











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkThe geological history and rugged topography of Mexico have given rise to a broad mosaic of climates and ecosystems that includes most of the biomes recognized at global level. These characteristics are reflected in exceptional biological diversity, a large part of which originated and evolved in Mexican territory (Villaseñor, 2003; Challenger and Soberón, 2008). Understanding this biodiversity is one of the great challenges for Mexico, since on this knowledge depend many ecological processes that are essential for life on the planet and for human wellbeing. These processes include climate regulation, flows of energy and elements in the biogeochemical cycles, ecosystem productivity, food production, acquisition of medicines and natural services for the control of pests, among others (Naeem et al., 1999; Cardinales et al., 2012).
The high diversification of Mexican plants can be demonstrated by the fact that the phanerogamic flora of Mexico is calculated to comprise 21,000-22,000 species, a number that represents 10-12 % of the flora worldwide and places it among the top 10 largest floras of the world (Rzedowski, 1991; Mittermeier et al., 1999; Villaseñor, 2003). Furthermore, many families and genera have their centers of diversification in Mexico (Villaseñor et al., 2007), while around 50 % of the vascular plants in Mexico and more than 200 genera are endemic to the country (Rzedowski, 1991; Delgadillo et al., 2003; Villaseñor 2003).
The flora of Mexico has been studied since pre-Hispanic times; however, such study only adopted scientific rigor from the beginning of the 20th century. From that period, there was an explosion of studies of flora, vegetation, ecology and biogeography in many regions and floristic groups and this is when an exceptional advance took place in the knowledge of the Mexican flora (Rzedowski, 2006). While there is relatively extensive current knowledge of some families (e.g. Asteraceae, Fabaceae and Poaceae), there are still large groups, such as the pteridophytes and bryophytes, of which little is known (Villaseñor et al., 2007; CONABIO, 2008). Important gaps therefore exist in the knowledge of our biodiversity and study of the Mexican flora remains a priority.
Anthropogenic activities have put this biodiversity at risk; for example, at least 26 species of plants have been lost in recent years and factors driving the loss of species are increasingly numerous and intense (CONABIO, 2008). In view of the environmental degradation of the country, there is an urgent need for robust information on which to base sound decisions regarding conservation and sustainable use. Without this information, many species will be lost and with them millions of years of evolution and the opportunity to conserve as yet unknown environmental services.
One way in which to generate knowledge of the biodiversity of plants and other biological groups is through ecological niche modeling, since this technique allows mapping of the potential distribution of species (Guisan and Thuiller, 2005; Elith and Leathwick, 2009). Information used to generate these models includes presence data, obtained from herbaria, biological collections, relevant literature or information sources that integrate data originating from primary sources (e.g., GBIF; the Global Biodiversity Information Facility) (Anderson et al., 2003; Anderson and Martínez-Meyer, 2004; Soberón and Peterson, 2009). The techniques of niche modeling add value to the presence data that have been accumulating over time, and new knowledge of great utility is created.
In Mexico, the use of these modeling techniques in plant species is not new. For example, there are studies in which the potential distribution of cactus species has been modeled (Téllez-Valdés and Dávila-Aranda, 2003), as well as species of the genus Jefea (Asteraceae) (Villaseñor and Téllez-Valdés, 2004), Polianthes (Asparagaceae) (Solano and Feria, 2007) and for the families Asteracea, Fabaceae and Poaceae (Villaseñor et al., 2007) as well as for 190 species of Mexican tropical trees (Pennington and Sarukhán, 2005), among others.
Advances in the development of niche modeling methodologies and the level of knowledge currently held in Mexico motivated CONABIO to adopt niche modeling as a tool with which to advance the knowledge of biodiversity. In this context, this institution is currently supporting the development of 20 research projects, which are expected to produce distribution models for approximately 20 % of the plants of Mexico (Fig. 1). This strategy began with a workshop conducted by Townsend Peterson and Jorge Soberón from the Department of Ecology & Evolutionary Biology of the University of Kansas, aimed at the researchers and technical staff of these projects. This workshop focused on disseminating the methodologies and in reaching consensus regarding the best practices of modeling. The information generated from this initiative shares a common methodological base, including a detailed taxonomic and geographic depuration conducted by experts and the definition of a ‘modeling region’ according to biological and biogeographic criteria, in which the expert knowledge deployed is reflected. Other recommended practices include the implementation of methods to avoid collinearity among selected variables, definition of clear and transparent criteria to define the binary maps of distribution of each species and statistical evaluation of the performance of each model. These practices confer greater reliability in the use of this information for different purposes, such as the planning of new scientific explorations. This initiative will produce an important advance in the knowledge of species of the Mexican flora. Of around 24,950 terrestrial vascular plant species in Mexico, including Gymnosperms, Angiosperms and Ferns/Allies, around 5,250 species will have a reliable model of distribution (Fig. 1).
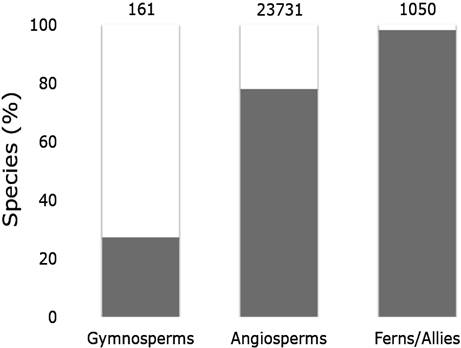
Figure 1 Percentage (and number) of species from different plant groups with (open bars) and without (closed bars) distribution models, following the CONABIO initiative.
The initiative of presenting a special section in this edition of Botanical Sciences arose from the symposium “Modeling the distribution of Mexican plants: knowledge and challenges in the face of biodiversity loss” organized by Leonel Lopez-Toledo and Pilar Rodríguez during the XIX Mexican Botanical Congress, in Tuxtla Gutiérrez, Chiapas, Mexico in November 2013, following a detailed process of peer review evaluation and editing.
The section includes three articles arising from the projects supported by CONABIO. These include detailed studies of trees, shrubs, herbs and epiphytes; species that inhabit temperate as well as warm-humid environments, and studies in which different modeling methods are utilized, using contemporary and/or future scenarios and considering different climate change models.
Martínez-Méndez and collaborators (2016) develop ecological niche models for eight species of Abies with an evolutionary and conservationist focus. Through comparison of the ecological niches of the species, these authors explore whether evolutionary divergences are also reflected at ecological level and identify priority areas for species conservation. The use of static models utilizing climatic variables for modeling species distribution is analyzed in Vaca and Golicher (2016), who conclude that this approach is insufficient. In contrast, these authors propose the development of mechanistic models that utilize variables that integrate the physiological characteristics of the organisms. With this approach, the authors develop models for 258 tree species of different ecosystems that vary in terms of degree of exposure to periods of drought.
Garza-López et al. (2016) demonstrate that, assuming climate change, the climatic habitat of Swietenia macrophylla will practically disappear in a near future in Quintana Roo and will be displaced toward the West. The authors propose a strategy of ex-situ conservation in order to recouple the populations to their future suitable climate. Additionally, in a previous edition of this journal a work of Saénz and collaborators (2015) was published. In that article, the authors demonstrate the use of an ecological niche approach in the modeling of species under future climate change scenarios. These authors develop models for Pinus leiophylla showing how the zones of greatest environmental suitability for this species will differ in the future.
The studies included in this special section, together with that of Sáenz and collaborators (Botanical Sciences 93:709-718, 2015), are a small sample of the current research and possible applications of niche modeling towards understanding the biodiversity and evolution of plants and other organisms. It is gratifying to witness the continued growth of an academic community involved in these research lines, since a megadiverse country such as Mexico requires the continual development of strategies and new tools in order to advance the knowledge of its biodiversity. Among these multiple strategies, there is no doubt that niche modeling is an approach with great potential.

