











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Soil seed banks play an important role as sources of regeneration (Fenner 1985, Peterson & Carson 1996). The soil seed bank depends on the dynamics between local seed rain, germination, predation and dispersal (Álvarez-Buylla & Martínez-Ramos 1990, Dalling & Denslow 1998). Soil seed banks are reservoirs mainly for pioneer species as they represent a pool of regenerative potential by replacing the adult plants that die of natural causes or because of human actions (Fenner 1985, Peterson & Carson 1996, Hopfensperger 2007, Kwiatkowska-Falińska et al. 2012).
The seeds may be stored at different depths depending on their size, soil structure and dispersers (Werner & Platt 1976, Saatkamp et al. 2009). Seed banks can be classified as persistent or transient according to their duration (Thompson & Grime 1979). Transient seed banks are those containing seeds viable for up to 1 year, while in persistent seed banks seeds remain viable for more than 1 year (Thompson et al. 1997). The depth at which seeds of a particular species are found can depend on the successional age of the site (Werner & Platt 1976), soil characteristics and dispersers. Changes undergone by community seed banks may permit the exploration of species replacement mechanisms during succession (Connell & Slatyer 1977, Hills & Morris 1992).
Early successional plant species produce abundant quantities of seeds that are highly dependent on light for germination. These plants are essential in early forest successional stages (Fenner 1985). In forest successional progress the level of available light declines and the seed bank density of early successional species also decreases (Quinta-Ascencio et al. 1996, Dupuy & Chazdon 1998, Baider et al. 2001, Maza-Villalobos et al. 2011).
Two methodologies have been developed to assess the presence of soil seed banks: germination and direct separation (Ishikawa-Goto & Tsuyuzaki 2004, Price et al. 2010, Gonzalez & Ghermandi 2012). In the former, germinated seedlings are identified and quantified (Roberts 1981, Ter Heerdt et al. 1996), while in the latter the seeds are separated by both seed size and density. Seeds are separated from the soil by washing, sieving or flotation (Malone 1967, Roberts 1981, Tsuyuzaki 1994). Depending on the method employed, differences in both density and species composition may be found, even in the same location (Tsuyuzaki 1994, Ishikawa-Goto & Tsuyuzaki 2004, Bernhardt et al. 2008, Price et al. 2010). Most studies that have used these two methods to compare the soil seed bank have found different results and concluded that the two methods are complementary (Price et al. 2010, Gonzalez & Ghermandi 2012). Despite these differences, most studies only employ the germination method (Jaimes-Sánchez & Rivera-Ospina 1991, Cárdenas et al. 2002, Acosta 2004, Montenegro & Vargas 2005, Moscoso & Diez 2005).
Andean and high Andean ecosystems are very important in Colombia because of their high biological richness and endemism (Olson & Dinerstein 1998). These ecosystems are, however, under considerable threat, since only 27 % of the original cover remains (Etter 1993), because they have been exposed to high levels of human intervention, such as conversion to land for crop and livestock production. This leads to fragmentation processes and forest cover reduction, with an associated loss of biodiversity and ecosystem services (Cavelier & Tobler 1998, Fandiño & Ferreira 1998). Clearance of old-growth forest and subsequent land abandonment has led to the establishment of different forest successional stages, including pasture, shrubland and secondary forest (Sánchez et al. 2007). Given the constant disturbances to which these ecosystems are exposed, it is essential to develop strategies to increase diversity in degraded sites and to recover connectivity between fragments (Dalling & Denslow 1998, Meli 2003, Cardona-Cardozo & Vargas-Ríos 2004). Depending on the particular ecosystem, soil seed banks may be among the most important sources of regeneration, although both seedling banks and seed rain are important contributions for natural recovery of the structure and function of ecosystems (Baider et al. 2001, Martins & Engel 2007, Vieira & Proctor 2007).
Few studies exist of the north-eastern Colombian Andean forests, and to our knowledge, there is only one study that explores the soil seed banks of this area (Jaimes-Sánchez & RiveraOspina 1991). But this particular study does not describe the regenerative potential of these ecosystems, only basic processes. Thus, we believe there are still gaps in the knowledge regarding the ecology of Andean forest soil seed banks, and in particular, there is a requirement for studies conducted in the context of succession. We hypothesize that the soil seed banks of different forest successional stages are different in terms of density, structure, and species composition. For example, seeds of early successional species may be more abundant in shrublands with a higher density in the top layers of the soil, while intermediate or late species could be expected to be more abundant and diverse in old-growth forests; these species would form seed banks in deeper layers, which may indicate successional mechanisms. Specifically, in this study, we try to answer the following questions: 1) What is the most effective method for describing the composition and structure of the soil seed bank of Andean forests? and 2) What is the composition and structure of soil seed banks in old-growth forest and shrubland sites in the Andean montane tropical forest of the North Santander region of Colombia?
Methods
Study site. Our study was conducted in the El Volcán reserve, which covers approximately 5,900 ha in the southern state of Pamplona, Colombia (7o 20’ 57.4” N; 72o 42’ 12.2” W). Elevation ranges from 2900 to 3500 m a.s.l., within a rugged landscape featuring canyons and steep slopes. The study area comprises the source of the Pamplonita River, which supplies water to the city of Pamplona. The dominant vegetation in the area is Andean forest and alpine páramo (Sánchez & Gelviz-Gelvez, 2004). The Andean forest comprises ecosystems that are located between 2,800 and 3,200 m, with trees 8-15 meters tall and cold climates with temperatures of almost 0 °C to 21 °C and two peaks of precipitation, from March to October and from August to February. Species characteristic of these environments are Weinmannia balbisiana, Clusia multiflora, Ternstroemia meridionalis and Drimys granadensis (Cuatrecasas 1958, Murcia-Rodríguez 2010, Cantillo & Rangel 2011). The successional stages were confirmed by previous studies of the vegetation (Sánchez & Gelviz-Gelvez 2004, Murcia-Rodríguez 2010); and interviews with scientists, farmers and forest guards. Land use for crops and cattle has provoked a fragmented matrix of successional stages and old-growth forest including recently abandoned pastures and shrubland with different ages of abandonment. The shrublands used in this study are considered to be one successional stage with an age of 25 years since abandonment following agricultural use. These shrublands are defined by a high density of shrubs (80 % of stems), with average height 2-4 m height and < 5 cm dbh. The old-growth forest is uncleared forest with an overall closed canopy, dominated by slow-growing tree species > 12 m in height (Cortés 2003) and abandoned for 50 years or more since their agricultural use (Murcia-Rodríguez, 2010). In our case the shrubland had been abandoned for 25 years since its agricultural use and the old-growth forest to our knowledge has never been cleared (Murcia-Rodríguez 2010).
Sampling design. To evaluate differences in the soil seed bank along the secondary succession gradient, we compared shrubland and old-growth forests. Soil sampling was conducted in June 2010. For each study site, 15 samples of soil were taken randomly from each site of 30 × 30 cm quadrants. The total volume was approximately 13,500 cm2, which weighs more than the minimum volume proposed by Cardenas et al. (1999). Each sample was divided into two subsamples of 6,700 cm2 for testing the evaluation methods of germination and direct separation. In each subsample, three soil depths were taken: 0-5 cm, 5-10 cm and 10-15 cm, each 2250 cm2 (Chippindale & Milton 1934).
For the germination method, soil cores were sieved to remove organic debris and stones. The subsamples (0-5 cm, 5-10 cm and 10-15) were placed in a 30 × 30 cm plastic tray in a shade house covered with plastic mesh (< 0.5 mm aperture) located in an open area at 3,200 m a.s.l. in the El Volcán reserve. Samples were watered daily in order to induce germination. Every week, new emerged seedlings were identified, counted and removed to avoid confusion with newly emerged plants. The samples were stored under dry conditions (5 °C) in paper bags until late August, when the rainy season started, since the beginning of germination in the Andean forest usually coincides with the rain. Monitoring started in September and ended 10 months later when germination stopped. To check for contamination, 10 additional control trays containing substrate that had been sterilized by adding boiling water to the soil samples were included and distributed randomly among the experimental trays. No seedling emergence was recorded in the control trays during the study period. Samples of all morphospecies were grown in pots to facilitate taxonomic identification. Identification was carried out by local botanists and professional taxonomists with expert knowledge of the local flora. Voucher species were deposited at the Catatumbo-Sarare Herbarium (HECASA) of Pamplona University.
For the direct separation method, the second soil subsample was separated into four fractions by washing it with a strong jet of water through a coarse (10 mm) and three fine sieves (5 mm, 2 mm and 1 mm). Sieved soil was visually inspected using a stereoscopic microscope, and all seeds ≥ 1 mm diameter were separated. Seeds were identified to the species level by utilizing a reference collection at the HECASA Herbarium at the University of Pamplona, Colombia. We conducted seed viability tests with 10 % of the seeds of each species, employing the tetrazolium chloride method (Costa et al. 2003).
Based on the literature of the local flora and previous studies, each seedling was identified to species or genus level and categorized into one of four successional groups according to the vegetation description provided by Sánchez et al. (2007) and Murcia-Rodríguez (2010): i) disturbed, ii) early succession, iii) intermediate succession and iv) late succession (Sánchez et al., 2007; Murcia-Rodríguez, 2010). Disturbed species are herbaceous species from grassland communities that appear after removal of the shrub and tree community. Plants from this group are short-life cycle species (1-2 years). Early succession species are species of the shrubland successional stage; vegetation consisting of high density shrubs with an average height of 4 m and stems < 5 cm dbh, and 20 to 25 years abandonment. Intermediate successional species are mainly trees 8-10 m high in secondary forests with abandonment times of 35-40 years. Late successional species are those from the old growth forest. In general they are slow-growing species 12 meters in height and with ages of abandonment of 50 years or more. Analyses were carried out based on these groups.
Data analysis. The first analysis was carried out to test the effectiveness of the two evaluation methods separately for old-growth forest and shrubland. We used a paired t-test to compare estimates from the two methods obtained from the same soil sample. A second analysis was then used to compare seed density among habitats and soil sample depths for seed density obtained from pooling the estimates from the two methods. Since Soil Layer is a trial factor, due to repeated measurements on the same plot, the data were analyzed by fitting a generalized linear mixed model (GLMM). Also, because seed estimates were obtained based on counts, we used a Poisson error distribution (Bates et al. 2012, Crawley 2012). In these analyses, Habitat (with two levels; old-growth forest and shrubland), Soil layer (with three levels: top, middle and bottom layer) and the interaction between these two factors (Habitat × Soil layer) were used as discrete fixed factors. Plot nested within Soil Layer was included as a random factor (Pinheiro & Bates 2000, Faraway 2005, Bates et al. 2012, Crawley 2012). We tested overdispersion by estimating the deviance divided by the approximate degrees of freedom. A model was overdispersed if the ratio was ≥ 2. For overdispersed models, an alternative negative binomial model may be applied using the glmmADMB package (Crawley 2012, Fournier et al. 2012, Skaug et al. 2012); however, none of the models showed overdispersion.
A third group of analyses were conducted for each of the four different successional groups (disturbed, early, intermediate and late). To carry out these analyses, we also pooled estimations from the two methods. We used the same generalized linear mixed models with a Poisson error distribution (Bates et al. 2012, Crawley 2012) and Habitat, Soil layer and their interaction (Habitat × Soil layer) as discrete fixed factors and Plot nested within Soil Layer as a random factor (Pinheiro & Bates 2000, Faraway 2005, Crawley 2012, Bates et al. 2012). We also tested overdispersion as explained above. All analyses were carried out in R 3.2.4 (R Development Core Team 2016) using the lme4 package (Bates et al. 2012). Saturated models, which included the main terms and the two-way interactions, were fitted first. The models were then reduced and simplified based on sequential removal of non-significant fixed effects and detecting significant changes in residual deviance at each step (Crawley 2012). Through the use of the glmer function and maximum likelihood ratio tests (for fixed effects) we obtained the significance of terms. We tested the effects of removing factors and the interaction and to evaluate the significance of terms in the model, and all non-significant terms were removed (Pinheiro & Bates 2000, Faraway 2005). The reported likelihood ratio test (LRχ2), degrees of freedom and p-values are those from the likelihood ratio tests (Pinheiro & Bates 2000, Crawley 2012).
To calculate the equitability of species between habitats, rank-abundance curves were generated by plotting the abundance of each species as a function of its abundance rank within the community (Mueller-Dombois & Ellenberg 1974). Logarithmic regression models were then fitted (Izsák 2006). Ordination (an estimate of species dominance) and slopes of the curves were estimated based on these models. An analysis of co-variance (ANCOVA) was applied to test differences in community evenness between habitats (old-growth forest and shrubland).
Finally, to test differences in seed viability, percentage viability was compared among habitats, soil layers and their interaction using a GLM with binomial distribution. The ANCOVA and GLM-binomial were also calculated with R 3.2.4 (The R Development Core Team 2016) using the stats package ver. 3.30 (The R Development Core Team 2016).
Results
Direct separation vs. germination method. Pooling the results from the two methods, we recorded a total of 3,503 seeds belonging to 21 morphospecies. Eight of these morphospecies were identified to species, 12 to genus and one to family (Table SI1). Comparing the two methods in terms of seed detection and estimating soil seed bank density, the direct separation method significantly detected the highest number of seeds both for old-growth forest (18-fold; Figure 1) and for shrubland (222-fold; Figure 1). In addition, this method captured a greater diversity and abundance of species from both successional stages. Furthermore, the separation method was more efficient because it found more species: 17 species (16 in old-growth forest and 9 in shrubland) compared to only 13 with the germination method (13 in old-growth forest and 4 in shrubland forest). Nevertheless, the germination method revealed four small-seeded species that had gone undetected by the direct separation method (Escallonia sp., Lachemilla sp., Smilax sp., and Valeriana sp., Table SI1).
Table SI1 Seed bank density (seeds.m2, 20 cm depth) for identified taxa recorded in old-growth forest and shrubland sites at reserve El Volcán, Pamplona, Colombia. Species are arranged by successional forest and sampling method.

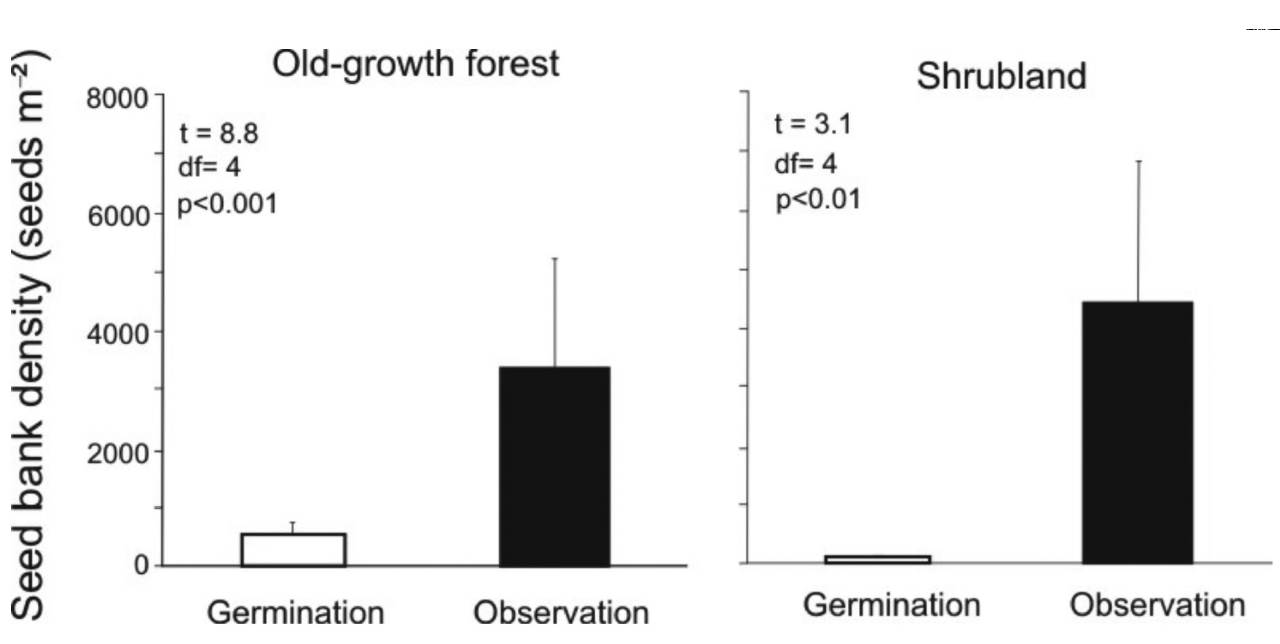
Figure 1 Soil seed bank density in old-growth forests and secondary forests of Andean forest in the El Volcán reserve, northeastern Colombia, comparing the different estimation methods for each successional stage. Error bars represent one standard error. The paired t-test results are included.
Seed bank density. The seed bank density of the total community, estimated from pooling the direct separation and germination method results, was significantly higher in the old-growth forest than in the shrubland (Table 1). Seed density in the old-growth forest increased with soil depth: lower density values were found in the top layer (204.3 ± 93.7 seeds m-2), followed by the middle layer, and the highest values were found in the bottom layer (523.5 ± 255.8 seeds m-2 Figure 2; Table 1). The opposite pattern was found in the shrubland, where seeds were concentrated in the top layer (although a wide variation was found with mean ± SE = 204 ± 245 seeds m2), containing about 65 % of the total seeds. The middle and bottom layer had lower seed density (Figure 2; Table 1).
Table 1 Statistical analysis of the soil seed bank densities for the total community and for four different successional plant groups in two stages of Andean forest in Pamplona, northeasternn Colombia. χ2, degrees of freedom and p-values correspond to a generalized linear mixed model. * and ns denote significant and nonsignificant factors, respectively.
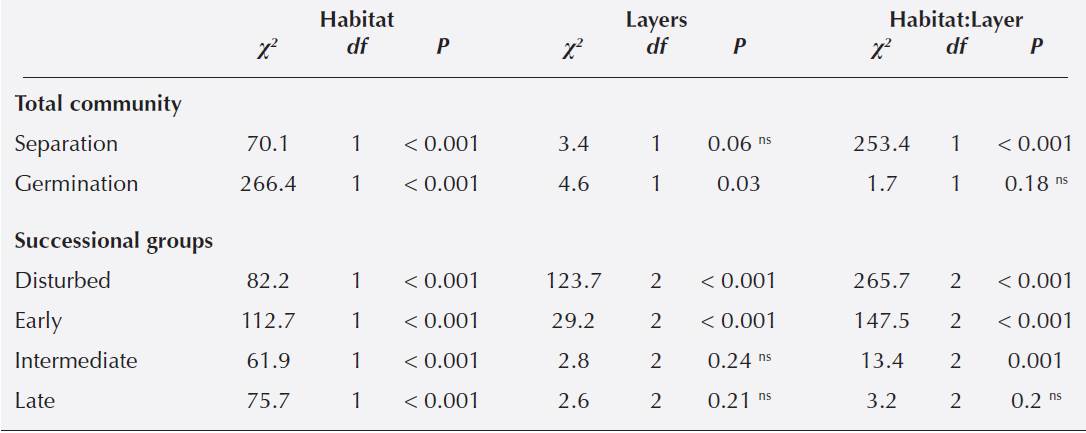

Figure 2 Soil seed bank density in old-growth forests and secondary forest of Andean forest in the El Volcán reserve, northeastern Colombia, comparing the different estimation methods of direct separation and germination. Bars represent different soil sample depths: 0-5cm white, 5-10 cm light grey and 10-20 cm dark grey. Error bars represent one standard error.
Seed bank community structure. We recorded the presence of all four different plant groups in the soil seed banks of both habitats. The relative abundance of each group differed significantly between habitats (χ2 = 202.8, P < 0.001; Figure 3). The shrubland was dominated (up to 91 %) by seeds of plants from disturbed areas, while only 74 % were from this group in the old-growth forest (Figure 2). In contrast, in the soil seed bank of the old-growth forest, seeds from intermediate and late successional stages were recorded with 4 and 20-fold higher densities than those from shrubland, respectively (Figure 3).
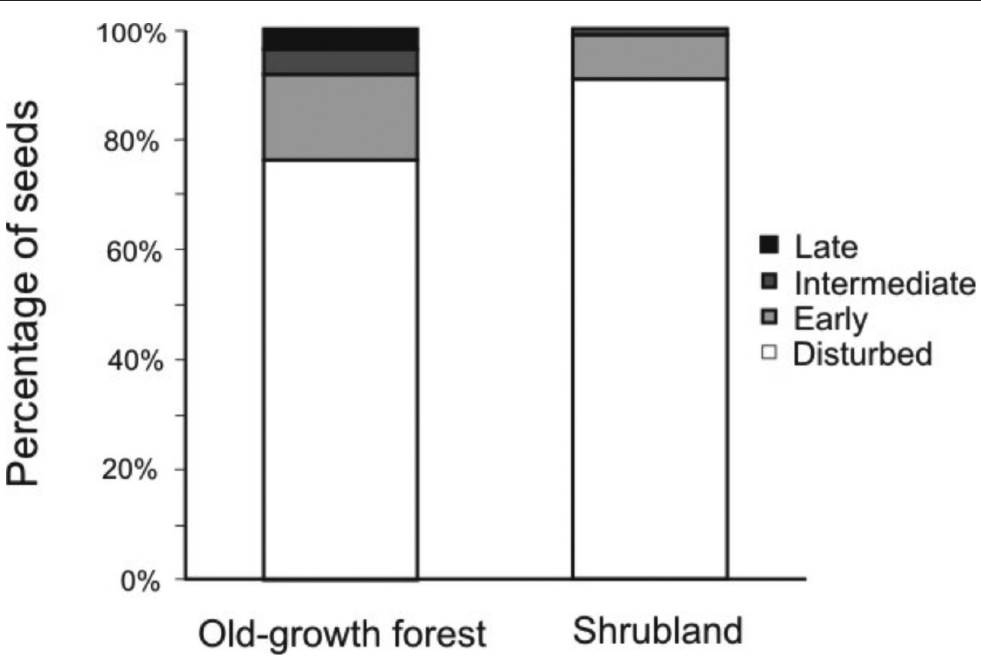
Figure 3 Structure of the soil seed bank in two successional stages of Andean forest in the El Volcán reserve, northeastern Colombia. The structure is based on four different successional groups pooling seed densities from the separation and germination methods.
The disturbed group showed higher seed bank density than the other groups. Shrubland is especially dominated by this group of plants in the top soil layer with a mean of 113.7 (± 69.2) and very low or null density in deeper layers. In the old-growth forest, the density was significantly similar among layers (Table 1, Figure 4).
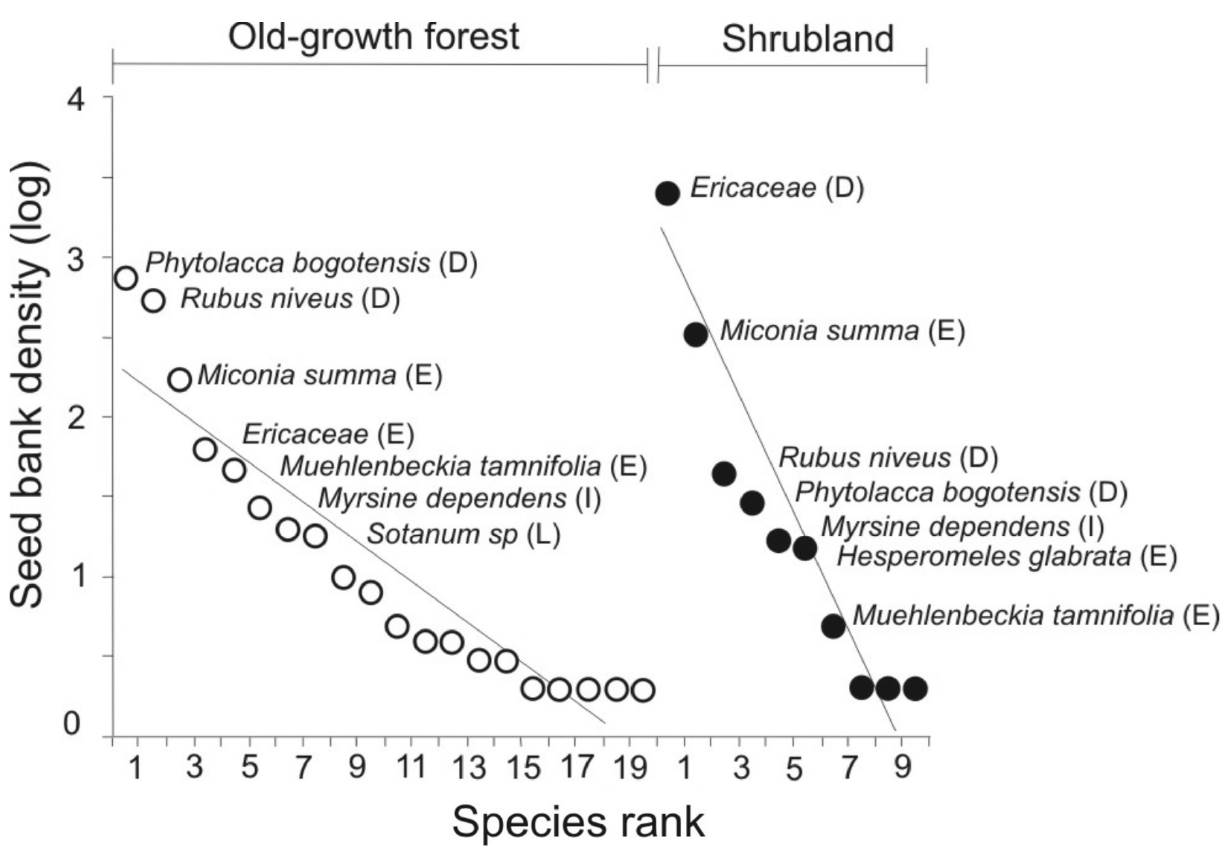
Figure 4 Dominance of communities based on rankabundance curves for two successional stages of Andean forest in the El Volcán reserve, northeastern Colombia. The letter beside the species name denotes the successional group to which that species belongs: D = disturbed, E = early, I = intermediate and L = late species. The y-axis is scaled logarithmically.
For the early plants group, the highest seed bank density was found in the middle soil layer in the old-growth forest, although it did not differ significantly from the other layers (Table 1, Figure 4). The seed bank densities of the species from the two more advanced successional groups (intermediate and late) showed significantly higher values in the old-growth forest than in the shrubland and densities among layers were similar. Late species in particular are relatively absent in the shrubland (Table 1, Figure 4).
The slope of the rank-abundance curves differed significantly between habitats (F1,17 = 7.24, P = 0.001), being more pronounced in the shrubland (b = -0.33) than in the old-growth forest (b = -0.14). This indicates a higher dominance of species in the shrubland and higher equitability in the old-growth forest. Ericaceae and Miconia summa, from the disturbed and early groups, were highly dominant in the shrubland and accounted for around 90 % of the seeds. In contrast, in the old-growth forest, five species were most abundant, and while these also corresponded to disturbed and early groups, there was no clearly dominant species. Moreover, most of the less abundant species, such as Solanum sp., Oreopanax and Bomarea sp., belonged to the intermediate and late groups, which were not found in the shrubland (Figure 5).
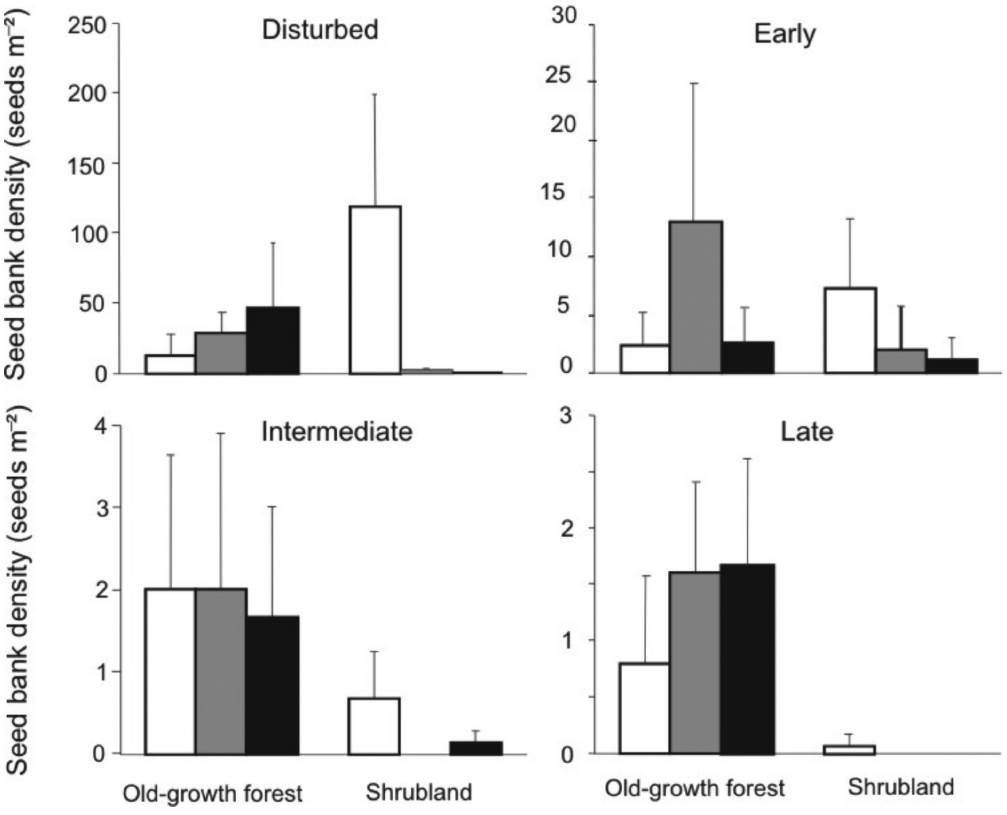
Figure 5 Soil seed bank density in two successional stages in the El Volcán reserve, northeastern Colombia. Charts indicate the four different successional groups. Bars represent different soil sample depths: 0-5cm white, 5-10 cm light grey and 10-20 cm dark grey. Error bars represent one standard error.

Figure 6. Proportion of viable seeds in the soil seed bank in old-growth forest and secondary forest of Andean forest in the El Volcán reserve, northeastern Colombia. Bars represent different soil sample depths: 0-5cm white, 5-10 cm light grey and 10-20 cm dark grey. Error bars represent one standard error.
Viability. Overall the viability of seeds found with the direct separation method was low, with only 6.8 % and 5.9 % of the seeds from old-growth forest and shrubland, respectively, being viable. In general, viability decreases significantly with depth in the soil (Lay: χ2 = 8.3, df = 2, P = 0.01). In the old-growth forest, the seed bank from the bottom layer still showed viability, but this was not the case in the shrubland, where none of the seeds recorded from this depth were found to be viable (Lay:Hab: χ2 = 7.6, df = 3, P = 0.05, Figure 5).
Discussion
Direct separation vs. germination method. The fact that the diversity and density of the species found by using the direct separation method was higher compared to the germination method can be explained by three non-exclusive factors: 1) physiological, such as seed dormancy, longevity, and germination traits (Saatkamp et al. 2009, Gonzalez & Ghermandi, 2012); 2) ecological, such as storage time, germination requirements and high rates of dispersal/predation (Thompson et al. 2003, Daws et al. 2005, Walck et al. 2005, Bernhardt et al. 2008, Esquivel et al. 2008, Price et al. 2010, Gonzalez & Ghermandi 2012); and 3) morphological, such as seed size and seed coat (De Villiers et al. 1994, Facelli et al. 2005, Venable 2007, Price et al. 2010, Gonzalez & Ghermandi 2012). Similar results were found in a study with seed banks from natural grasslands in Patagonia by Gonzalez and Ghermandi (2012). These authors report a seed density which is four times higher with the direct separation method than with the germination method.
However, in the use of these two methods, there was very little overlap between them in species composition. Small seeds (< 1mm) were only found with the seedling emergence method, whereas the seeds of species with specific germination requirements were found with the seed extraction method. Similar results area also reported by Ishikawa-Goto and Tsuyuzaki (2004) in an evaluation of seed banks of Japanese broad-leaved plants from the old-growth forest. They compared two standard seed-bank estimation techniques (germination and flotation), finding that flotation could detect more species and seeds in the soils than germination. However, many species that were extracted by flotation did not germinate because the seeds cannot be readily germinated and are structurally weak.
All of the studies evaluating seed banks in high Andean ecosystems in Colombia use the germination method exclusively (Jaimes-Sánchez & Rivera-Ospina 1991, Acosta 2004, Cardona-Cardozo & Vargas-Ríos 2004, Moscoso & Diez 2005, Cantillo et al. 2008). We believe that this is a limited method by which to characterize the soil seed banks of these ecosystems, principally because species with low seed production are difficult to detect in the soil seed bank (Thompson & Grime 1979). Therefore, when the goal of a study is to compare successional stages in order to evaluate the regenerative potential of the ecosystem through the soil seed bank, the germination method may be biased and should not be employed as the only method. Analysis of seed bank composition by a single method may produce differences in terms of density and diversity of species since the extraction method includes apparently healthy but not viable seeds (Warr et al. 1993) and the germination method only detects the germination fraction of the seed bank (Brown 1992). We therefore believe that the method of direct separation should be used as a complementary method in combination with another method for a more precise characterization of the soil seed bank in the Andean forest. The results of this study should be interpreted with caution.
Soil seed bank density. When the direct separation method was used, the total density of the soil seed bank in the old-growth forest was lower and showed less variation than that found in the shrubland. In general, it is accepted that old-growth forests have lower soil seed bank densities than those in disturbed areas (Gómez-Pompa et al. 1972, López-Toledo & Martinez-Ramos 2011). Comparing the soil seed bank of the old-growth forest with the sole previous study of Andean forests in Colombia and elsewhere, we found an approximately 0.4-fold lower density (1,813 seeds m-2; Jaimes-Sánchez & Rivera-Ospina 1991). The dominant species in that study included a non-pioneer tree (Weinmannia tomentosa), pioneer herbs, lianas and shrub species characteristic of the Andean forest and alpine páramo. Many other species, however, such as cyperaceae, shrubs and herbs, were also included. Species such as Hypochoeris radicata and Phytolacca bogotensis, which are characteristic of disturbed and agricultural areas, were found with very high densities (Jaimes-Sánchez & Rivera-Ospina 1991). The study was conducted on a small forest reserve that is highly fragmented and disturbed, as well as being surrounded by pasture and croplands, which may explain the presence of this high seed bank density (Jaimes-Sánchez & Rivera-Ospina 1991).
We found that the density of soil seed banks of old-growth forests increased with depth. The same trend was found by Kjellsson (1992) in a study of seed banks in Danish deciduous forest. Upon evaluating the distribution pattern in soil, he found that small seeded grasses and herbs with hard seed coats (e.g. Carex and Rubus spp.) were predominantly located deep in the soil. He related the depth at which the seeds were found with their ability to survive long term (Kjellsson 1992). Although this tendency is not generalized, it has been observed in some species such as Carex pilulifera (30 yr, Hill & Stevens, 1981). For the other part in shrubland, higher densities were mainly found in the top layer (0-5 cm). Other studies of high Andean ecosystems, such as páramo (Vargas & Rivera-Ospina 1990, Cardenas et al. 2002), cloud forest (Alvarez-Aquino et al. 2005) and oak forest (Moscoso & Diez 2005) also report higher seed densities in the top layer. In our study, the shrubland had higher seed density in the surface soil, while seed density in the old-growth forest was highest at greater depth. The presence of seeds in deeper soil layers may be explained by the vertical transport of seeds by water movement or animals such as ants, beetles, worms and rodents, horizontal transport by wind, or runoff migration into deeper soil layers (Garwood 1989, Gutterman 1993, Dalling & Denslow. 1998, Cárdenas et al. 2002). These seeds may play a key role, especially in natural gap areas produced by treefall, when seeds become exposed and can initiate the regeneration process (Martinez-Ramos et al. 1988).
Seed bank community structure. In our study, we found a clear successional trend, as the shrubland was mainly dominated by disturbed and early species. While in old growth forest there was a high density of disturbed species, intermediate and late stage species were also present. The study by Martins and Engel (2007) in a Brazilian tropical forest supports these results; these authors report a higher proportion of secondary species in the soil seed banks of disturbed sites, which demonstrates the importance of the seed bank for forest succession and restoration. The soil seed banks of less disturbed sites, however, presented a higher abundance of trees, shrubs and lianas. Similar results have been found by Baider et al. (2001) in forest stands of different fallow ages. These authors report dominance by herbaceous plants at the early successional stage (5 years old), whereas woody species dominate the advanced stages, such as secondary (27 years old) and old-growth forest. Elsewhere other studies report similar results to those described here (Ma et al. 2011, Maza-Villalobos et al. 2011).
Early successional pioneer species are common in the seed banks of several successional stages, including forest (Maza-Villalobos et al. 2011). In our case, we found species such as Miconia summa, Cestrum parvifolium, Hesperomeles goudotiana, Muehlenbeckia sp., and Buddleja sp. in both stages. In general, this pattern is observed in tropical environments where the highest densities of seeds belong to early successional species (Rico-Gray & García-Franco, 1992, Miller 1999, López-Toledo & Martínez-Ramos 2011, Maza-Villalobos et al. 2011). High seed density and a large number of pioneer species in the soil seed bank can contribute to the regeneration process, under natural conditions or as part of active restoration of degraded sites (Vázquez-Yanes & Orozco-Sergovia 1990, Thompson et al. 1997, Grime 2001, Ma et al. 2011).
Late successional species are mainly found at advanced successional stages, but they can also be found at early or intermediate stages (Baider et al. 2001, Maza-Villalobos et al. 2011). In the case of our study, we found tree species such as Gaultheria sp., Gaiadendron punctatum, Smilax sp., Valeriana sp., and Oreopanax sp. which were exclusive to old-growth forest. Alvarez-Aquino et al. (2005) assessed areas with different degrees of disturbance in cloud forest fragments in Mexico where the seed bank is larger and mainly dominated by tree species. We believe the absence of seeds of these species in the shrubland is a result of the isolation, size and severity of disturbance, and the regeneration strategy of the colonizing species, as has been found in other studies (Garwood 1989). Such characteristics are common in fragmented landscapes (Poschlod et al. 1996).
Viability. The observed average viability of seeds from these samples (3-30%) is consistent with levels reported in other studies in which various methods were used to determine viability (Roberts 1981, Gross 1990). Gross (1990) found, however, that the presence of large numbers of viable seeds in soil makes it difficult to draw conclusions from studies about seed densities using separation and direct counting. The low viability found with the direct separation method could be the result of fungal attack, predation (Schafer & Kotanen 2003) or simply the aging of seeds (Priestley 1986) We found differences in the number of viable seeds between successional stages; in old-growth forest, we found a greater number of viable seeds at all depths, possibly caused by litter accumulation (Facelli & Pickett 1991, Egawa & Tsuyuzaki 2013) or differences in soil type and moisture content (Bekker et al. 2000). The opposite pattern was observed in the shrubland, a fact that has been explained by low litter accumulation in the initial stages of succession; litter then gradually accumulates and increases as succession progresses (Facelli & Pickett 1991). Litter reduces light intensity and temperature fluctuations, among other micro-environmental factors (Donath & Eckstein 2010, Egawa & Tsuyuzaki 2013). This in turn may act to maintain seeds within or below the litter in an ungerminated but viable state (Wijayratne & Pyke 2012).
Although the tetrazolium viability test has been widely used and is officially recognized by the International Seed Testing Association (Hartmann & Kester 1983), this method has frequently been found to be inconsistent. This has been attributed to variation in the permeability of the seed coat (Van Waes & Debergh 1986). Moreover, the reliability of the test has been questioned given the bias associated with the preparation and interpretation of the stain (van Hezewijk et al. 1993). Although we used a stereoscope to evaluate the external structure of all seeds extracted by the method of direct separation in order to make sure they did not have apparent damage, in order to have a more accurate approximation of the structure and density of the seed bank it is necessary to evaluate other methods used to assess the viability of the seeds obtained by the method of direct separation such as the recovery rate of seeds or microscopic evaluation of the embryo, and thus avoid overestimation of seed density by this method.
Our results show that the direct separation method enables a higher density and diversity in the soil seed bank to be detected. By using the germination method, however, we recorded species that were undetected by the separation method. We therefore recommend the complementary use of these two methods in order to obtain an optimum description of the composition and structure of the soil seed bank of Andean forests. The seeds found in the soil seed banks of the two stages belong to different successional groups, a fact which could increase ecosystem resilience to disturbance. We believe one key point in the study of soil seed banks is to identify the process and mechanisms that trigger/inhibit seed germination in the forest soil and the manner in which the soil seed bank responds to different disturbances. Because recovery of the high Andean ecosystems may be limited by the propagule sources, we believe that the composition of seed banks at different depths may be essential to the natural recovery process and even to planned restoration measures.

