










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBotanical Sciences
versión On-line ISSN 2007-4476versión impresa ISSN 2007-4298
Bot. sci vol.93 no.1 México mar. 2015
https://doi.org/10.17129/botsci.233
Ecología
Espectro de dispersión de la flora leñosa del bosque tropical caducifolio en el Neotrópico
Dispersal spectrum of woody flora in Neotropical dry forests
Libertad Arredondo-Amezcua1, Leonel López-Toledo2 y Guillermo Ibarra-Manríquez1,3
1 Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, Morelia, Michoacán, México.
2 Instituto de Investigaciones sobre los Recursos Naturales, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, México.
3 Autor por correspondencia: gibarra@cieco.unam.mx.
Recibido: 30 de noviembre de 2013.
Aceptado: 9 de marzo de 2014.
Resumen: Mediante una revisión de información bibliográfica y de herbario, se analizó el espectro de dispersión de árboles y lianas de localidades con bosque tropical caducifolio en América, con el objetivo de determinar si este atributo se modifica al considerar diferentes conjuntos florísticos, como floras leñosas regionales o parcelas de muestreo de 0.1 ha. Para estas últimas, se cuantificó el grado de similitud del espectro de dispersión, obtenido mediante el conteo de especies o por el número de individuos. A una escala local, las proporciones de los síndromes de dispersión fueron similares entre la flora y lo obtenido en los muestreos, pero los síndromes de dispersión difirieron entre sí de manera significativa. A nivel Neotropical, el análisis entre tres floras también varió significativamente entre síndromes. Las comparaciones entre parcelas mostraron que existen diferencias significativas entre localidades, tanto entre síndromes de dispersión como entre la interacción síndrome-localidad. Este resultado persiste al analizar formas de crecimiento de manera independiente o considerando su abundancia. Las divergencias encontradas entre localidades no fueron explicadas por la latitud o variables climáticas; pero tanto a nivel local como Neotropical, estas son más claras para las formas de crecimiento, ya que la zoocoria es más importante para árboles y la anemocoria para lianas. Los resultados constatan la complejidad del fenómeno de la dispersión de especies leñosas del bosque tropical caducifolio, por lo que futuros estudios deberán indagar con más detalle la influencia de la escala espacial, así como factores proximales e históricos.
Palabras clave: árboles, Chamela, lianas, síndromes de dispersión.
Abstract: Through a bibliographic and herbarium review, an analysis was conducted of the dispersal spectrum of woody plants in various locations of the tropical deciduous forest of America. Our aim was to determine whether modifications to the dispersal spectrum exist among different floristic groups, particularly in the floristic checklist or within plots of 0.1 ha. The degree of similarity among dispersal spectra was estimated, considering the number of species or individuals. At the local scale, percentages of dispersal syndromes did not differ among the flora obtained by sampling, but the dispersal syndromes differed significantly from each other. At the Neotropical level, the dispersal syndromes of three floras also differed significantly. At plot level, different sites also differed significantly in terms of dispersal syndromes and in syndrome-site interactions. This result persisted on analysis of different growth forms or when considering abundance. Inter-site differences were not explained by latitude or climatic variables. At both local and Neotropical levels, the differences were more evident in terms of growth form, since trees and lianas were more zoochorous and anemochorous, respectively. This study demonstrates the complexity of dispersal in woody species of the tropical deciduous forest, and further studies should explore the influence of spatial scale, as well as proximal and historic factors.
Key words: Chamela, dispersal syndromes, lianas, trees.
Al comparar distintos espectros de dispersión del bosque tropical caducifolio en el Neotrópico, se ha detectado que la diseminación por animales (zoocoria) y por viento (anemocoria) son los síndromes mejor representados (Gottsberger y Silberbauer-Gottsberger, 1983; Wikander, 1984; Gentry, 1995; Machado et al., 1997; Gillespie, 1999; Grizy Machado, 2001; Chain-Guadarrama, 2005; Jara-Guerrero et al., 2011). Para el caso particular de los árboles, la zoocoria es el más frecuente, con valores que oscilan entre 45 y 65.7%, seguido por la anemocoria, que engloba entre 23 y 38% de las especies (Frankie et al., 1974; Gentry, 1995; Gillespie, 1999; Arredondo-Amezcua, 2004; Chain-Guadarrama, 2005; Jara-Guerrero et al., 2011). Sin embargo, se deben considerar los resultados de Wikander (1984), y Griz y Machado (2001), quienes concuerdan en señalar una importancia equivalente para ambos mecanismos (cerca de 30%), para los árboles de bosques tropicales secos de Charavalle (Venezuela) y de Caatinga (Brasil). La tendencia para las lianas es más clara, ya que independientemente de la localidad en estudio, la mayoría de sus miembros son anemócoros (60-75%), en tanto que la segunda categoría es ocupada por la dispersión zoócora, con porcentajes que van desde 18 hasta 25% (Wikander, 1984; Gentry, 1995; Gillespie, 1999; Griz y Machado, 2001; Solórzano et al., 2002). Sin embargo, es importante destacar que las comparaciones entre estos estudios no pueden ser tan directas, dado que se utilizaron distintos métodos de muestreo.
Por otro lado, se ha señalado que factores ambientales relacionados con la humedad, influyen fuertemente en la diversidad y aspectos funcionales de las plantas del bosque tropical caducifolio (Gentry, 1982, 1995; Wikander, 1984; Griz y Machado, 2001). En ese sentido, se ha detectado una sincronía particular entre la maduración de los frutos y semillas con las condiciones ambientales adecuadas para su dispersión, lo que ha relacionado a la zoocoria con áreas o temporadas del año con mayor humedad y a la anemocoria con condiciones de sequía (Frankie et al., 1974; Griz y Machado, 2001; Gottsberger y Silberbauer-Gottsberger, 1983; Wikander, 1984; Bullock y Solis-Magallanes, 1990; Bullock, 1995; Oliveira y Moreira, 1992; Justiniano y Fredericksen, 2000; Griz y Machado, 2001; Jara-Guerrero et al., 2011). Sin embargo, no se ha explorado la posible existencia de un patrón al respecto a lo largo del Neotrópico.
Una pregunta que ha sido escasamente abordada y para la que los pocos estudios previos muestran resultados controvertidos, es si las proporciones en que se presentan los mecanismos de dispersión podrían cambiar al evaluarlos con base en la abundancia de las especies y no solo considerando la presencia de estas en una comunidad particular. Jara-Guerrero et al. (2011) indican que para un bosque tropical caducifolio en Ecuador, la zoocoria representa el mecanismo predominante; mientras que si se considera la abundancia, la anemocoria resulta ser el más importante. Previamente, Gillespie (1999), realizando esta comparación para siete comunidades en Centroamérica y, Chain-Guadarrama (2005), haciendo lo propio en dos localidades en México, no encontraron diferencias al respecto.
Otro aspecto que es importante destacar, es que la mayoría de los estudios de dispersión mencionados previamente, se basan en el censo de parcelas y no en inventarios florísticos completos. Un cuestionamiento inmediato, sobre este punto, es si los resultados obtenidos de ambos diseños de estudio difieren respecto a su espectro de dispersión. El único antecedente conocido es el de Gentry (1982), quien no encontró diferencias en este atributo al comparar los árboles y lianas censados en parcelas de 0.1 ha, con respecto a lo obtenido para toda la flora de un bosque tropical caducifolio (Parque Santa Rosa, Costa Rica), y la de dos bosques tropicales húmedos (Barro Colorado, Panamá; Río Palenque, Ecuador).
Tomando en cuenta los antecedentes expuestos previamente, el presente estudio se propuso responder las siguientes preguntas: (1) ¿existen diferencias en los espectros de dispersión de los árboles y lianas obtenidos en parcelas de muestreo, con respecto al que presenta el total de especies registradas en una localidad particular?, (2) ¿en qué medida, el espectro de dispersión difiere entre comunidades de bosque tropical caducifolio del Neotrópico?, (3) ¿el espectro de dispersión se modifica al tomarse en cuenta la abundancia de las especies?, y finalmente, (4) ¿la proporción en que se presentan los patrones de dispersión en los bosques caducifolios Neotropicales se asocia con factores climáticos y geográficos?
Para ello, se utilizaron tres enfoques de análisis. Se eligió la Estación de Biología Chamela, Jalisco, México, para realizar una comparación a nivel local, donde se contrastó el espectro de dispersión, obtenido mediante una revisión de ejemplares de herbario, de las especies registradas para la flora leñosa y las censadas en tres parcelas de 0.1 ha. Un segundo enfoque contrastó los espectros de dispersión reportados para dos floras ubicadas en México y una de Costa Rica. Finalmente, un análisis a nivel Neotropical utilizó la información de los espectros de dispersión publicada para diversas localidades a lo largo de la distribución del bosque tropical caducifolio en América.
Materiales y Métodos
Ecosistema y localidades de estudio. El bosque tropical caducifolio se desarrolla entre los 20° y 10° de latitud al norte y sur del Ecuador, en regiones de clima cálido, con una larga estación seca que puede variar entre cuatro y siete meses, y una precipitación anual menor a 2,000 mm (Murphy y Lugo, 1986; Rzedowski, 1978; Linares-Palomino et al., 2011). Este ecosistema es dominado por especies arborescentes que pierden sus hojas en la época seca del año durante un lapso variable, pero que por lo general oscila alrededor de seis meses (Rzedowski, 1978). En el Neotrópico, representa el 22% de las áreas boscosas en Sudamérica y casi el 50% en Centroamérica (Murphy y Lugo, 1986; Linares-Palomino et al., 2011). Al igual que el bosque tropical húmedo y lluvioso, en el bosque tropical caducifolio, las familias de mayor importancia son Fabaceae para los árboles y Bignoniaceae para las lianas, y aunque es menos diverso que sus contrapartes más húmedas, se caracteriza por su alto nivel de endemismos (Gentry, 1995). Las localidades de bosque tropical caducifolio incluidas en el presente trabajo se distribuyen a lo largo de la costa del Pacífico, desde México hasta Perú (Figura 1).
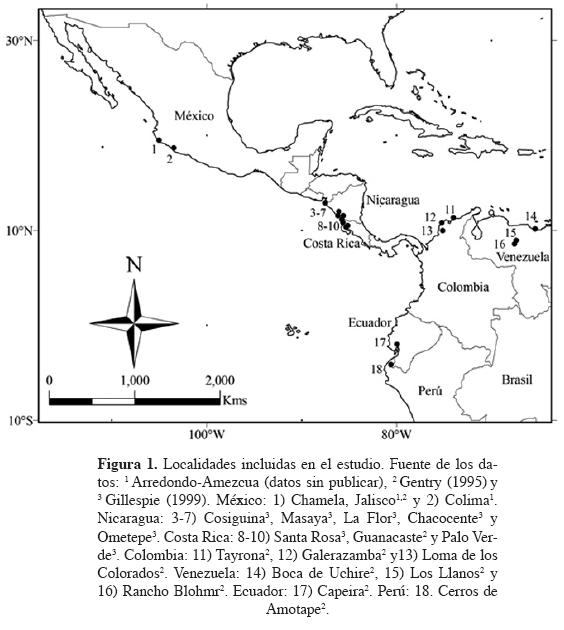
Obtención de la información. El análisis a nivel local se realizó para Chamela, México, ya que de acuerdo con Gentry (1995), representa una de las localidades más diversas del bosque tropical caducifolio a lo largo del Neotrópico y se caracteriza por ser un sitio cuya flora leñosa está adecuadamente caracterizada (Lott y Atkinson, 2002). Chamela cuenta con un listado florístico detallado (Lott, 2002) y tres parcelas censadas con el método de muestreo de Gentry, que consiste en muestrear todas las plantas leñosas ≥ de 2.5 cm de diámetro a la altura del pecho (D.A.P.) enraizadas en diez transectos de 50 × 2 m (0.1 ha), los cuales se distribuyen paralelamente y que se separan entre sí por una distancia de 20 m (Gentry, 1982). La flora leñosa de Chamela se obtuvo del listado florístico de Lott (2002) para el bosque tropical caducifolio, considerando los árboles y arbustos (agrupados en la categoría árbol), y trepadoras leñosas y lianas (agrupadas en la categoría liana), y se complementó con otras especies referidas por Gentry (1995), mientras que las tres parcelas muestreadas con el método de Gentry (1982), corresponden a los trabajos de Lott et al. (1987) y Gentry (1995), y sus bases de datos fueron obtenidas del Alwyn H. Gentry Forest Transect Data Set (Missouri Botanical Garden, 1995-2013).
A cada especie se le asignó un síndrome de dispersión, que de acuerdo a van der Pijl (1972), corresponde a las modificaciones morfológicas de frutos y/o semillas para facilitar su dispersión, y que corresponden a los mencionados recurrentemente en estudios realizados en el bosque tropical caducifolio (Gentry, 1982, 1995; Wikander, 1984; Gillespie, 1999; Griz y Machado, 2001; Arredondo-Amezcua, 2004; Chain-Guadarrama, 2005). De esta manera, la zoocoria se asoció a frutos carnosos o semillas con apéndices suculentos, como arilos o eliosomas; la anemocoria se relacionó con estructuras ligeras, de tamaño diminuto o con la presencia de apéndices como alas o pelos que les permitirían flotar usando corrientes de viento; finalmente, la autocoria se asignó a especies que tienen la posibilidad de expulsar o depositar sus diásporas de manera autónoma, por lo que éstas no presentan modificaciones específicas o son esparcidas básicamente por gravedad (van der Pijl, 1972).
Los síndromes de dispersión se designaron con información obtenida por medio de la consulta de literatura florístico-taxonómica (p.ej. Bravo-Hollis, 1978; Chiang, 1984; McVaugh, 1987; Arias et al., 1997; Avendaño, 1998; Niembro, 1998; Pennington y Sarukhán, 1998; Martínez et al., 2002; Daniel y Acosta, 2003; Diego-Pérez, 2004) y revisión de ejemplares depositados en el herbario IEB, perteneciente al Instituto de Ecología, A.C. y en el Herbario Nacional (MEXU), de la Universidad Nacional Autónoma de México, examinando frutos de ejemplares (p. ej., frutos ya dehiscentes) o utilizando la información en torno a la morfología de los frutos que los colectores del ejemplar aportaban en la etiqueta. De las 445 especies que comprenden la flora leñosa de Chamela, se definió el síndrome de dispersión para el 88.9%.
Los análisis a nivel Neotropical consideraron tres floras leñosas y diversas localidades cuyos espectros de dispersión han sido determinados en trabajos anteriores (Gentry, 1995; Gillespie, 1999; Arredondo-Amezcua, datos sin publicar), y que en el caso de los muestreos, fueron realizados con el método de Gentry (1982), descrito previamente. Se contrastó el espectro de dispersión entre las floras de Chamela, Jalisco y del estado de Colima, México (Arredondo-Amezcua, datos sin publicar), así como la de Santa Rosa, Costa Rica (Gentry, 1982). Para el caso de muestreos basados en el método de Gentry (1982), las parcelas de 0.1 ha contrastadas corresponden a 18 localidades para los árboles, 11 para las lianas y 11 que incluyen ambas formas de crecimiento; un total de nueve localidades fueron incluidas al considerar la abundancia por especie, las cuales consideran los árboles y lianas (Arredondo-Amezcua, datos sin publicar; Gentry, 1995; Gillespie, 1999). La comparación de los muestreos excluyó la autocoria debido a que Gentry (1995) no precisa sus proporciones.
Análisis de datos. Las pruebas estadísticas se hicieron mediante modelos lineales generalizados (GLM) realizados con el programa R version 2.14.0 (R Development Core Team, 2012). Para lo anterior, se definió un error binomial debido a que la comparación se llevó a cabo utilizando las proporciones de la abundancia, dado que estas son diferentes entre localidades. Los factores a probar fueron sitio (S), síndrome de dispersión (SD) y la interacción entre ambos (S:SD). La variable de respuesta fue la proporción del número de especies o de individuos para cada síndrome, haciendo análisis por separado para árboles, lianas y leñosas (la suma de ambas).
A nivel Neotropical, también se llevó a cabo un análisis para explorar si la latitud o algunos factores climáticos afectan el espectro de dispersión (medido como la abundancia o la proporción de la zoocoria y anemocoria). Esta relación se exploró mediante regresiones lineales, utilizando un modelo lineal generalizado, con error Poisson para el caso de la abundancia y un error binomial para el caso de la proporción (Faraway, 2005; Crawley, 2007). En los GLM, la devianza obtenida para cada factor explica la variación dada por este y se aproxima a una distribución de χ2. Los valores de esta distribución son los que indican la significancia estadística y son los reportados en los resultados (Crawley, 2007).
Específicamente, se probó la relación entre la abundancia y la proporción de cada síndrome con la latitud y cinco variables climáticas: (1) Temperatura promedio, (2) temperatura máxima, (3) precipitación total, (4) número de meses secos y (5) la precipitación en la temporada de sequía. Para el caso de las variables climáticas, el análisis únicamente se llevó a cabo utilizando los datos de estaciones meteorológicas de siete localidades que cuentan con esta información. Para ello, se hizo una búsqueda de estaciones cercanas a las localidades incluidas en las comparaciones Neotropicales, en los sitios web de meteorología nacionales de los países correspondientes (p.ej. Instituto Meteorológico Nacional de Costa Rica o Sistema de Meteorología de la Aviación de Venezuela).
Resultados
Análisis local. Para Chamela, la zoocoria y la autocoria fueron los síndromes mejor representados, con valores que oscilan entre 28 a 48%, independientemente si se analizan simultáneamente las lianas y los árboles, o únicamente estos últimos (Figura 2), en tanto que el síndrome de dispersión con mayor importancia para las lianas fue la anemocoria, con valores entre 50 y 75% (Figura 2). No se detectaron diferencias significativas entre el espectro de dispersión de la flora y el obtenido en las tres parcelas de 0.1 ha, ya sea si se analizan los árboles, las lianas o ambas formas de crecimiento en conjunto (Cuadro 1, Figura 2). Por el contrario, la proporción de los tres síndromes de dispersión sí fue significativamente diferente (Cuadro 1, Figura 2).


Tomando en cuenta la abundancia de las especies, se encontró que los árboles tienen valores similares entre zoocoria y autocoria, al igual que lo observado en el análisis de la proporción de especies (Cuadro 1, Figuras 2, 3). Para el caso de las lianas, las proporciones de los síndromes varían entre parcelas pero la anemocoria sigue siendo el que presenta un mayor porcentaje (Cuadro 1, Figura 3). Al analizar las abundancias de árboles y lianas en conjunto no se encontraron diferencias significativas entre parcelas ni entre los síndromes de dispersión (Cuadro 1, Figura 3).

Análisis Neotropical. En la comparación entre las floras, la zoocoria fue el síndrome más importante, con valores de 39.4 y 58.7%. Un porcentaje mayor de especies autócoras se obtuvo en las floras más norteñas (Chamela, 32.9%; Colima, 24.2%), mientras que en Santa Rosa sólo alcanzó el 8.1%. A pesar de que el espectro de dispersión entre las floras mostró variación significativa entre los tres síndromes, no se detectaron diferencias entre sitios o en la interacción entre ambos factores (Cuadro 2).

Al contrastar el espectro de dispersión de distintas localidades por medio de muestreos (Figura 4), la zoocoria predominó en los árboles para la mayor parte de los sitios; excepto en Colima, La Flor y Capeira, donde los porcentajes de entre este síndrome y la anemocoria fueron similares, oscilando entre 44 y 56%. Para el caso de las lianas (Figura 4), este último síndrome resultó el preponderante, llegando a tener hasta 100% en las localidades de Guanacaste y Tayrona; sólo Galerazamba presentó porcentajes similares para los dos síndromes de dispersión comparados (anemocoria 47% y zoocoria 53%). Tomando en cuenta simultáneamente los árboles y lianas, la devianza explicada, tanto por el síndrome de dispersión como por la interacción sitio-síndrome, fueron significativas, siendo este último tipo de análisis el que explicó la mayor parte de la devianza (Cuadro 2).

Esto obedece a que las proporciones de cada síndrome de dispersión en las distintas localidades es contrastante y conforma tres principales tendencias (Figura 4): (1) cuatro localidades presentaron porcentajes más altos de anemocoria, de 59 a 69% (Guanacaste, Tayrona, Los Llanos y Capeira), (2) Chamela y Galerazamba tuvieron una mayor proporción de zoocoria (64 y 71%, respectivamente) y (3) cinco localidades comparten valores similares entre ambos síndromes, los cuales oscilan entre 44 y 56% (Colima, Boca de Uchire, Rancho Blohmr, Loma los Colorados y Amotape).
En la comparación de los espectros de dispersión, tomando en cuenta el número de individuos por especie, el análisis de nueve localidades indica que la mayor parte de la devianza es explicada por la interacción y en menor medida, por el síndrome de dispersión, siendo ambas pruebas significativas (Cuadro 2). La zoocoria fue el síndrome más representativo en cinco sitios, fluctuando de 60 a 76% (Cosiguina, Masaya, Chacocente, Ometepe y Palo Verde). Por el contrario, Chamela, Colima y Santa Rosa tuvieron valores similares entre zoocoria y anemocoria, mientras que La Flor fue la única localidad donde este último síndrome fue el más importante (Figura 5).

En general, es más alta la proporción de especies dispersadas por viento en sitios más cercanos al Ecuador (hasta con 64.9%) y disminuye en latitudes mayores; sin embargo, esta relación no fue significativa (r = 0.38, P = 0.1). Por el contrario, la zoocoria no presentó ninguna tendencia con respecto a la ubicación latitudinal de los sitios (r = 0.22, P = 0.2). Tampoco se encontró ninguna relación entre las variables climáticas y los síndromes de dispersión, independientemente de si se analiza abundancia y/o la proporción de las especies.
Discusión
Análisis local. Las proporciones en que se presentan los síndromes de dispersión entre las parcelas y la flora leñosa de Chamela no mostraron diferencias significativas entre sí (Cuadro 1), un resultado similar al encontrado por Gentry (1982), quien comparó tres floras y varios sitios muestreados de 0.1 ha en Centro y Sudamérica. Estos resultados indican que la dispersión de las leñosas de Chamela puede ser abordado al analizar la flora en su totalidad o mediante un muestreo de un sitio dado. Este patrón no se modifica si se toma en cuenta la abundancia de las especies.
En Chamela se detectó una proporción alta de especies dispersadas por autocoria (Cuadro 1, Figura 2). La importancia de este síndrome puede ser considerada respecto a su posible asociación a la zoocoria como parte de un complejo proceso de dispersión. Por ejemplo, Pirie et al. (2000) encontraron que las diásporas de dos especies autócoras de Chamela, Jatropha standleyi y J. chamelensis (Euphorbiaceae), son acarreadas por roedores después de caer al suelo, y su destino final está íntimamente relacionado con el comportamiento de estos mamíferos, quienes llevan las semillas a varios metros de distancia de la planta progenitora. Estudios en otras localidades de bosques tropicales han documentado resultados similares (Hallwachs, 1986; Adler y Kestell, 1998; Brewer y Rejmánek, 1999). Debido a lo anterior, Vander Wall et al. (2005) consideran que la caída de las diásporas es parte de un proceso de presentación ante el vector de dispersión y que los animales que llevan después las semillas serían realmente los dispersores.
Por otra parte, las características que han sido señaladas como típicas de diásporas dispersadas por animales (Dansereau y Lems, 1957; van der Pijl, 1972), excluyen a los frutos secos tipo nueces o semillas, sin apéndices carnosos, a pesar de que en ambos casos son objeto de forrajeo por parte de animales granívoros, los cuales, aunque actúan principalmente como depredadores, también pueden ser dispersores efectivos. Un caso típico de esto es el de las semillas pequeñas (sin eliosomas) que son dispersadas por hormigas granívoras (Álvarez-Sánchez y Naranjo-García, 2003).
Debido a que dichas diásporas no presentan modificaciones aparentes para el proceso de diseminación, es difícil clasificarlas como zoócoras sin observar este proceso in situ, por lo que la categoría autocoria, en muchas ocasiones, puede resultar ambigua. Estos resultados apuntan la necesidad de realizar trabajo de campo para corroborar los síndromes de dispersión, especialmente para aquellas diásporas incluidas en esta última categoría.
Respecto a las formas de crecimiento, para Chamela la zoocoria resultó importante para los árboles, de manera similar a lo indicado para otros bosques tropicales caducifolios en el Neotrópico (Frankie et al., 1974; Gentry, 1995; Gillespie, 1999; Arredondo-Amezcua, 2004; Chain-Guadarrama, 2005), mientras que la anemocoria lo fue para las lianas (Wikander, 1984; Gentry, 1995; Gillespie, 1999; Griz y Machado, 2001; Solórzano et al., 2002). Ambos patrones fueron consistentes, tanto si fueron determinados por la proporción del número de sus especies o por su abundancia.
Análisis Neotropical. El análisis de las floras de las localidades de Chamela, Colima y Santa Rosa mostró a la zoocoria como el síndrome preponderante en el bosque tropical caducifolio, tal como lo propone Gentry (1982). Por otro lado, el incremento en el porcentaje de autocoria para Chamela y Colima comparada con Santa Rosa sustentan las observaciones de Gentry (1995), de que dicho síndrome aumenta en la medida que se avanza hacia el extremo más norteño de la zona Neotropical, lo que se atribuye principalmente al incremento en la diversidad de Euphorbiaceae. Esta afirmación es apoyada por los trabajos florísticos realizados en Chamela (Lott y Atkinson, 2002) y Colima (Padilla-Velarde et al., 2006), ya que la riqueza de especies de esta familia solo es superada por Fabaceae.
De acuerdo con Gentry (1982), la proporción de especies dispersadas por viento para los árboles y lianas es mayor en el bosque tropical caducifolio, que el registrado para sus contrapartes menos estacionales, como el bosque tropical lluvioso o el bosque tropical húmedo, lo cual se explica principalmente por las diferencias en la precipitación que estos ecosistemas presentan. En el presente estudio, al considerar varias localidades del bosque tropical caducifolio se obtuvieron valores promedio de 28% para los árboles y 72.8% para las lianas. Estos porcentajes son cercanos a los obtenidos por Gentry (1982), ya que para la primera de estas formas de crecimiento indica un valor medio entre 30 y 40% en bosques tropicales caducifolios, lo que contrasta con los valores de 15 y 6% en bosques tropicales lluviosos y húmedos, respectivamente. Para el caso de las lianas, en los dos últimos tipos de vegetación, este autor indica porcentajes de 60 y 48%, respectivamente, los cuales están por debajo de lo registrado en el bosque tropical caducifolio (81%).
Contrariamente a lo esperado, la dispersión entre las localidades Neotropicales, combinando las formas de crecimiento analizadas, varió desde porcentajes semejantes de zoocoria y anemocoria, hasta la predominancia de alguno de estos síndromes (Cuadro 2). Esto puede deberse a diversas causas como la topografía, el tamaño del fragmento de bosque donde se lleve a cabo el muestreo o la humedad relacionada con la precipitación total o meses de lluvia. Por ejemplo, respecto a la anemocoria, de acuerdo con Wikander (1984), las velocidades de las corrientes de aire que se presentan en la temporada de defoliación en Charavalle, Venezuela, particularmente en las colinas, generan condiciones que facilitan la dispersión por viento, sobre todo de las especies que alcanzan los estratos más altos o medios de la vegetación, o que están apostadas en las cimas. Por otro lado, Gillespie (1999) encontró que en fragmentos pequeños de bosque tropical caducifolio, la proporción de especies transportadas por mamíferos disminuye. En cuanto a la humedad, Bullock (1995) discutió que el aumento de la diseminación por animales es más común en regiones donde la precipitación total anual es mayor que 1,000 mm. Relacionado con este factor, se ha encontrado que la zoocoria incrementa en la medida que lo hace el número de meses de lluvias (Machado et al., 1997).
En el presente estudio no se encontró ninguna relación significativa respecto al posible efecto de factores climáticos sobre el espectro de dispersión. Esto puede obedecer, por un lado, a que el análisis se realizó para pocas localidades, ya que únicamente siete contaban con datos climáticos. Por otra parte, aunque alguno o varios de estos factores climáticos podrían explicar los procesos de dispersión a nivel local, no parecen ser suficientes para entender las diferencias encontradas entre las localidades Neotropicales a escalas más amplias. Por ejemplo, cuando se comparan sitios con una duración similar de meses de sequía (en este caso correspondiente a seis meses), se tiene que Galerazamba, con 500 mm de precipitación anual, resultó mayoritariamente zoócora; mientras que Charavalle, con una mayor precipitación (957 mm), presentó valores equivalentes entre zoocoria y anemocoria. Lo anterior contrasta con Tayrona, cuya flora es predominantemente anemócora, pero con un registro más alto de precipitación que los dos sitios previamente mencionados (1,500 mm).
De acuerdo con Gentry (1995), las proporciones de diseminación por animales que se encontraron en Galerazamba, a pesar de la fuerte sequía de la localidad, están relacionadas más con la inusual diversidad de la familia Capparaceae, especialista de selvas muy secas. Además, géneros como Bursera (Burseraceae), Caesalpinia y Lonchocarpus (Fabaceae), de alta abundancia en algunos sitios del bosque tropical caducifolio, pueden ejercer influencia en la proporción de los síndromes de dispersión. Dados estos hallazgos, resulta importante realizar análisis biogeográficos de los taxa para tener una mejor comprensión de los patrones de dispersión de las comunidades vegetales (Gentry, 1995; Brown y Lomolino, 1998; Lavin y Matos, 2008), ya que diversos estudios han mostrado que la variación que muestran los frutos a nivel de especie (p.ej., tamaño o vectores de dispersión con los que se asocian), tiene una relación significativa con los géneros o familias a las que pertenecen, conservando preferencias ecológicas ancestrales (Ibarra-Manríquez y Oyama, 1992; Willson y Traveset, 2000; Ibarra-Manríquez et al., 2001; Webb et al., 2002; Chazdon et al., 2003; Bolmgren y Eriksson, 2005).
Por otra parte, en los bosques Neotropicales caducifolios parece existir un patrón en relación con las formas de crecimiento. Para los árboles, la zoocoria fue el síndrome de dispersión más importante, incluso al tomar en cuenta la abundancia de las especies. En el análisis del número de especies, las excepciones fueron Colima, La Flor y Capeira, que obtuvieron un porcentaje de zoocoria similar a la anemocoria. Cuando se contrastan las abundancias, La Flor cambió de tendencia, predominando la anemocoria con 79%, mientras que Chamela, Colima y Santa Rosa obtuvieron una proporción similar entre síndromes. Por otra parte, para las lianas, el síndrome mejor representado fue la anemocoria, excepto en Galerazamba, donde la zoocoria tuvo la misma proporción. A pesar de las excepciones previamente mencionadas, la predominancia de la zoocoria en árboles y de la anemocoria en lianas, son patrones congruentes con lo documentado en otras localidades de bosque tropical caducifolio en el continente americano (Frankie et al., 1974; Oliveira y Moreira, 1992; Bullock, 1995; Machado et al., 1997; Griz y Machado, 2001).
Más allá de las complicaciones y ambigüedades que representa el estudio de la diseminación a partir de la morfología de las diásporas, este tipo de enfoque resulta útil como una primera aproximación al complejo proceso de la dispersión a nivel comunitario y, de acuerdo a los resultados obtenidos en este trabajo, parece ser más consistente para realizar comparaciones entre formas de crecimiento. Sin embargo, para entender de manera más integral la dispersión de las especies del bosque tropical caducifolio, futuros estudios deberán realizar trabajo de campo para verificar si los síndromes asignados son correctos, particularmente en el caso de la autocoria, e incorporar necesariamente el peso que tienen, a diferentes escalas espaciales, los factores ecológicos e históricos.
Agradecimientos
Se agradece al programa de maestría del Posgrado en Ciencias Biológicas de la Universidad Nacional Autónoma de México y al aporte económico de la beca con registro 193222 del Consejo Nacional de Ciencia y Tecnología (CONACyT), otorgada a la primera autora, que hicieron posible esta investigación. Se reconocen las revisiones y aportaciones de Ellen Andresen, Katherine Renton, Jorge Arturo Meave del Castillo, Mauricio Quesada Avendaño, Miguel Martínez Ramos y dos revisores anónimos, que mejoraron los análisis y diversas partes del manuscrito. Finalmente, se agradece también al Missouri Botanical Garden por el acceso a las bases de datos sustento de este trabajo y al M. en C. Juan Martínez Cruz, quien también compartió información para analizar la flora leñosa de Colima.
Literatura citada
Adler G.H. y Kestell D.W. 1998. Fates of Neotropical tree seeds influenced by spiny rats (Proechimys semispinosus). Biotropica 30:677-681. [ Links ]
Álvarez-Sánchez J. y Naranjo-García E. Eds. 2003. Ecología del Suelo en una Selva Tropical Húmeda de México. Instituto de Ecología A. C., Instituto de Biología, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Arias M.S., Gama L.S y Guzmán C.L.U. 1997. Cactaceae. Flora del Valle de Tehuacán-Cuicatlán 14:1-146. [ Links ]
Arredondo-Amezcua L. 2004.Tipos de dispersión de los árboles de la selva baja caducifolia de Colima, México. Tesis de Licenciatura, Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo, Morelia. 56 pp. [ Links ]
Avendaño R.S. 1998. Bombacaceae. Flora de Veracruz 107:1-40. [ Links ]
Bolmgren K. y Eriksson O. 2005. Fleshy fruits -origins, niche shifts, and diversification. Oikos 109:255-272. [ Links ]
Bravo-Hollis H. 1978. Las Cactáceas de México. Volumen I. Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Brewer S.W. y Rejmánek M. 1999. Small rodents are significant dispersers of tree seeds in a Neotropical forest. Journal of Vegetation Science 10:165-174. [ Links ]
Brown J.H. y Lomolino M.V. 1998. Biogeography. 2a ed. Sinauer Associates, Sunderland. [ Links ]
Bullock S.H. 1995. Plant reproduction in Neotropical dry forests. En: Bullock S.H., Money H.A. y Medina E. Eds. Seasonally Dry Tropical Forests, pp. 277-303, Cambridge University Press, Cambridge. [ Links ]
Bullock S.H. y Solis-Magallanes J.A. 1990. Phenology of canopy trees of a deciduous forest in Mexico. Biotropica 22:22-35. [ Links ]
Chain-Guadarrama A. 2005. Síndromes de dispersión en el mosaico vegetacional de la región de Nizanda (Oaxaca), México. Tesis licenciatura, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F. 67 pp. [ Links ]
Chazdon R.L., Careaga S., Webb C. y Vargas O. 2003. Community and phylogenetic structure of reproductive traits of woody species in wet tropical forests. Ecological Monographs 73:331-348. [ Links ]
Chiang F. 1984. A new species of Helietta (Rutaceae) from Guerrero and Jalisco, Mexico; a synopsis of the North American species. Brittonia 36:455-457. [ Links ]
Crawley M.J. 2007. The R Book. John Wiley & Sons Ltd., Chichester. [ Links ]
Daniel T.F. y Acosta C.S. 2003. Familia Acanthaceae. Flora del Bajío y Regiones Adyacentes 117:1-173. [ Links ]
Dansereau P. y Lems K. 1957. The grading of dispersal types in plant communities and their ecological significance. Contributions to the Institute of Botany, University of Montreal 71:1-52. [ Links ]
Diego-Pérez N. 2004. Apocynaceae. Flora de Guerrero 20:1-117. [ Links ]
Faraway J.J. 2005. Extending the Linear Model with R: Generalized Linear, Mixed Effects and Nonparametric Regression Models, Chapman & Hall, CRC, Boca Raton. [ Links ]
Frankie G.W., Baker H.G. y Opler P.A. 1974. Comparative phenological studies of trees in tropical wet and dry forests in the lowlands of Costa Rica. Journal of Ecology 62:881-919. [ Links ]
Gentry A.H. 1982. Patterns of Neotropical plants species diversity. Evolutionary Biology 15:1-84. [ Links ]
Gentry A.H. 1995. Diversity and floristic composition of Neotropical dry forests. En: Bullock S.H., Money H.A. y Medina E. Eds. Seasonally Dry Tropical Forests, pp. 146-194, Cambridge University Press, Cambridge. [ Links ]
Gillespie T.W. 1999. Life history characteristics and rarity of woody plants in tropical dry forest fragments of Central America. Journal of Tropical Ecology 15:637-649. [ Links ]
Gottsberger G. y Silberbauer-Gottsberger I. 1983. Dispersal and distribution in the Cerrado vegetation of Brazil. Sonderbaende des Naturwissenschaftlichen Vereins in Hamburg 7:315-352. [ Links ]
Griz L.M.S. y Machado I.C.S. 2001. Fruiting phenology and seed dispersal syndromes in Caatinga, a tropical dry forest in the northeast of Brazil. Journal of Tropical Ecology 17:303-321. [ Links ]
Hallwachs W. 1986. Agoutis (Dasyprocta punctata): the inheritors of guapinol (Hymenaea courbaril, Leguminosae). En: Estrada A. y Fleming T.H. Eds. Frugivores and Seed Dispersal, pp 285-304, Dr. W. Junk Publishers, Dordrecht. [ Links ]
Ibarra-Manríquez G. y Oyama K. 1992. Ecological correlates of reproductive traits of Mexican rain forest trees. American Journal of Botany 79:383-394. [ Links ]
Ibarra-Manríquez G., Martínez-Ramos M. y Oyama K. 2001. Seedling functional types in a lowland rain forest in Mexico. American Journal Botany 88:1801-1812. [ Links ]
Jara-Guerrero A., De la Cruz M. y Méndez M. 2011. Seed dispersal spectrum of woody species in south Ecuadorian dry forests: environmental correlates and the effect of considering species abundance. Biotropica 43:722-730. [ Links ]
Justiniano M.J. y Fredericksen T.S. 2000. Phenology of tree species in Bolivian dry forests. Biotropica 32:276-281. [ Links ]
Lavin M. y Matos A.B. 2008. The impact of ecology and biogeography on legume diversity, endemism, and phylogeny in the Caribbean region: a new direction in historical biogeography. The Botanical Review 74:178-196. [ Links ]
Linares-Palomino R., Oliveira-Filho A.T. y Pennington R.T. 2011. Neotropical seasonally dry forests: diversity, endemism, and biogeography of woody plants. En: Dirzo R., Young H.S., Mooney H.A. y Ceballos G. Eds. Seasonally Dry Tropical Forests. Ecology and Conservation, pp. 3-22, Island Press, Washington, D.C. [ Links ]
Lott E.J. 2002. Lista anotada de las plantas vasculares de Chamela-Cuixmala. En: Noguera F.A., Vega R.J.H., García A.A.N. y Quesada A.M. Comp. Historia Natural de Chamela, pp. 99-136, Instituto de Biología, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Lott E.J. y Atkinson T.H. 2002. Biodiversidad y fitogeografía de Chamela-Cuixmala, Jalisco. En: Noguera F.A., Vega R.J.H., García A.A.N. y Quesada A.M. Comp. Historia Natural de Chamela, pp. 83-97, Instituto de Biología, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Lott E.J., Bullock S.H. y Solís-Magallanes J.A. 1987. Floristic diversity and structure of upland and arroyo forests of coastal Jalisco. Biotropica 19:228-235. [ Links ]
Machado I.C.S., Barros L.M. y Sampaio E.V.S.B. 1997. Phenology of Caatinga species at Serra Talhada, PE, northeastern Brazil. Biotropica 29:57-68. [ Links ]
Martínez G.M., Jiménez R.J., Cruz D.R., Juárez A.E., García R., Cervantes A. y Mejía H.R. 2002. Los géneros de la familia Euphorbiaceae en México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Botánica 73:155-281. [ Links ]
McVaugh R. 1987. Leguminosae. Flora Novo-Galiciana 5:1-786. [ Links ]
Missouri Botanical Garden. 1995-2013. The Alwin H. Gentry forest transect data set. <http://www.mobot.org/mobot/research/gentry/welcome.shtml> (consultado 21 marzo 2006). [ Links ]
Murphy P.G. y Lugo A.E. 1986. Ecology of tropical dry forest. Annual Review of Ecology and Systematics 17:67-88. [ Links ]
Niembro R.A. 1998. Semillas de Árboles y Arbustos. Ontogenia y Estructura. Limusa, México, D.F. [ Links ]
Oliveira P.E.A.M. de y Moreira A.G. 1992. Anemocoria em espécies de cerrado e mata de galeria de Brasilia, D.F. Revista Brasileira de Botánica 15:163-174. [ Links ]
Padilla-Velarde E., Cuevas-Guzmán R., Ibarra-Manríquez G. y Moreno-Gómez S. 2006. Riqueza y biogeografía de la flora arbórea del estado de Colima, México. Revista Mexicana de Biodiversidad 77:271-295. [ Links ]
Pennington T.D. y Sarukhán J. 1998. Árboles Tropicales de México: Manual para la Identificación de las Principales Especies. Instituto de Ecología, Universidad Nacional Autónoma de México, Fondo de Cultura Económica, México, D.F. [ Links ]
Pirie C.D., Walmsley S., Ingle R., Pérez J.A., Solís M.A. y Kelly C.K. 2000. Investigations in plant commonness and rarity: a comparison of seed removal patterns in the widespread Jatropha standleyi and the endemic J. chamelensis (Euphorbiaceae). Biological Journal of the Linnean Society 71:501-512. [ Links ]
R Development Core Team. 2012. R ver. 2.14.0: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna. [ Links ]
Rzedowski J. 1978. Vegetación de México. Limusa, México, D.F. [ Links ]
Solórzano S., Ibarra-Manríquez G. y Oyama K. 2002. Liana diversity and reproductive attributes in two tropical forests in Mexico. Biodiversity and Conservation 11:197-212. [ Links ]
van der Pijl L. 1972. Principles of Dispersal in Higher Plants. 2a ed. Springer-Verlag, Nueva York. [ Links ]
Vander Wall S.B., Forget P.M, Lambert J.E. y Hulme P.E. 2005. Seed fate pathways: filling the gap between parent and offspring. En: Forget P.M., Lambert J.E, Hulme P.E. y Vander Wall S.B. Eds. Seed Fate. Predation, Dispersal and Seedling Establishment, pp. 1-8, CAB International, Wallingford. [ Links ]
Webb C.O., Ackerly D.D., McPeek M.A. y Donoghue M.J. 2002. Phylogenies and community ecology. Annual Review of Ecology and Systematics 33:475-505. [ Links ]
Wikander T. 1984. Mecanismos de dispersión de diásporas de una selva decidua en Venezuela. Biotropica 16:276-283. [ Links ]
Willson M. F. y Traveset A. 2000.The ecology of seed dispersal. En: Fenner M. Ed. Seeds: The Ecology of Regeneration in Plant Communities, 2a ed., pp. 85-110, CAB International, Wallingford. [ Links ]

