. The P-value is less than 0.05 in summer and spring, indicating a statistically significant relationship between leaf litter accumulation and foliar biomass production at the 95 % confidence level, while there is a highly significant relationship (P < 0.001) between these variables in autumn. With the R-Squared statistic obtained, their correspondent models as fitted explain 27.4968 % of leaf litter variability in summer, 36.0759 % in spring and 68.7923 % in autumn. Therefore, there is a moderately strong relationship between the variables in autumn, since its correlation coefficient equals -0.829411 (Figure 5).
Table 4 Equations of the correlations between the seasonal leaf litter accumulation and foliar biomass production of five woody species of the semi-arid zone of northeastern Mexico
| Seasons | Equations of correlations | Correlation coefficient |
R2 | P-value |
|---|---|---|---|---|
| Summer | LL = 139,352 - 0,00723143 x FB | -0,524374 | 27,4968 | 0,0448 |
| Autumn | LL = 109,788 - 0,00585361 x FB | -0,829411 | 68,7923 | 0,0001* |
| Winter | LL = 149,018 + 0,00245595 x FB | 0,143127 | 2,04853 | 0,6108 |
| Spring | LL = 172,011 + 0,119417 x FB | 0,600633 | 36,0759 | 0,0179 |
LL. Leaf litter, FB. Foliar biomass
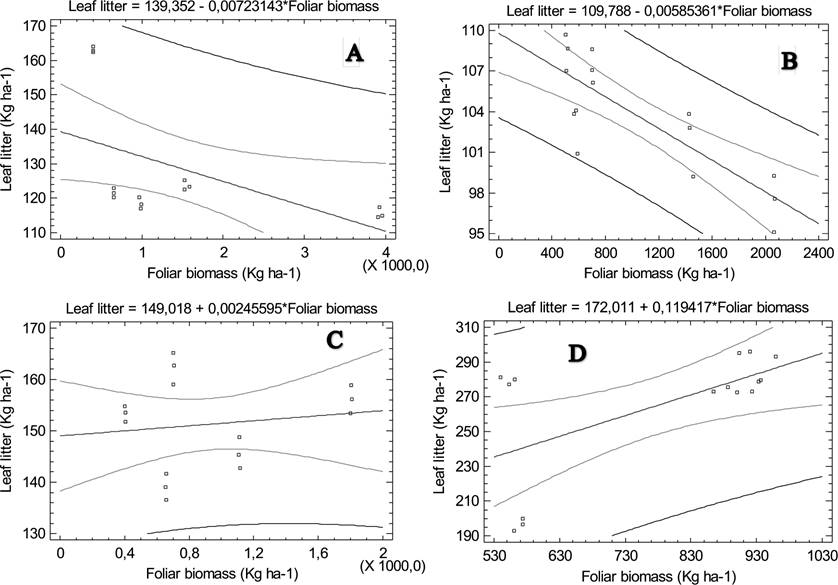
Figure 5 Relationships between foliar biomass production and leaf litter accumulation of five woody species of the semi-arid zone of northeastern Mexico, by season: A) Summer, B) Autumn, C) Winter, D) Spring.
For winter, the P-value is greater to 0.05, indicating that there is no statistically significant relationship between leaf litter and foliar biomass at the 95 % or higher confidence level. The model as fitted just explains 2.04853 % (R2) of the variability in leaf litter, and the correlation coefficient equals 0.143127, indicating a relatively weak relationship between the variables (Figure 5).
Discussion
Foliar biomass. The higher annual production of foliar biomass registered with Ebenopsis ebano can be explained by the more coverage that reported Ngangyo et al. (2014b) for the same species, which presented a dense canopy. The lowest values recorded in winter, show that the warm weather is a determining factor in the foliar productivity. As indicated by Návar-Cháidez & Jurado-Ybarra (2009), the species of scrubs bloom and produce leaves in the early stages of spring.
The evaluated values of foliar biomass obtained in this study are lower than 25,000 kg ha-1 found by Yerena-Yamallel et al. (2011) in a primary scrub, and similar to 12,930 and 13,033 kg ha-1 found by Návar-Cháidez (2008) and Búrquez et al. (2010) respectively, on a Tamaulipan thorn scrub. The variations in the obtained ranges of foliar biomass prove the wide range of growth patterns, leaf diversity and longevity, that characterize the vegetation of northeastern Mexico, according to Reid et al. (1990) and Domínguez-Gómez et al. (2013) who studied the dynamics of growth and phenological development of the Tamaulipan thorn scrub.
The high value of foliar biomass observed by Helietta parvifolia, Ebenopsis ebano and Acacia berlandieri, can be related to the larger crown area they covered in comparison to Havardia pallens and Acacia wrightii, as presented by Ngangyo et al. (2014b), by determining the canopy coverage of these five species in study. Moreover, according to Tracy (2014), H. parvifolia is characterized by high leaf area, and E. ebano is characterized by multiple branches extending in zigzag on a large space to mature. This allows considering the crown density as an important factor in the foliar biomass production.
Leaf litter. The significant differences among seasons in the amount of leaf litter imply a temporal variation of litterfall accumulation during a year. A similar result was observed by González-Rodríguez et al. (2013) in a scrubland of the northeastern Mexico. According to Prause et al. (1997), spatial and temporal variation is probably associated with the floristic composition of a site. In this study, the higher value of leaf litter was recorded in winter (December and January) and can be related to the extreme temperature conditions, since winter is the coldest season of the year, reaching temperatures of 5.2 °C (CONAGUA, 2014). Such temperatures caused an increase in the abscission of the leaves (Álvarez et al. 2009). This occurs mostly in regions with arid or semi-arid climates where gradual leaf abscission seems to be an adaptation to water stress, as indicated by López-Hernández et al. (2013).
Moreover, the species Helietta parvifolia (295 kg ha-1) and Acacia wrightii (296 kg ha-1) which registered high values of litterfall are highlighted by Ngangyo et al. (2014b), to present a high crown area. Degen et al. (1997) mentioned that the genus Acacia is a great source of foliage. This could explain that the loss of leaves was bigger in this species than the others.
The lower value recorded in spring coincides with the renewal of the leaves, which is proper this season. And Ebenopsis ebano which have multiple branches that extending in zigzag (Tracy 2014), does not lose many leaves.
The monthly variation in litterfall production and its components has been reported previously by Prause et al. (2003) who found values ranging from 0.42 to 28.2 g m-2 per year. The values provided in this study for litterfall accumulation are within the range defined for plantations established with species characteristic of poor and degraded soils (Bernhard-Reversat 1993, Lim 1988).
Relationship between foliar biomass production and leaf litter accumulation. The coefficient of determination observed in winter (R2 = 2.04853 %) indicating a relatively weak relationship between foliar biomass and leaf litter. In summer (R2 = 27.4968 %) and spring (R2 = 36.0759 %), the coefficients of determination indicating a moderately relationship between the variables. The best coefficient of determination (68.7923 %) observed in autumn indicating a moderately strong relationship between the variables, with a negative and significant linear correlation between leaf litter accumulation and foliar biomass production. These significant variations in the relationship foliar biomass/leaf litter can be explained by the weather conditions, in accordance with Estrada-Castillón et al. (2012), who found in a study of the structure and diversity of submontane scrub in northeastern Mexico, that the environmental conditions, particularly rainfall, accentuate and favor significant changes in the vegetation structure. In effect, in extreme weather conditions (winter which is the coldest season with lower temperatures, and summer which is the warmest season with higher temperatures), the plants are in stress condition, and present a difficulty in coordinating the mechanisms of production and loss of leaves, whereas in temperate conditions (autumn), there is a certain balance between loss and leaf production, so the model obtained in this case can be used for predictions of future observations. But from the physiological point of view, Návar-Cháidez & Jurado-Ybarra (2009) indicate that it is more acceptable a logarithmic relationship, that is born of origin. However, the same authors note that with the linear model, they obtained a better coefficient of determination (35 %) that is lower than the best coefficient obtained in the present study (68.79 %). Brown & Lugo (1982) had already found a relationship between total litterfall production and biomass. Also in recent studies, Agudelo & Aristizabal (2003) registered association (R2 = 74 %) between the foliar biomass and litterfall.
Leaves are short-lived organs and therefore, the recycling of nutrients previously incorporated in them and that have already carried out their contribution to photosynthesis can have a great adaptive significance (Escudero & Mediavilla 2003) and represent a considerable part of annual nutrient requirements for construction of foliar biomass.

