











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkInga inicuil Schltdl. & Cham. ex G. Don (Ingeae, Mimosoideae, Leguminosae) is a tree native to Mesoamerica where it is used as a shade tree in coffee plantations (Pennington & Sarukhán 1998). In the mountainous region of Coatepec, Veracruz, Mexico this tree and fruit are commonly known as “jinicuil”. The seeds are appreciated for their sweetness fleshy sarcotesta and the pods are sold as fresh fruit. Jinicuil is also the common name used at the coastal plains of Veracruz, where people there eat the seeds cooked and salted as a snack. The cotyledons contain protein (Geilfus 1994, Bressani 2010) and germination can reach 100 % (Pennington 1997), which gives the species high potential as a food crop. Additionally, pods are sold in local markets in Coatepec. In spite of its cultural importance, there is not any agricultural management of the tree and the rate of local loss of trees in this area has been estimated at 2.4 % annually (Pulido-Salas 2009, 2013).
One challenge in achieving agricultural sustainability is the development of agroecosystems with a predominance of native species (Altieri & Nicholls 2000). To accomplish this, it is necessary to learn more about these species (Vázquez-Yanes et al. 1999) as well as characterize them in order to design specific management programs (Martínez-Castillo et al. 2004). The diversity and genetic structure of populations constitute a basic starting point for developing management protocols for native species or for initiating genetic conservation programs. If biodiversity is not preserved, heterozygosity may be reduced and introgression can occur with the concomitant deterioration of local genetic populations and of the species itself (Baverstock & Moritz 1996). Knowing the genetic diversity of native species allows for the development of long-term programs to manage, monitor, and preserve populations (Karp et al. 1997). To achieve this, molecular approaches allow the identification of polymorphisms in different regions of the genome with various mutation rates (Parker et al. 1998). There are several molecular techniques for exploring genetic diversity. Both Amplified Fragment Length Polymorphisms (AFLPs) and Random Amplified Polymorphic DNA (RAPDs) can detect genetic variation across the entire genome. Others include Restriction Fragment Length Polymorphisms (RFLPs), Simple Sequence Repeats (SSR, microsatellites) or genic sequences (González 1998), which can detect variation in more specific regions of the genome. For example, in an AFLP study with 56 samples of 30 varieties of Macadamia spp., it was possible to quantify the degree of polymorphism in the varieties and their progenitors, and to propose a possible differentiated agricultural management (Robledo 2003). In another study, using RFLP (Restriction Fragment Length Polymorphism) markers in fragmented populations of Sorbus torminalis, a species appreciated for its fruit and wood, significant genetic variation was found that could be attributed to recent changes in silvicultural practices in Central Europe (Angelone et al. 2007). Also, a study with SSR markers for the characterization and analysis of genetic diversity in cultivars of Corylus avellana found high genetic diversity and polymorphism that was suitable for distinguishing different hazelnut lineages (Bassil et al. 2013). Furthermore, research with DNA sequence of non-coding regions has also provided insight into genetic diversity in a variety of organisms because these segments of DNA are less functionally constrained and are, therefore, more variable (González & Vovides 2002). For example, a study with the trnH-trnK non-coding region of chloroplast DNA sequence data in apricot germplasm revealed significant genetic diversity of nucleotide sequences in both haplotypic and nucleotide diversity tests. The results provided some clues for the origin of apricot species and useful information for the management of apricot genetic resources (Batnini et al. 2014).
There are few studies on genetic diversity for species of Inga, particularly for I. inicuil and I. paterno Harms. Those that exist focus mainly on the preservation of species in tropical forests, such as I. edulis (e.g. Hollingsworth et al. 2005, Dawson et al. 2008, Rollo et al. 2016), I. thibaudiana (Schierenbeck et al. 1997), and I. vera (Cruz-Neto et al. 2014). Other studies of Inga have proposed hybridization between species that grow together in coffee plantations. Pennington (1997) recorded seven species that were auto-incompatible and inter-sterile and observed that the best fruit of I. inicuil (i.e. longer pods and sweeter sarcotesta) was on trees separated by over one kilometer. The possibility of hybridization, however, has not been proven genetically.
On the other hand, chemical studies of Inga species have discovered a relationship between altitude and the amount of some chemical compounds. For example, it has been found that the amount of pipeolitic acid (a non-proteinic amino acid) in the leaves of several species varies according to altitude. Although I. inicuil was not included in that study, it was shown that species growing at a similar altitude have similar patterns of pipeolitic acid, which is interpreted as a defense strategy against predators or pathogens such as ants and fungi (Morton et al. 1991, Kite 1997). Also, Koptur (1985) reported varying concentrations of phenolic compounds in leaves of I. densiflora and I. punctata that are also correlated with altitude. Nonetheless, it is unknown if there is a connection between altitude and sugar content in the fruit of I. inicuil.
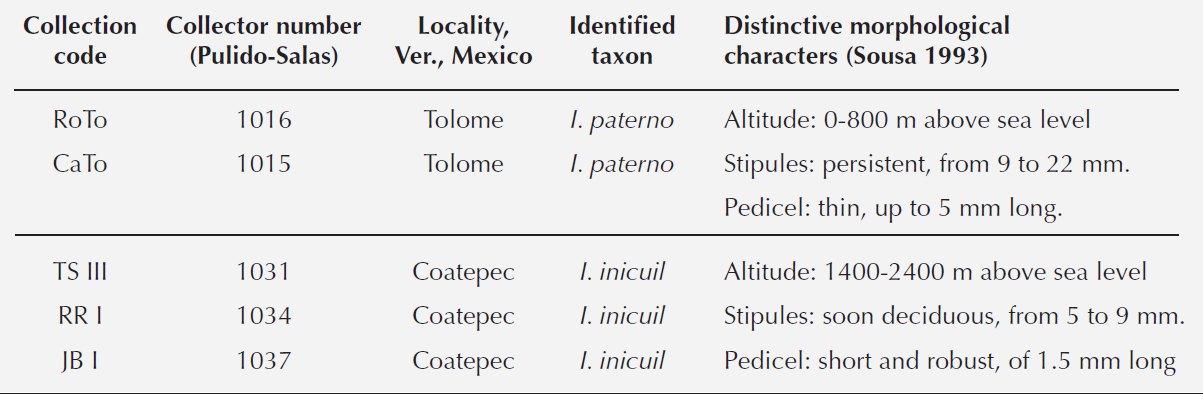
Inga has great morphological variation (Pennington 1997). It is considered a genus in the process of diversification (Richardson et al. 2001). It includes around 300 species within the tropical Americas, ranging in altitude from sea level to 3,000 m. The genus has had a complex taxonomic history and unstable nomenclature (e.g. Sousa 1993, 2001, 2009, Pennington 1997, Brown et al. 2008). Currently, two taxonomic hypotheses exist for I. inicuil. The first states that what is commonly known as “jinicuil” belongs to two species described as I. inicuil and I. paterno, with morphological differences and separated by altitude (Sousa 1993, 2001, Ricker et al. 2013). Inga paterno was described having stipules persistent, from 9 to 22 mm long, and pedicel thin, up to 5 mm long growing between 0-800 m a.s.l.; while I. inicuil has stipules soon deciduous, from 5 to 9 mm long, and pedicel short and robust, of 1.5 mm long and grows above 850 m a.s.l. The second hypothesis proposes that jinicuil is a single species (I. inicuil) with a wide altitudinal distribution, spanning from the mountainous cloud forest to the regions with evergreen and sub-deciduous forests near sea level and with wide ranges of morphological variation (Pennington 1997). Under this scenario the name I. paterno falls into synonymy with the older name I. inicuil (Pennington 1997, Pennington & Sarukhán 1998, Groom 2012). In a phylogenetic analysis with nuclear and chloroplast DNA, the genus Inga as a whole was shown to be monophyletic. However, very little variation was found in the sequences generating unresolved topologies, which led to the conclusion that Inga represents a recently diversified genus (Richardson et al. 2001). In a molecular phylogenetic analysis aimed at elucidating the evolution of defenses to herbivory in the genus, several species of Inga were analyzed (Kursar et al. 2009), and the results confirmed the lack of variation within species of Inga. None of these studies included I. inicuil or I. paterno. The aim of this study was to explore variation in sequences of non-coding regions from the chloroplast and nuclear genomes within individuals of I. inicuil from contrasting altitude. We hypothesized that phylogenetic analyses with sequence data would separate exemplars of I. inicuil in two different species and as the case with other metabolites, the sugar content in the sarcotesta would vary with altitude.
Materials and methods
Material studied. From 57 located trees, 22 adult individuals were selected (DBH > 15 cm). Trees were not physically close to each other to avoid possible kinship, and were chosen from contrasting altitude. From these, 19 were from the municipality of Coatepec, Veracruz, located between 850 and 1,530 m a.s.l. on the edge of the central mountainous region of Mexico and were used as shade trees in coffee plantations, while three were from the town of Tolome, municipality of Paso de Ovejas, Veracruz in the coastal region at 50 m a.s.l. Fertile branches/ samples from the chosen trees were collected for herbarium specimens and some were also sent to a taxonomist specialist on Inga for an accurate identification. Representative collections were deposited at the herbaria XAL, Instituto de Ecología, A. C. in Xalapa, Veracruz and MEXU, Instituto de Biología, Universidad Nacional Autónoma de México in Mexico City.
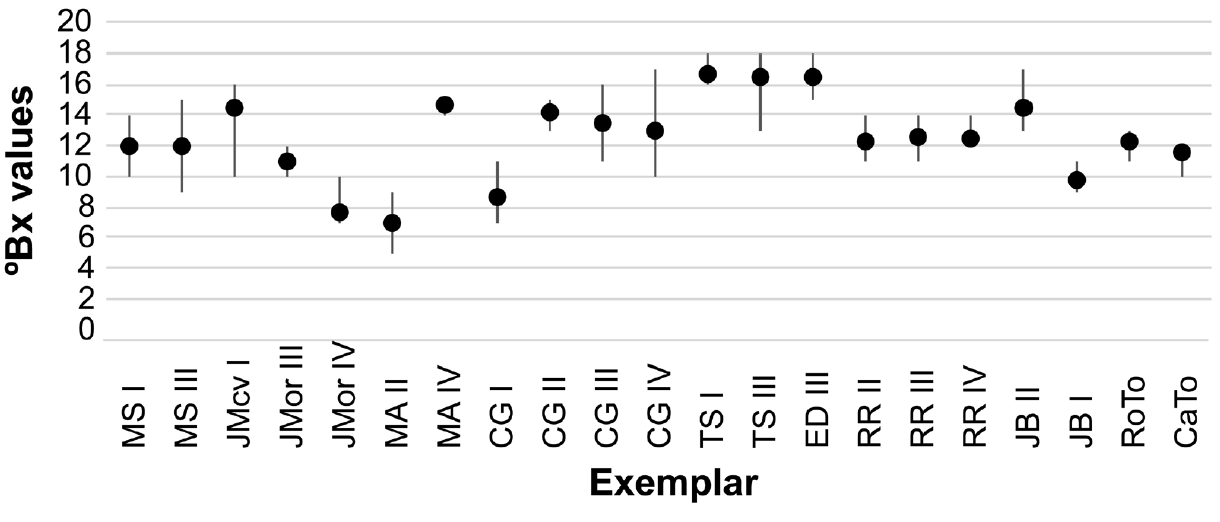
Sugar content in fruit. Given that the sweetness of the pulp covering the seeds is one of the characteristics that increases the commercial potential of I. inicuil, and with the hypothesis that, as in the case with other metabolites, there could be a connection between altitude and sugar content, we took a sample of 16 pods per tree from 19 trees from the mountain region and three from the coastal region, four for each quarter of an imaginary square on the crown (Martínez-Moreno et al. 2006). The sugar content was measured (°Bx average/tree) in ripe fruit of the juice coming out from the sarcotesta using a refractometer (ATAGO ATC-1; range 0-32 °Bx) during the summer. The four highest values per tree were used to calculate the average. Different seeds from the same pod were measured separately. The Pearson correlation coefficient was measured in Excel.
DNA extraction and amplification. For DNA extraction, 25 representative trees were selected from localities with contrasting altitude (Table 1). Twelve were from the mountainous region (municipality of Coatepec) and 13 from the coastal region (three from Tolome and ten from San Pancho, in the municipalities of Paso de Ovejas and La Antigua, respectively). Genomic DNA was extracted from young recently collected leaves, as suggested by Doyle & Doyle (1987). Extraction and purification of DNA was conducted with the DNeasy plant mini kit (Qiagen, Valencia, CA, USA), following the instructions of the manufacturer. To verify the quantity and quality of the DNA, an aliquot was taken from the extraction and loaded in a 1.2 % agarose gel dyed with ethidium bromide. A molecular-weight marker of known concentration (25 ng/μl) was included in the gel. Each sample was amplified with specific primers for two molecular markers, one nuclear and the other from chloroplast. The nuclear marker corresponded to the ITS1-5.8SITS2 region (ITS), and the chloroplast marker to a section of the trnL-F intergenic spacer. Primers used for amplifying the ITS were ITS1 (5’TCCGTAGGTGAACCTGCGG -3’) and ITS-4 (5’-TCCTCCGCTTATTGATATGC-3’; White et al. 1990). Primers for the trnL-F were “e” and “f” (5’-GGTTCAAGTCCCTCTATCCC-3’ and 5’-ATTTGAACTGGTGACACGAG-3’ respectively; Taberlet et al. 1991). Reactions were performed in a 25 μl mixture containing 10-20 ng of DNA, 5 μl of PCR buffer, 200 μM of each of the four deoxynucleoside triphosphates, 5 pmol of each primer, 2.5 mM MgCl2, 2.5 U of Go Taq flexi DNA polymerase (Promega, Madison, WI, USA), and distilled water to volume. The amplifications were performed on a thermocycler (Mastercycler, Eppendorf, Hamburg, Germany). The amplification program included an initial denaturation at 94 °C for 5 min, followed by 35 cycles with denaturation at 94 °C for 1 min, annealing at 51-59 °C for 1 min, and extension at 72°C for 2 min, and a final extension for 7 min at 72 °C.
Table 1 Municipality, altitude and GenBank accession numbers for specimens used in phylogenetic analyses. *indicates specimens with an insertion/deletion (indel) of about 309 bp in the trnL-F region.

DNA sequencing and analyses. Amplified DNA was purified before sequencing with the Wizard SV gel and PCR clean-up system kit as described by the manufacturer (Promega, Madison, WI, USA), and sequenced using ABI PRISM BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) according to manufacturer’s instructions. Cycle sequence products were cleaned with an isopropanol precipitation and electrophoresed using an ABI 310 genetic analyzer (Applied Biosystems, Foster City, CA, USA). The resulting sequences were edited with BioEdit software version 7.1.3.0 (Hall 1999) and aligned using ClustalW with default parameters for gap opening and extension penalties (Thompson et al. 1994). Sequence variation was obtained using the Arlequin software v. 3.1 (Excoffier et al. 2005). Phylogenetic analyses of sequence data were performed separately and combined with Maximum Likelihood (ML) using GARLI v. 0.951 (Zwickl 2006) and Bayesian Phylogenetic Inference (BPI) using MrBayes 3.1.2 (Huelsenbeck & Ronquist 2001, Ronquist & Huelsenbeck 2003, Altekar et al. 2004). BPI analyses were executed with model parameters GTR + Γ (nst = 6; rates = gamma), and default values for priors. ML analyses were performed specifying in the script that model parameters were computed at the same time during the searches. Analyses consisted of 10 replicates to ensure that results were consistent and reproducible. Branch support for ML was determined simultaneously by performing 100 non-parametric bootstrap iterations in each of the 10 replicates. BPI analyses comprised two independent 1-million generation runs, with four chains (one cold and three hot) each, until an average standard deviation of split frequencies of 0.01 or less was reached. We sampled trees every 100th generation and discarded initial samples applying a “burnin” value of 25 % before calculating the majority consensus tree and posterior probabilities (PP) for clades.
Four matrices were prepared for analyses. One included only trnL-F sequences from 69 terminals. They corresponded to twenty-five sequences generated in this study (12 from the mountainous region and 13 from the coastal plains), and 38 from GenBank. From the GenBank sequences, 28 correspond to identified Inga species (I. alba, I. edulis and I. thibaudiana with two, four and two representatives respectively), five to non-identified Inga spp., and six to representatives from the tribe Ingeae, which were used as the outgroup (Zapoteca sousae, Pithecellobium diversifolium, Abarema piresii, and three for Zygia spp.). Two matrices contained only ITS sequences. One comprised 464 terminals, 25 from this study and 434 from GenBank. From these, 45 correspond to identified Inga species (some species having up to 27 representatives), 52 to non-identified Inga spp., and five representatives from the outgroup. Other matrix had only 110 terminals. This included 25 from this study, two of each identified Inga species (10 species with only one) and the representatives of the tribe Ingeae. A concatenated matrix was also constructed. This included the 25 terminals from this study, two species of Inga with same voucher (I. edulis and I. thibaudiana) that have sequences in GenBank for both loci, and three representatives of the tribe Ingeae. All cladograms were edited with Adobe Illustrator v 13.0.2. Alignment and resulting trees from multi-locus gene analyses are deposited in TreeBASE (http://purl.org/phylo/treebase/phylows/study/TB2:S17506).
Results
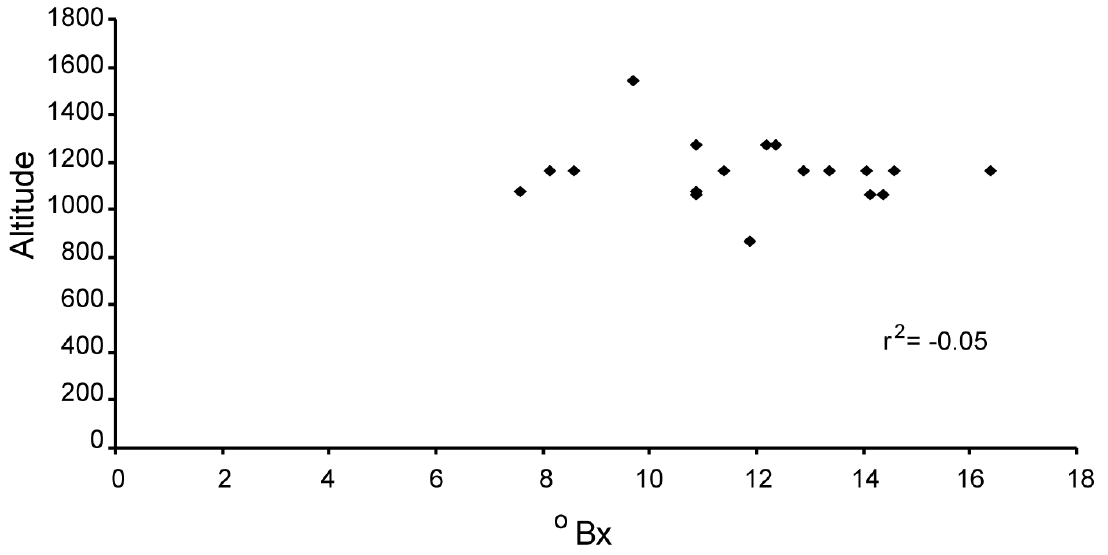
Sugar content and sequence analyses. The analysis of sugar content in the fruits revealed unexpected variation. Values obtained do not show any discernible pattern (Figure 1) and are not homogeneous among seeds even in the same pod. For example, data obtained from seeds in one of the pods tested varied from 13 to 18 °Bx. There was also no relationship of sugar content and altitude (Figure 2; r2 = -0.05) or a relationship of sugar content with DNA sequence variation.
The results for sequence data confirmed that sequences from both loci had little variation. Nevertheless, DNA sequences from ITS and trnL-F in exemplars from the coastal and mountainous regions varied according to altitude. The trnL-F region did not have any nucleotide substitution among the 25 obtained sequences, but there was an insertion/deletion (indel) of about 309 base pairs (bp) exclusively in the 13 trees from the coastal region (marked with an asterisk in Table 1). The sizes of the amplicons for the trnL-F with the primers e-f were 495 bp for the 12 exemplars from the mountain region and 186 bp for the 13 trees from the coastal region (Figure 3). This indel is not present in any of the 33 species of Inga that have sequences for this locus in GenBank. In contrast, sequences from the ITS have only nucleotide substitutions among the 25 sequences obtained in this study. The size of the amplicon for the ITS with the primers ITS1ITS4 was 657 bp (Table 2). As with the trnL-F locus, there was distinctive variation in nucleotide substitutions for the 13 exemplars from the coastal plains (Figure 4) among 437 exemplars of Inga that have ITS sequences in GenBank.
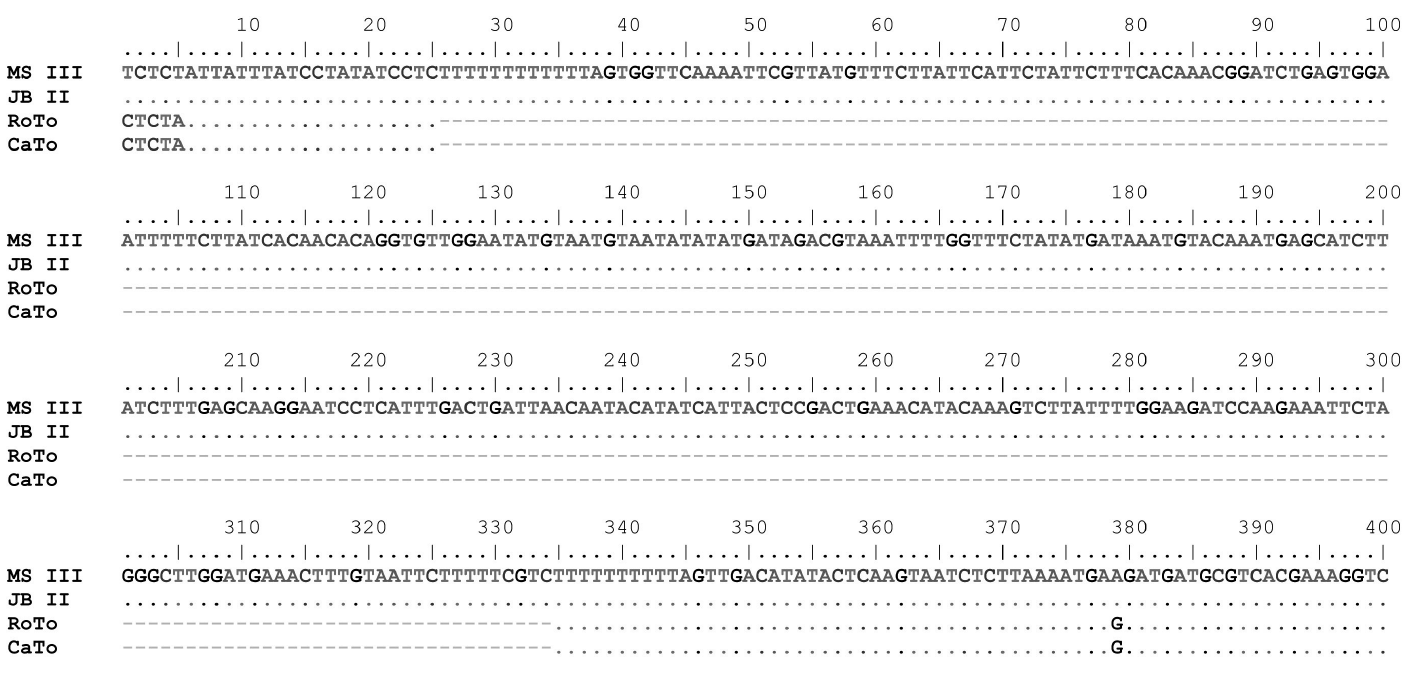
Figure 3 Sequence alignment of a section of the trnL-F gene in four exemplars of Inga showing the large indel in exemplars of the coastal plains (Roto and CaTo). MS III and JB II were collected from the mountainous region.

Figure 4 Sequence alignment of a section of the ITS gene in four exemplars of Inga showing differences in nucleotide substitutions between exemplars from the coastal plains (Roto and CaTo), and from the mountainous region (MS III and JB II).
Table 2 Summary of sequence variation in the fruit tree Inga inicuil. Most of the polymorphic sites are due to the indel in exemplars from the coastal region.
Phylogenetic analyses. ML and BPI analyses with the trnL-F locus and 69 terminals generated unresolved cladograms (Figure 5). Low DNA sequence variation was a problem for the analyses due to few characters. The best tree from the ML analysis had a log likelihood (-lnL) score of -1333.8458 and revealed a relationship between the 13 exemplars from the coastal plains and Pithecellobium diversifolium, Zapoteca sousae, and Abarema piresii. However, this result is very likely artificial since it is the result of lack of characters in the sequences with the 309 bp indel. In contrast, ML analysis with the matrix of 464 sequences for the locus ITS generated a tree with several groups in terminal branches, but without resolution in basal branches (not shown), supporting the proposed recent divergence of the species within this genus. In the second ML analysis with only 110 ITS sequences (-lnL = 2755.6728), Inga auristellae was the sister group to the 13 exemplars of the coastal plains and I. suaveolens to the 12 exemplars of the mountainous region (Figure 6). Our results indicate that the sequences generated in this study do not correspond to any of Inga species deposited in GenBank. Analyses of ML and BPI of combined sequence data showed a clear distinction between the exemplars of the coastal plains and the mountainous region. Both clades had high support values. Clade containing exemplars of the coastal plains had a bootstrap value of 98 %, and a PP of 0.98 while clade containing exemplars from the mountainous region had 100 and 1.0 respectively (Figure 7). ML analysis recovered a tree with a log likelihood score of -3106.1068. Consequently, molecular data and morphologically distinctive characters noted by the taxonomist of the genus on herbarium specimens (Table 3) show clearly that both sources of information distinguish between exemplars from the mountainous region and the coastal region, supporting the hypothesis that they are two different species.
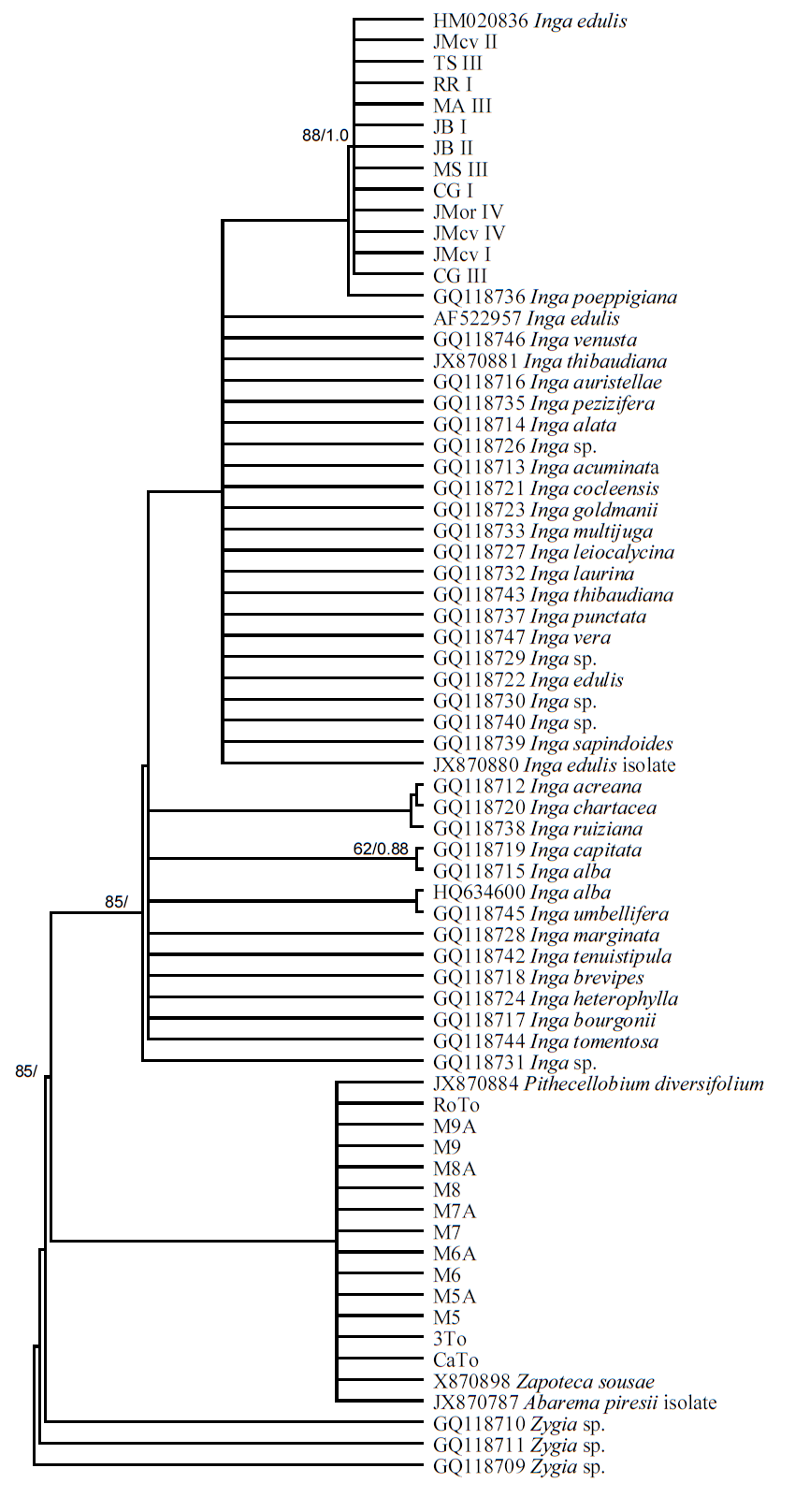
Figure 5 Phylogenetic tree for the trnL-F spacer showing the topological placement of Inga’s exemplars from the mountainous and coastal regions relative to other Inga spp. Analysis was performed with Maximum Likelihood. Values of support are indicated above branches (ML bootstrap/PP).
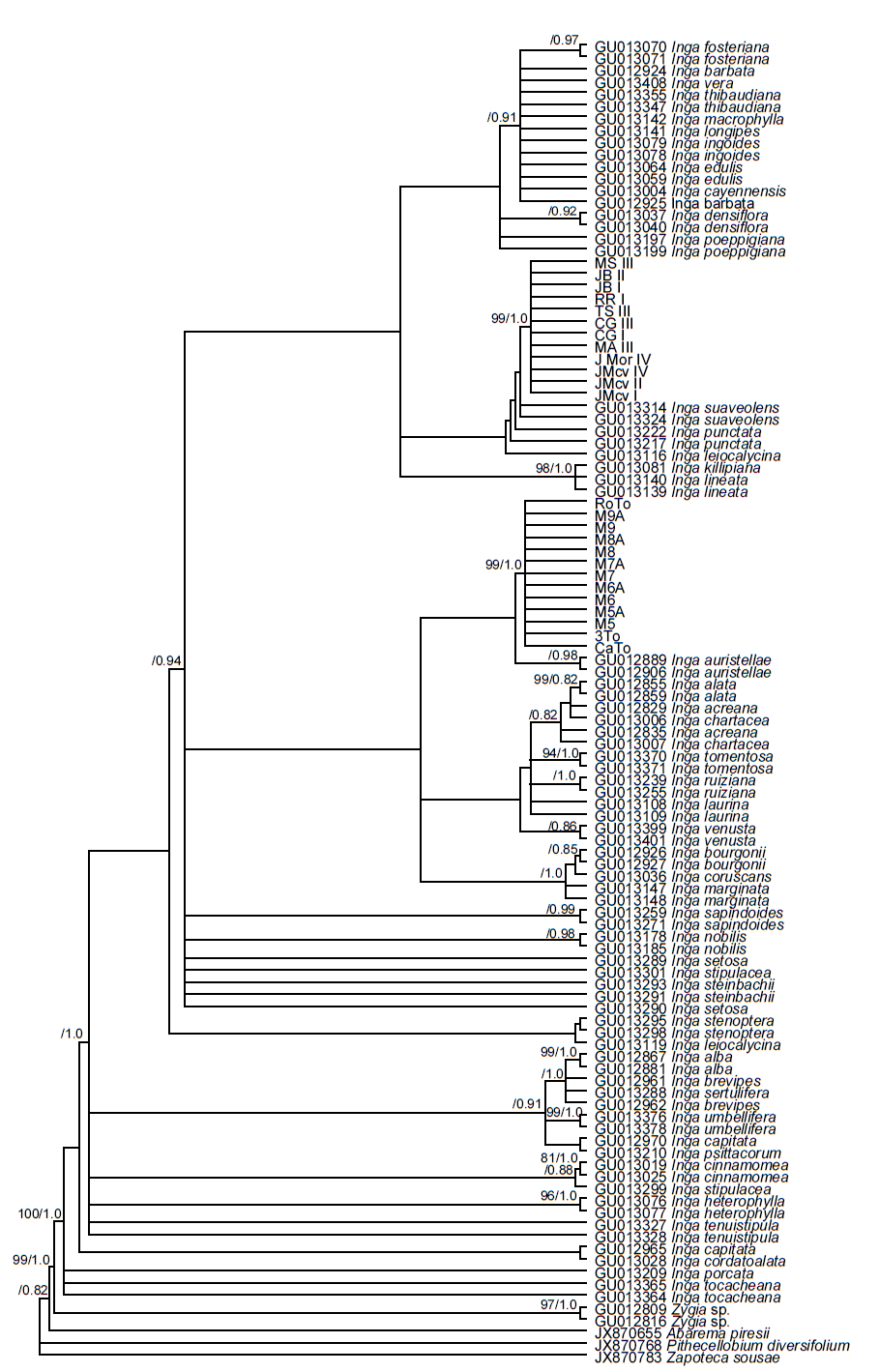
Figure 6 Phylogenetic tree for the ITS region showing the topological placement of Inga’s exemplars from the mountainous and coastal regions relative to other Inga spp. Analysis was performed with Maximum Likelihood. Values of support are indicated above branches (ML bootstrap/PP).
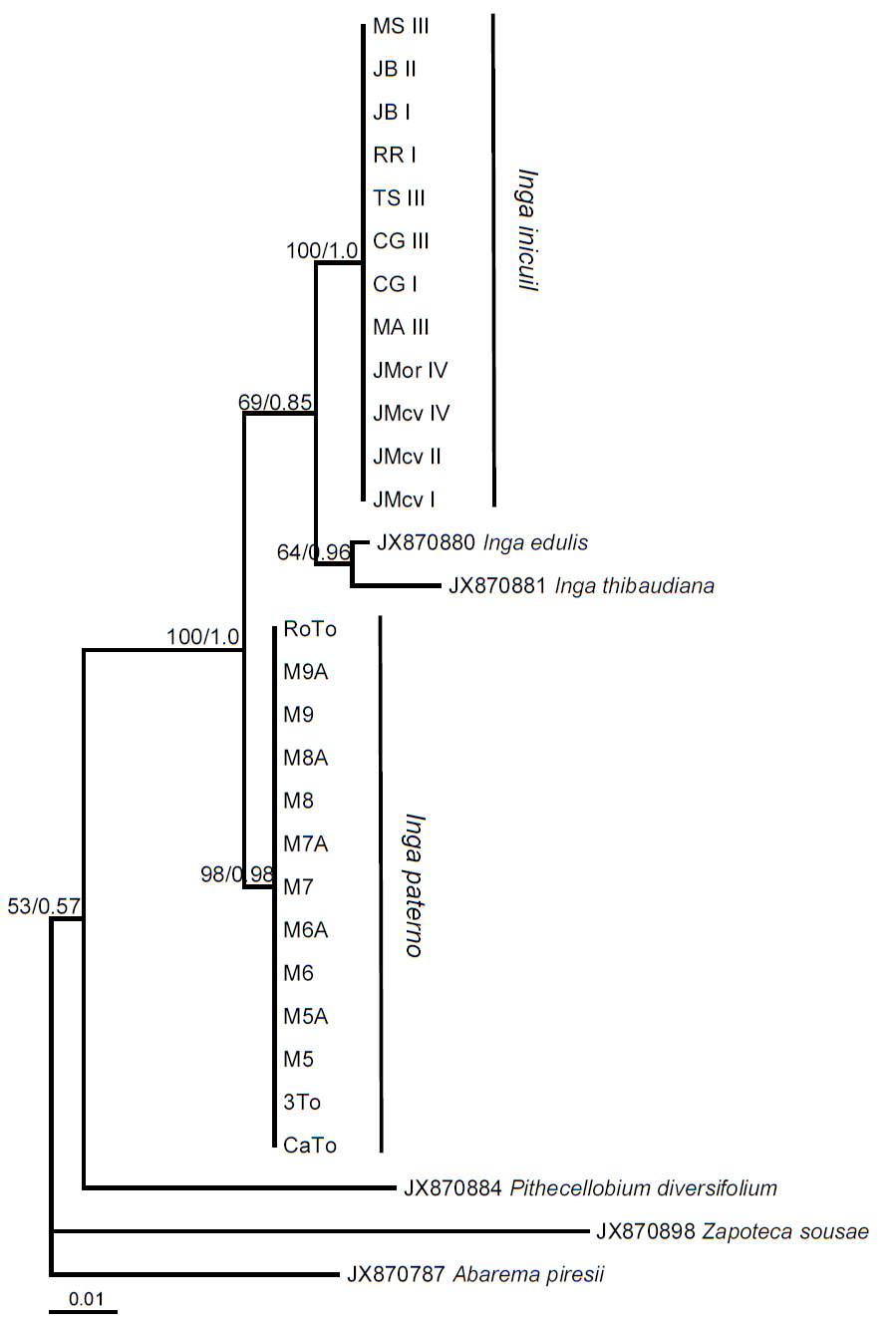
Figure 7 Hypothesis of inferred relationships of Inga inicuil and I. paterno, based on Maximum Likelihood of the concatenated data set of two loci (ITS and trnL-F). Values of support are indicated above branches (ML bootstrap/PP).
Discussion
Variation in the sugar content of the sarcotesta did not support one of our hypotheses given that there was no relationship between altitude and sugar content, as has been observed with phenolic compounds and pipeolitic acid. The possible causes for the heterogeneous distribution of sugar could due to intrinsic factors in the production of fruits in Inga similar to those observed in other species, such as auto-incompatibility, limited resources in the production of fruits, selective abortion, and issues related to the ontogenetic development of the fruit (Koptur 1983, 1984, 1985). Also, the possible spatial overlap of several species of Inga in coffee plantations may favor cross-pollination as has been suggested by Richardson et al. (2001). It can be inferred that the heterogeneity in the sugar content in the fruit shows the lack of genetic enhancement of this fruit-tree, which opens the possibility of planning genetic improvement programs for more productive use of the seeds and fruits.
Molecular variation and altitude. The variation found in the trnL-F and ITS sequences reflects two clearly defined lineages according to the altitudinal origin of the samples. DNA sequences were homogeneous in all 12 trees sampled from the mountainous region (municipality of Coatepec, Ver.); similarly, in the 13 trees from the coastal plains (Tolome and San Pancho, municipality of Paso de Ovejas and La Antigua Ver.). However, there were important dissimilarities at the nucleotide level between both groups. The conserved indel in trees from the coastal region appears to be a synapomorphic character delimiting natural lineages as has been observed in other organisms (e.g. Calviño & Downie 2007, Chiari et al. 2009, Soltis et al. 1998). Indels from the trnL-F region have resulted good phylogenetic markers for some taxa (e.g. Richardson et al. 2000, Holt et al. 2004, Ghamkhar et al. 2007, Drábková et al. 2004), as well as homoplasious for others (e.g. Kellermann & Udovicic 2007). The trnL-F region is often of different length, which can make sequence alignment and the determination of homologous bases a matter of concern in phylogenetic analyses (González et al. 2006). However, considering our results we conclude that although not devoid of homoplasy, indels can be useful markers of shared history at lower taxonomic levels as in the case of Inga inicuil and I. paterno. Differences found in this work support previous observations based on morphological characters (Sousa 1993) suggesting that I. inicuil and I. paterno are distinct species occupying habitats at different altitudes: Inga inicuil corresponds to specimens collected from Coatepec, Ver., (range 850 to 1,530 m a.s.l.), while trees sampled in Tolome and San Pancho (20-50 m a.s.l.) correspond to I. paterno (Sousa 1993). Due to the clear differences found between the trees from the mountainous versus the coastal regions, it would be useful to continue the study with a detailed ethnobotanical exploration of the uses for seeds and fruits of these species by people in localities at different altitudes. Also, the remarkable differences of sequence data from the trnL-F region within the species make further research necessary, increasing taxon sampling along with its distribution range. The study of DNA variation in what popularly is called “jinicuil” in the physiographically diverse state of Veracruz, Mexico, provides useful data that help to clarify the taxonomy of this group and which in turn, considering the uses and potential food of these species, provide a firmer basis for designing sustainable management of these native fruit-trees.

