











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkFruit and seed sizes have an impact on plant dispersion patterns and reproductive success (Primack 1987, Bolmgren & Eriksson 2005, Bolmgren & Eriksson 2010). Nevertheless, few studies have integrally analyzed the relationships between these attributes and their ecologic implications (Bolmgren & Eriksson 2010), although they can provide relevant information for establishing ecological restoration strategies in tropical ecosystems (Orozco-Segovia & Sánchez-Coronado 2009).
In general, it has been demonstrated that fruit type influences the size of seeds; species with fleshy fruits have larger seeds than species with dry fruits (Bolmgren & Eriksson 2010). In addition, there is a positive relationship between seed size and fruit size in species with fleshy fruits, which is explained in terms of a cost-benefit approach between the increment of seed dispersal capability and the reward for endozoochory dispersal (Coomes & Grubb 2003, Moles & Westoby 2006, Bolmgren & Eriksson 2010). Another factor affecting size and number of seeds in fleshy fruits is the size of the mother plants (Moles & Westoby 2006).
Seed biomass is one of the most important functional traits for plant performance and fitness (Coomes & Grubb 2003, Orozco-Segovia & Sánchez-Coronado 2009). This characteristic is associated with the amount of energy reserves in the embryos (Cipollini & Stiles 1991, Moegenburg 1996, Aparicio et al. 2002), the probability for successful germination (Baloch et al. 2001) and the ability to avoid pathogens and herbivores (Coomes & Grubb 2003). In general, it has been reported among different ecosystems that the seed size of a plant is negatively related to the number of seeds it can produce, and plant species that produce smaller-seeds are considered competitively superior in colonizing different environments (Silvertown 1989, Coomes & Grubb 2003). Conversely, biomass is positively associated with seedling survival because larger seeds develop more vigorous seedlings (with greater biomass in their roots and cotyledons) and with greater soil emerging potential (Pieta & Ellis 1991, Aparicio et al. 2002), which confers them competitive advantages when facing the lack of resources (light and nutrients) or the negative influence of random events, such as dry periods or partial damage caused by herbivores (Coomes & Grubb 2003). Another aspect influenced by the size and shape of seeds is the amount of time they can persist in the soil seed bank (Cerabolini et al. 2003). Smaller, more compacted seeds, tend to remain in the soil longer than larger, more elongated seeds (Funes et al. 1999).
Although the importance of fruit size, seed size and seed shape on the performance and fitness of plants has been widely analyzed at the community level, little attention has been paid to the intra-specific variation of these variables and their ecological implications (Khurana & Singh 2001, Bolmgren & Eriksson 2010). The limited information available shows that variation in seed-size is very frequent within populations and even among individuals, which suggests that different selective forces regulate size variation within a population (Moegenburg 1996, Bu et al. 2007, Lázaro & Traveset 2009). At the intra-specific level, it has been recognized that seed size and shape influence germination rates, seed dormancy, vigor and survival of seedlings (Anderson & Milberg 1998, Moles et al. 2000, Baloch et al. 2001). This variation can be relevant for the survival of many species in heterogeneous environmental conditions (Khurana & Singh 2001, Coomes & Grubb 2003). For example in Albizia procera, a tropical dry forest (TDF) tree, it has been reported that larger seeds produce more vigorous seedlings with a greater ability to withstand water stress compared to smaller seeds. This could favor the survival of this species under variable dry periods (Khurana & Singh 2000).
Currently, TDF are seriously endangered due to human activities that threaten the biodiversity of these ecosystems. In México, TDF occupy only 21 % of their original extension, and annual deforestation rates are estimated to be nearly 2 % (Trejo & Dirzo 2000). Given the critical status of the TDF, strategies need to be developed to reverse the current trend of deforestation processes; thus, it is crucial to study the functional traits of seeds, their relationship with germination and the establishment of plants (Khurana & Singh 2001, Orozco-Segovia & Sanchéz 2009).
Ceiba aesculifolia (Kunth) Britten & Baker f. is a native tree from the Mexican TDF and shows great variation in size and shape of its fruits and seeds (Niembro et al. 2010). These characteristics make it an ideal candidate for studying the effects of morphological variation in fruits and seeds, seedling performance, and their possible use in restoration programs. In this study we analyzed morphological variation of fruits and seeds of this species to answer the following questions: Do size and fruit shape influence the number and size of seeds? Is there a positive relationship between seed size, germination and seedling growth? This study also discusses the usefulness of these data for the implementation of management and reproduction programs for this species in ecological restoration in the TDF of Mexico.
Materials and methods
Study species. Ceiba aesculifolia (Malvaceae) is a tree 15 to 20 m tall with a diameter at breast height of up to 60 cm. It is a monopodial tree; its trunk is covered with conical thorns, and its canopy is rounded and dense. The flowers are actinomorphic with funnel-shaped calyx and 5 brown petals up to 14 cm long. Its fruit is a dehiscent capsule with a woody pericarp, purple to bright red or reddish brown. The seeds are ovoid-globose 8 to 10 mm in diameter. Each seed has a tiny embryo with folded foliar cotyledons and a fleshy endosperm wrapping around the embryo and the cotyledons. The seeds are orthodox and can be stored at -20 °C with 15 % relative humidity (Avedaño 1998, Niembro et al. 2010). This tree species grows in preserved and secondary TDF in Mexico; therefore, it has been considered an important species for the restoration of these ecosystems (Valle-Díaz et al. 2009, Niembro et al. 2010).
Study area. This study was conducted in a lava field in the locations of “Trapiche del Rosario and Chicuasen”, in Actopan, Veracruz. In these locations, TDF fragments, secondary vegetation, riparian vegetation and agricultural lands integrate the landscape in the high river basin of the Actopan River (Castillo-Campos et al. 2007). These ecosystems develop on a lava field with upwelling rocks. The soil is heterogeneous in depth, but is mainly shallow. The TDF of this region has a very rich flora in which over 660 species of vascular plants have been recorded. The most common tree species are: Bursera cinerea, Cephalocereus palmeri var. sartorianus, Lysiloma microphylla, L. acapulcense and Pseudobombax ellipticum var. tenuiflorum (Castillo-Campos et al. 2007).
The climate is Aw1(w), warm-humid with rains in the summer (winter rainfall is lower than 5 %), with an average annual temperature of 22.3 °C (a minimum temperature of 11 °C and a maximum of 30 °C) (García 1988). The annual average rainfall is 1053.5 mm, with a dry season between October and May (average rainfall is 22 mm) and a rainy season from June to September (monthly average rainfall is 209 mm) (García 1988). Soils are shallow lithosols (Rossignol & Geissert 1987).
Data collection. Taking into account that the proximity of these collecting locations was less than 1 km, it was considered that the individuals of Ceiba aesculifolia used in this study were part of the same population. Thirty five individuals with fruits of different shapes and sizes were selected. Each tree produces a single category of form and size fruit. Between 5 and 10 fruits were collected from each individual. Two fruits per individual were randomly selected (N = 70) and fresh fruit mass, length and width were estimated for each one. From the mass distribution of the fruits, three size categories were established: small (150-250 g), medium (251-350 g) and large (> 351 g). Subsequently, the fruits were dried at room temperature until the capsules opened (3-5 days). The seeds from each fruit were counted and separated from the silky fiber that covers them. From each fruit, 20 seeds were randomly selected to determine fresh seed mass; these seeds were used for the germination test (N = 1,400). The seeds were sown in 10 germination trays with independent compartments and a capacity of 200 ml, and the trays were filled with a mixture of sand and soil (1:1) from the TDF. In each tray, 20 seeds were sown at 0.5 cm in depth. The germination tests were carried out in a greenhouse, at 24 °C. The trays were watered every day at field capacity and monitored daily for 30 days. A seed was considered germinated when the radicle emerged from the seed coat (sensuFenner & Thompson 2005). To determine seedlings dry mass, seedlings were harvested when they begun to produce true leaves (approximately 35 days after germination). The seedlings were dried for 72 hours, until they reached a stable weight and were weighed separating stems and root. Then, the root:shoot ratio was calculated.
Seed lipid and water content. Seed lipid content of each fruit category was determined in 28 mg of dry tissue in accordance with the Bligh and Dyer (1959) method of extraction. Seed water content was calculated as an oil-free percentage (OF) on a wet (WCofwb) and dry basis (WCofdb) in accordance with Caddick (2005):
where OCdb is the oil content on a dry basis.
Data Analysis. Fruit mass, fruit length:width ratio, seed fresh mass and seedlings dry biomass, were all compared among the three categories with ANOVA tests. Multiple comparisons were performed with the Tuckey´s test (P ≤ 0.05) using JMP software (Version 8.0.2., SAS Institute Inc., Cary NC, USA). The number of seeds per fruit, seed lipid and water contents (fresh and dry basis) were analyzed using a generalized linear model (GLM) with a Poisson distribution and a log link function. These statistical analyses were conducted in R 2.5.2 (R development core team) with the glme function (in the gmodels library). To fit linear effects, model contrasts were performed with the estimable function (Warnes et al. 2006).
Accumulated germination percentage of each category was adjusted to the sigmoid exponential model, Y = a + b(-cx), with the program Table Curve 2Dv 5 (AISN. Software, Inc., Chicago IL USA), with which, lag time and germination rates (velocity) for seeds of each fruit category were calculated (González-Zertuche et al. 2001). Subsequently, both variables were compared among the fruit categories, using a generalized linear model (GLM) with a Poisson distribution and a log link function. The final germination percentage among fruit categories, dry seedling mass and root shoot ratio were also analyzed with ANOVA tests. Percentages were previously arc-sin transformed. The relationships between seed and fruit variables were determined with linear regression analysis, and the following relationships were analyzed: 1) dry and fresh seed mass, 2) seed lipid content with seed dry mass, fresh mass and water content; 3) dry or fresh mass with seed water content (fresh and dry basis), 4) the relationship between seed water content on a wet basis with the water content on a dry basis 5) seedling dry mass and oil seed content. Fittings were done using Table Curve, v.3 software (AISN Software, Chicago, IL, USA).
Results
Shape and mass of fruits and seeds. Among the fruit categories of C. aesculifolia, there were significant differences in fruit mass, shape (length:width ratio) and seed mass (Figure 1). The average mass of the fruits varied between 194.2 and 382.0 g, the highest values were recorded in the large category (F (6,67) = 126.9, P < 0.0001; Figure 1a). Regarding fruit shape, based on the length:width ratio, the small and large categories showed the most elongated fruits (F (6,67) = 6.11, P < 0.01; Figure 1b). There were significant differences in the number of seeds per fruit category (χ2 = 51.27, d.f. = 2, P=0.0001), the small category had a lower mean value than medium and large categories (χ2 = 47.51, d.f. = 2, P < 0.0001; χ2 = 27.52, d.f. = 2, P < 0.000; respectively, Figure 1c). Seed fresh mass varied between 256.6 and 298.1 mg; the heaviest seeds were recorded in the large category (F (2,69) = 8.13, P = 0.0007; Figure 1d).
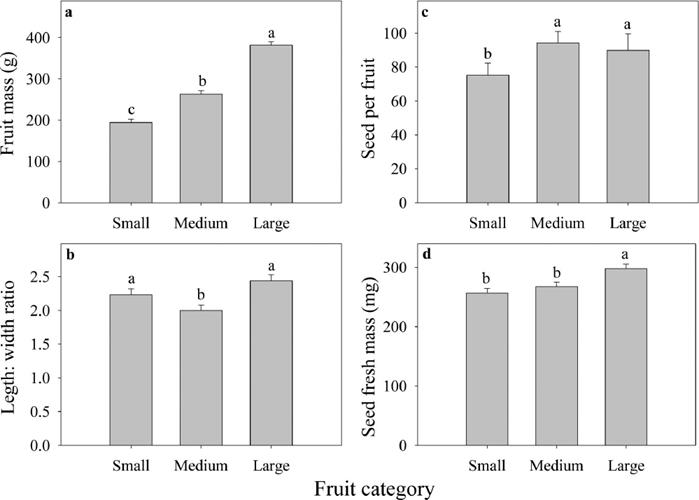
Figure 1 Morphological traits of fruits and seeds of three mass fruit categories of C. aesculifolia. Different letters on bars indicate significant differences between categories. Mean values (+SE).
The fruit length:width ratio and the number of seeds had a low but significant negative relationship (R 2 = 28, P < 0.001). The fruit mass and the seed mass had a positive relationship (R 2 = 0.22; P < 0.0001). The more spherical fruits had more seeds. The heaviest fruits had seeds with greater mass.
Seeds lipid and water content. Seed lipid content significantly varied among the categories (χ2 = 17.45, d.f. = 2, P < 0.0001) from 19.27 % to 28.89 % (Figure 2a). The significantly highest values were found in the large and medium seed categories (χ2 = 17.05, d.f. = 1, P < 0.0001; χ2 = 5.37, d.f. = 1, P < 0.05; respectively). Seed water content on fresh and dry basis also showed significant differences among the categories (χ2 = 18.88, d.f. = 2, P < 0.0001; χ2 = 23.38, d.f. = 2, P < 0.0001, respectively), with values between 9.58 ± 0.31 % and 13.49 ± 0.31 % on a fresh basis, and between 10.35 ± 0.4 % and 14.93 ± 0.26 % on a dry basis (Figure 2b, c). In both cases the highest values were found in the large category, and the lowest values were found in the small (χ2 = 12.18, d.f. = 1, P < 0.001; χ2 = 14.78, d.f. = 1, P < 0.001; respectively) and medium categories (χ2 = 15.11, d.f. = 1, P < 0.001; χ2 = 18.87, d.f. = 1, P < 0.0001, respectively).
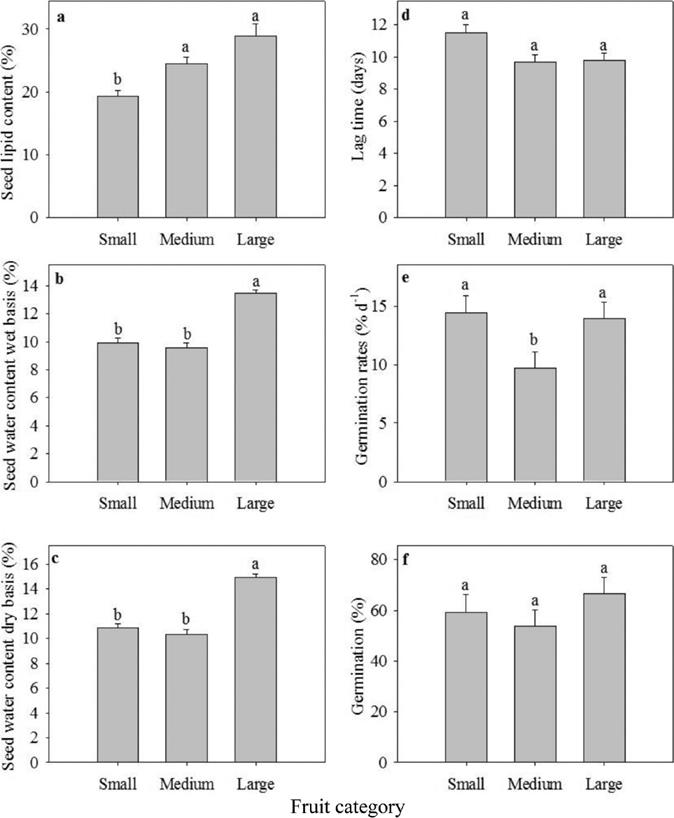
Figure 2 Lipid and water content in seeds and germination traits of the three mass fruit categories of C. aesculifolia. Different letters on bars indicate significant differences between categories. Mean values (+SE).
All relationships between seed variables of the three categories were significant and positive. The functions that describe each of these relationships, as well their statistical values, are shown in Table 1.
Table 1 Relationships between seed variables of Ceiba aesculifolia. WCofwb =water content oil free fresh basis, WCofdb = water content oil free dry basis.
| Seed variables | function | R2 | F | P |
|---|---|---|---|---|
| Fresh mass and dry mass | Y = a + bx | 0.99 | 26192.4 | 0.00001 |
| Fresh mass and lipid content | Y = a + bxc | 0.91 | 178.55 | 0.00001 |
| Dry mass and lipid content | Y = a + bxc | 0.88 | 152.52 | 0.00001 |
| Fresh mass and WCofwb | Y = a + bx3 | 0.93 | 476.09 | 0.00001 |
| Fresh mass and WCofdb | Y = a + bxc | 0.92 | 197.87 | 0.00001 |
| Dry mass and WCofwb | Y = a + bxc | 0.91 | 179.15 | 0.00001 |
| Dry mass and WCofdb | Y = a + bxc | 0.91 | 173.53 | 0.00001 |
| WCofwb and WCofdb | Y = a + bx | 0.99 | 136003.1 | 0.00001 |
Germination. Seed lag time varied between 9.8 and 11.5 days (Figure 2d), but there were no significant differences among seeds of the three fruit categories (χ2 = 4.14, d.f. = 2, P = 0.126). Germination rates varied between 9.7 and 14.4 % d-1 (Figure 2e). Seeds from the fruits of the small and large categories showed the highest values (χ2 = 25.69, d.f.= 2, P < 0.0001), and were significantly different from the medium category (χ2 = 20.22, d.f. = 2, P < 0.001; χ2 = 17.94, d.f. = 2, P < 0.0001, respectively). Final germination percentage did not change between categories (P > 0.05, Figure 2f). Accumulated germination is shown in Figure 3.
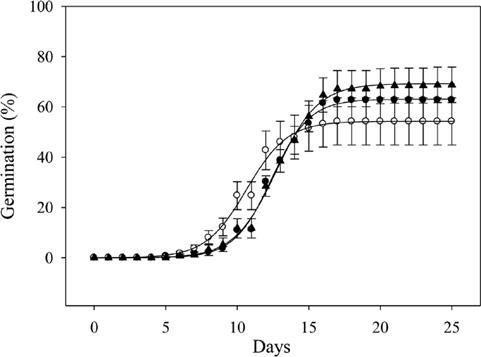
Figure 3 Cumulative germination of the seeds of Ceiba aesculifolia from the: (○) small, (●) medium and (▲) large mass fruit categories.
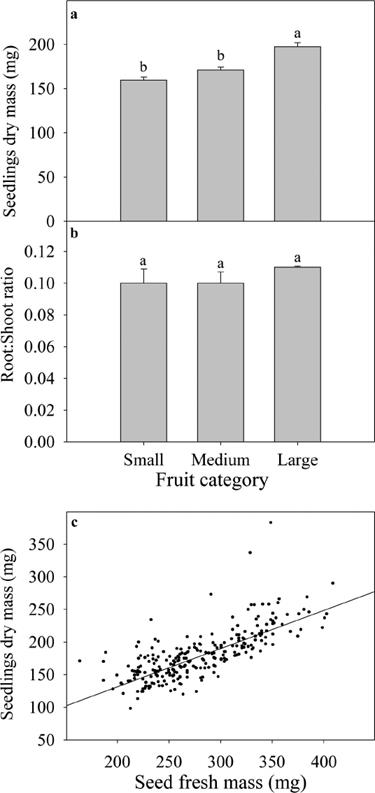
There were significant differences in seedling total dry mass between fruit categories (F (2,248) = 24.76, P < 0.0001), and the large category had the highest values (Figure 4a). In the root: shoot ratio, no significant differences were found among the categories (F (2,248) = 0.46, P = 0.625; Figure 4b). Seed mass significantly influenced seedling dry mass (R 2 = 0.62, P = 0.001, Figure 4c); heavier seeds produced larger seedlings. Seed lipid content had also a significant positive relationship with seedling dry mass (R 2 = 0.96, P = 0.02).
Discussion
In the TDFs, spatial-temporal variations in soil moisture, light, temperature, nutrients and intensity of predation can affect the reproductive traits of plants (Khurana & Singh 2001, Herrerías et al. 2008). Our results suggest that variation in the mass and shape of fruits of Ceiba aesculifolia is positively related with the number of seeds per fruit and seed mass. These results also suggest that the size of seeds affected the early growth of seedlings.
Fruit and seed traits. The relationships between fruit size and seed size are well understood in species with fleshy fruits dispersed by animals (Bolmgren & Eriksson 2010); however, among dry fruits, such as the dehiscent capsules of C. aesculifolia, these relationships have not been well studied. In general, in fleshy fruits there is a positive relationship between fruit mass and seed mass (Bolmgren & Eriksson 2010). In Ceiba aesculifolia, we found a similar relationship; the heaviest fruits produced heavier seeds. Moreover, fruit shape was related to the number of seeds; the most elongated fruits produced fewer seeds. The advantages of heavier seeds for the survival and establishment of seedlings have been recognized in moist and shady environments; however, in dry environments, their role is ambiguous and has been strongly questioned (Leishman & Westoby 1994, Orozco-Segovia & Sánchez-Coronado 2009). In the case of C. aesculifolia, heavier seeds produced larger seedlings; nevertheless, the advantages of heavier seeds for seedling establishment should be viewed with caution and tested in the field, because heavier seeds have more limited dispersal capabilities, and are more predated than smaller seeds (Herrerías et al. 2008).
Both in dry and humid tropical forests, environmental conditions experienced by mother plants in temporal or spatial scales affect the traits of their seeds (Grubb & Coomes 1997, Khurana & Singh 2001, Soriano et al. 2011). In some species of the TDF, it has been recorded that heavier seeds, with greater nitrogen and lipid content are produced in drier years (Soriano et al. 2011); conversely in trees and shrubs of the rainforest, nutrient concentration in the seeds is associated with the nutritional status of the soil in which the mother plants grow (Grubb & Coomes 1997). In the case of Ceiba aesculifolia, there is no evidence to help us discern which factors determine the variation recorded in fruits and seeds. The soil in the lava field where the TDF develops is heterogeneously deposited within rocks cavities, pockets, crevasses and slight depressions. In some of these, the soil depth and organic matter is higher than in others (Castillo-Campos et al. 2007). This soil variation might determine the resources available for the reproduction of individuals of C. aesculifolia. Trees with heavier seeds and fruits are mainly distributed in deeper soil sites (cavities, pockets) close to the farming areas (30 % of the trees in the collection showed this condition); while individuals with smaller fruits are more frequently found in areas with shallow soil (N. Velázquez-Rosas unpublished data).
Seed size, germination response and establishment of seedlings. In the TDF trees, it has been recognized that seed size affects germination percentage (Khurana & Singh 2001); heavier seeds have higher germination rates than smaller ones (Dileep et al. 1994, Singh & Khan 1998), because they have greater energetic reserves that can be used for germination (Khurana and Singh 2001). Previous studies showed that in Ceiba aesculifolia, heavier seeds germinate in greater proportions (95 %; Herrerías et al. 2008). Our results differ, because germination percentages recorded in all seed sizes were lower than 60 % and did not differed between fruit size categories. In C. aesculifolia, it is difficult to discern the causes that determine the relationship between seed size and the evaluated germination traits in terms of energy reserves (as has been suggested for other species), but we found no relationship between germination and seed lipid content. This pattern has been reported for other species of TDF (Vargas et al. 2015). The differences in the recorded germination responses in C. aesculifolia could be originated genetically, or due to phenotypic plasticity associated with maternal effects; the latter has been noted as a frequent trait among species growing in environments with heterogeneous and unpredictable conditions (Fenner 1991, Khurana & Singh 2000, Khurana & Singh 2001, Moles & Westoby 2006, Soriano et al. 2011).
Seed size and its influence on the establishment and survival of the seedlings have been discussed under a cost-benefit approach, between the number of seeds produced and their probability of establishment (Moles & Westoby 2006). In general, it is assumed that species with small seed produce more seedlings, but their establishment and survival success are lower, compared to those of species with larger seeds (Moles & Westoby 2006). These differences may be due to the amount of reserves stored by seed cotyledons, which are used for growth in early development stages (roots and photosynthetic tissue) (Hewitt 1998, Kitajima 2002, Soriano et al. 2011). In the case of Ceiba aesculifolia, trees with larger seeds, which show the highest contents of lipids and water, produced seedlings with greater biomass, which could increase their establishment and survival probabilities. This is consistent with what was reported for Albizia procera; larger seeds produce the most vigorous seedlings, and they have a greater ability to withstand water stress for longer periods, thus increasing their chances of establishment (Khurana & Singh 2000).
The persistence of a wide variation in the size of the seeds at an intra-specific level has been interpreted as a favorable condition for many species to thrive in highly heterogeneous, or unpredictable environments (Khurana & Singh 2000, Khurana & Singh 2001, Coomes & Grubb 2003). The results recorded for Ceiba aesculifolia in the relationships among seed size, lipid and water content and seedling biomass suggest that seed size variation can determine differential capabilities for settling in the TDF. It is necessary to develop field experiments to analyze where the most suitable microclimate conditions for germination, development and establishment of seedlings of different morph seeds are found.
Implications for ecological restoration. Knowledge of the functional characteristics of seeds and germination patterns, as well as the ecological requirements for germination and seedling growth, will encourage the development of ecological restoration strategies for the recovery of ecosystems (Khurana & Singh 2001, Orozco-Segovia & Sánchez-Coronado 2009). The ability to adapt to various environmental conditions may be an important attribute found in Ceiba aesculifolia that can be used in ecological restoration programs of the TDF (Valle-Díaz et al. 2009). For instance, in TDFs with nutrient deficient soils and low water retention, the selection of large seeds of a desired species might increase their establishment and survival success rates (Khurana & Singh 2001). This strategy can be applied for the introduction of C. aesculifolia in disturbed sites, whereas the use of small and medium size seeds could be considered for sites with more favorable environmental conditions. The introduction of adequate phenotypes to specific microclimatic conditions can increase their establishment success in restoration programs of the TDFs.

