










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBotanical Sciences
versión On-line ISSN 2007-4476versión impresa ISSN 2007-4298
Bot. sci vol.92 no.4 México dic. 2014
Ecología
Distribución potencial de ocho especies exóticas de carácter invasor en el estado de Baja California, México
Potential distribution to eight alien species with invasive nature in the state of Baja California, Mexico
Sara Palma-Ordaz1 y José Delgadillo-Rodríguez2
1 Programa de Posgrado, Centro Iberoamericano de la Biodiversidad (CIBIO), Instituto Universitario de Investigación, Universidad de Alicante, Alicante, España.
2 Herbario BCMEX, Facultad de Ciencias, Universidad Autónoma de Baja California, Ensenada, Baja California, México.
1 Autor para la correspondencia: sara_palmah@hotmail.com.
Recibido: 19 de septiembre de 2013.
Aceptado: 24 de enero de 2014.
Resumen
Se determinó la distribución potencial de ocho especies exóticas de carácter invasor en el estado de Baja California (Atriplex semibaccata, Brassica tournefortii, Bromus rubens, Centaurea melitensis, Cynodon dactylon, Salsola tragus, Schismus barbatus y Tamarix ramosissima), mediante el uso del programa de modelaje MaxEnt. Para ello se utilizaron 19 variables climáticas y la distribución conocida de las especies, obtenida de registros de herbario. Todos los modelos fueron evaluados para probar su habilidad de predicción y se analizaron las variables climáticas más importantes en el modelo predictivo de cada especie. Se identificó la zona costera noroeste (zona mediterránea), como el área de mayor probabilidad de presencia de las especies. Esto puede deberse a que es una zona en donde históricamente el intercambio florístico ha sido más intensivo y donde la actividad antropogénica es mayor; además, probablemente estas especies han tenido una mejor adaptación a las condiciones climáticas de la zona, sólo algunas de ellas extienden su distribución potencial hacia las regiones desérticas del estado. Este es el primer trabajo que se realiza en Baja California, que considera las especies exóticas y su potencial invasor a partir de modelos de distribución potencial. Estos modelos, definidos con la información actual disponible, pueden apoyar para determinar los sitios más importantes de concentración de las mismas.
Palabras clave: distribución potencial, MaxEnt, plantas exóticas, plantas invasivas, variables climáticas.
Abstract
The potential distribution was determined for eight exotic species which are invasive in Baja California (Atriplex semibaccata, Brassica tournefortii, Bromus rubens, Centaurea melitensis, Cynodon dactylon, Salsola tragus, Schismus barbatus, and Tamarix ramosissima) with MaxEnt modeling program. To do this we used 19 climate variables and the known distribution of the species obtained from herbarium records. All models were evaluated to test their predictive ability and analyzed the most important climate variables in the predictive model for each species. The coastal region of the northwest (Mediterranean area) was identified as the area of the highest probability of occurrence for the species studied. This may be because it is a region where historically the floristic exchange (mainly from the European Mediterranean region) has been more intensive and where anthropogenic activity is greater. Probably, these species have also had a better adaptation to the climatic conditions of this area and some extending their distribution to more arid areas (desert) of the state. This is the first study done in Baja California considering exotic species and their invasive potential distribution by models. These models, defined with the current information available, can assist in determining the most important concentration sites of these species.
Keywords: climatic variables, exotic plants, invasive plants, MaxEnt, potential distribution.
Los seres humanos han causado cambios sin precedentes en los ecosistemas de todo el planeta y han redistribuido las especies vegetales y animales de forma voluntaria o accidental, lo cual tiene un impacto en la historia biológica del planeta, en ocasiones esto ha llevado a grandes problemas ecológicos (Villaseñor y Magaña, 2006). La colonización y el comercio permitieron la dispersión de muchas plantas útiles, pero también trasladaron de manera involuntaria otras plantas, frecuentemente como maleza o contaminantes de sus productos (Mack, 1991).
Las plantas introducidas o exóticas se han convertido en un componente importante de la vegetación de casi todos los países del mundo (Weber, 1997). Aunque muchas plantas han sido introducidas en otros lugares que no son su origen, solo el 10% de las especies exóticas llega a naturalizarse, y de estas, el 10% a su vez, llegan a ser invasoras (Williamson, 1996). Las etapas en el proceso de invasión de las plantas exóticas son complejas y los procesos representan un continuo. La naturalización es un requisito fundamental para la invasión de una planta. Esta se produce cuando las barreras abióticas y bióticas para la supervivencia y reproducción son superadas. Las plantas naturalizadas pueden convertirse en invasoras en nuevos hábitats solo cuando producen una adecuada descendencia reproductiva (Richardson et al., 2000).
Las plantas exóticas invasoras pueden causar daños tanto ecológicos como económicos; estas pueden afectar directamente a las plantas nativas al convertirse en monopolizadoras o donadoras de recursos limitantes. Además pueden indirectamente afectar a las plantas nativas y causar cambios en los ecosistemas mediante alteraciones en la estabilidad del suelo, promoviendo la erosión, colonizando los sustratos abiertos, afectando la acumulación de basura, sal, u otros recursos del suelo y promoviendo o suprimiendo el fuego (Vitousek, 1990; Richardson et al. 2000). Los efectos causados pueden ser dramáticos cuando alteran los regímenes de perturbación más allá del rango de variación a la cual las especies nativas están adaptadas (D'Antonio et al., 1999), lo que resulta en cambios en la comunidad y transformaciones a nivel de ecosistemas (Mack y D'Antonio, 1998).
Importancia del uso de modelos de distribución de especies. Debido al impacto potencial de las especies invasoras sobre la estructura de las comunidades biológicas y al funcionamiento de los ecosistemas, se hace necesario conocer su distribución para implementar acciones de manejo y control. Un enfoque fundamental para la comprensión y gestión de especies invasoras es determinar su distribución potencial. Varios trabajos proporcionan una visión general de la modelación de distribución de especies o una comparación de los métodos de modelación (Fielding y Bell, 1997; Guisan y Zimmermann, 2000; Zaniewski et al., 2002; Anderson et al., 2003; Segurado y Araújo, 2004; Guisan y Thuiller, 2005). Estos modelos de distribución de especies tienen por objeto predecir áreas que describan las condiciones ambientales adecuadas para la supervivencia de las especies; es decir, la distribución potencial o nicho fundamental (Anderson et al., 2003; Guisan y Thuiller, 2005). En general, estos métodos de modelación combinan datos de localidades georeferenciadas de las especies donde ha sido confirmada su presencia con variables ambientales, para crear un modelo de requerimientos de la especie de acuerdo a las variables examinadas (Anderson et al., 2003). El modelo resultante es proyectado sobre un mapa de la región de estudio que muestra la distribución potencial de las especies estudiadas. Estos mapas pueden servir para detectar áreas donde las especies invasoras pueden estar presentes y donde posiblemente estarán en el futuro. El clima es un factor importante que afecta o determina la distribución de los organismos; por tal motivo, los análisis de variables climáticas ayudan a comprender el por qué una especie crece en un determinado sitio y no en otro (Lindenmayer et al., 1991).
Origen y estudios de la flora exótica en México. En cuanto al origen de la flora exótica en México, Villaseñor y Espinosa-García (2004), comentan que es muy diverso; la mayor parte (78.9%) proviene del viejo mundo (África, Asia y Europa) y se relaciona con los siglos de colonización y dominación española; posteriormente, están Sudamérica, Oceanía y Norteamérica. En el caso de algunas plantas, su origen no está determinado. Rzedowski y Calderón (1990), refieren que durante el siglo XX hubo un notable incremento de plantas exóticas de origen africano, con una predominancia de la familia Poaceae, en su mayoría se comportan como ruderales y por lo regular son elementos escapados de cultivo. Muchas, si no la mayoría, fueron introducidas a propósito como ornamentales, forrajeras, medicinales y para césped; entre ellas, varias que se estudian aquí.
En el listado de plantas exóticas para México, realizado por Villaseñor y Espinosa-García (2004), se encuentran 618 especies que pertenecen a 355 géneros y 87 familias. Las cuatro familias que registraron el mayor número de especies exóticas fueron: Poaceae, 74 géneros y 171 especies; Fabaceae, 36 géneros y 57 especies; Asteraceae, 39 géneros y 56 especies; y Brassicaceae con 25 géneros y 45 especies. Sin embargo, en datos recientes (Villaseñor, datos no publicados), elevan la cifra del número de especies exóticas a 665, la dinámica de descubrimientos de especies exóticas es aún sorprendente, por lo que no se puede descartar que en fechas próximas las cifras tengan que ser reevaluadas.
Con relación a estudios específicos en algunas zonas del estado de Baja California en torno a especies exóticas, Soto-Hopkins (1987) analizó las malezas en el cultivo de tomate en la región de San Quintín, B.C., registró 16 familias y 45 especies, todas ellas introducidas, estando en primer lugar las gramíneas. Garcillán et al. (2009) registraron en zonas urbanas de la ciudad de Ensenada, B.C., 33 familias, 122 géneros y 161 especies, de las cuales 98 (61%) fueron no nativas. Mientras que Villaseñor y Espinosa-García (2004) estimaron para Baja California 219 especies exóticas.
Avances en México con relación al tema de especies exóticas invasoras. Actualmente no se sabe con certeza cuántas especies exóticas invasoras de todos los grupos de organismos están establecidas en México, tampoco es conocida su distribución y el tamaño de sus poblaciones, pero como parte de los compromisos adquiridos por México en el Convenio sobre Diversidad Biológica (CDB) y la Estrategia Nacional de Biodiversidad de México (ENBM), y en respuesta a los diferentes retos que representan las invasiones biológicas, la Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT), identificó la necesidad de crear la Estrategia Nacional sobre Especies Invasoras en México: prevención, control y erradicación; para ello se instauró el Comité Asesor Nacional sobre Especies Invasoras, conformado por académicos especialistas en el tema, organizaciones civiles y representantes de diversos sectores del gobierno federal que aportaron su conocimiento y experiencia para consolidar esta herramienta de planificación para México, siendo la Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad (CONABIO) la encargada de elaborar la referida estrategia, la cual fue presentada en el 2010. La visión es lograr que en el año 2020, el país cuente con sistemas eficientes de prevención, detección y respuesta temprana, así como con instrumentos que operen dentro de un marco legal congruente y conforme a las necesidades de prevención, mitigación, control y erradicación de las especies invasoras (CONABIO, 2010). De manera preliminar, el Sistema Nacional sobre Especies Invasoras de la CONABIO ha generado un listado en donde incluye a 133 plantas exóticas invasoras, este se encuentra en continua actualización, las especies registradas no constituyen un listado exhaustivo, la intención es servir como una guía.
Baja California no cuenta con una lista concreta acerca de las plantas exóticas invasoras. Actualmente, en el trabajo de Garcillán et al. (2013), se presenta un listado exhaustivo de la flora no nativa naturalizada para toda la península de Baja California, en donde también identificaron a 18 especies (8% del total) con comportamiento invasor en la península, esta categorización no fue basada en un procedimiento sistemátizado, por lo que la lista de especies invasoras que reportan puede tener un sesgo subjetivo. Por lo mencionado anteriormente, en este trabajo se pretende contribuir con información relevante sobre cuál es la probable distribución de algunas plantas exóticas que tienen carácter invasor en el estado; de esta manera, el objetivo de este estudio fue analizar la distribución potencial de ocho especies de plantas exóticas con presencia en Baja California, usando el programa de modelaje MaxEnt (versión 3.3.3e, Phillips et al., 2006). Estas especies fueron seleccionadas teniendo el conocimiento de que son plantas naturalizadas y forman parte importante del paisaje vegetal en las estaciones de invierno-primavera. Las razones mencionadas anteriormente, en complemento con los criterios establecidos para la selección de especies en el método, determinaron el interés de estudio de estas ocho especies. Posteriormente se llevarán a cabo otros estudios en donde se generarán mapas de distribución potencial de estas mismas especies en territorios vecinos (tanto en México como en el sur de California), los cuales presenten una similitud climática con Baja California. El hecho de hacer estos estudios de manera independiente podrá aportarnos un mejor conocimiento de las especies estudiadas para cada área en específico.
Materiales y métodos
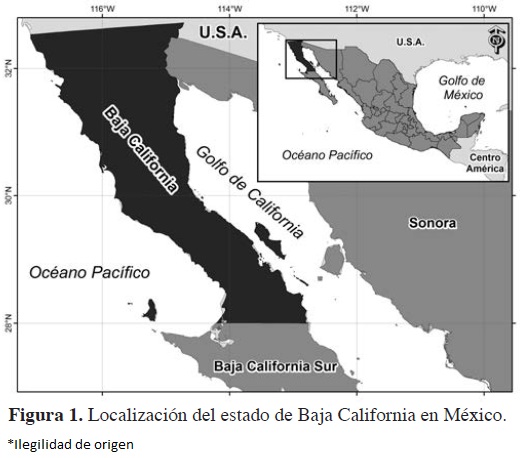
Área de estudio. El estado de Baja California está ubicado en el extremo noroeste de México, en la península del mismo nombre. Limita al norte con la frontera de Estados Unidos de América, al este con el Río Colorado y Golfo de California, al sur con el estado de Baja California Sur, y al oeste con el Océano Pacífico. Cuenta con una superficie de 70,113 km2 (Figura 1). Se ubica entre los meridianos 112° 48' y 117° 08' de longitud oeste y los paralelos 28° 00' y 32° 43' de latitud norte. El estado de Baja California está conformado por cinco municipios: Mexicali, que contiene a la capital del estado, Tijuana, Tecate, Ensenada y Playas de Rosarito (Gobierno del Estado de Baja California, 2011).
El clima de Baja California, por su diversidad altitudinal, latitudinal y topográfica, presenta diferencias notables y constantes. En términos generales, es más templada y fría la parte noroeste, mientras que la región este y central, seca y caliente. El clima del noroeste de la península es de tipo mediterráneo, al igual que la mayor parte del oeste de California, ya que presentan una similitud climática con otras partes del mundo, como la propia región mediterránea, parte central de Chile, sur de África y suroeste de Australia. Los climas de categoría BW (seco desértico), predominan en la mayor parte de la península, también se encuentra el clima BS (seco estepario). La categoría Cs es propia de las montañas de Baja California (Delgadillo, 1998).
Con respecto a la temperatura y precipitación del estado, la temperatura media anual es de 18 a 19 °C, y las temperaturas más altas, mayores de 30 °C, se presentan en los meses de mayo a septiembre, y las más bajas, alrededor de 5 °C, en el mes de enero. En la ciudad de Mexicali se han registrado temperaturas máximas extremas de hasta 52 °C entre los meses de julio y agosto. Las lluvias son muy escasas, alrededor de 200 mm de precipitación total anual. El municipio de Mexicali tiene uno de los registros de precipitación total anual más baja de todo el país, pues es menor a los 50 mm (INEGI, 2011). Los tipos de suelo presentes son: regosol, litosol, yermosol, xerosol, vertisol, feozem, fluvisol, solonetz, planosol y solonchak. De los diferentes tipos y asociaciones de suelos con que cuenta la entidad, destacan los regosoles, los litosoles y los yermosoles. Los más abundantes son los regosoles, que representan aproximadamente el 46% de la superficie del Estado (Gobierno del Estado de Baja California, 2011). De acuerdo a Delgadillo (1998), los tipos de vegetación para Baja California son: vegetación costera (marismas y dunas costeras), matorral costero suculento, chaparral, bosque de coníferas y matorral desértico.
Selección de especies. Se generó primero una lista de las plantas exóticas conocidas para el estado, se verificó que existieran registros de herbario en las base de datos del Consorcio Botánico de Baja California (http://bajaflora.org/), la cual cuenta con más de 78,000 especímenes. Estos se encuentran depositados en los herbarios del sur de California (SD, UCR, POM, RSA) y Baja California (BCMEX y HCIB). Esta lista posteriormente fue depurada, considerando los siguientes criterios: (1) que los taxa se encuentren en el listado de especies invasoras de México de la CONABIO, (2) que por lo menos cuenten con 13 registros de herbario en la base de datos del Consorcio Botánico y que los ejemplares depositados en el herbario BCMEX estuvieran correctamente identificados y (3) que estén catalogadas como invasoras en California de acuerdo a la base de datos de California Invasive Plant Inventory (Cal-IPC). Estos criterios fueron establecidos para trabajar principalmente con aquellas especies que están consideradas como invasoras para México, y que contaran con un número suficiente de registros de herbario para poder diseñar mejores modelos de distribución potencial. También se consideró que fueran invasoras de California, debido a que el área de estudio comparte características biogeográficas con el sur de California. Finalmente, ocho especies exóticas cumplieron con los criterios establecidos, para así proseguir a analizar la distribución potencial de las mismas (Cuadro 1).

Generación de modelos. Para generar los modelos se usaron los registros de herbario de las especies seleccionadas y 19 variables climáticas (Cuadro 2), las cuales se obtuvieron de la base de datos de WORLDCLIM (http://www.worldclim.org). Una vez obtenida la información necesaria, se utilizó el programa de predicción MaxEnt, el cual determina la relación entre las variables climáticas y los registros de las especies (Guisan y Thuiller, 2005). Se asume que el clima en los puntos de observación de una especie es representante de la gama ambiental del organismo; de esta forma, el clima se utiliza como el sistema de calibración para proponer un modelo de la probabilidad de presencia en base al clima (Jarvis et al., 2005). MaxEnt trata de encontrar la distribución de probabilidad de máxima entropía (cercana a la uniforme), sujeta a limitaciones impuestas por la información disponible en torno a la distribución observada de las especies y las condiciones ambientales del área de estudio. Al tener datos solo de presencia, MaxEnt crea puntos de pseudoausencias y divide la base de datos de registros de presencia en dos grupos: el primero se utiliza para construir el modelo mediante los datos de entrenamiento, mientras que el segundo es para evaluar el modelo al utilizar los datos de prueba (Phillips et al., 2006). Los modelos fueron generados con las características predeterminadas por MaxEnt; la única opción que se modificó fue el porcentaje aleatorio para los datos de prueba que validan los modelos.

Para Atriplex semibaccata y Centaurea melitensis la base de datos fue dividida en 75% para los datos que construyen el modelo y 25% para los datos de evaluación del modelo. En Brassica tournefortii se utilizó el 70% para los datos que construyen el modelo y 30% para los datos de evaluación del modelo, y en el caso de Bromus rubens y Schismus barbatus se utilizó 65% para los datos que construyen el modelo y 35% para los datos de evaluación del modelo. Para Cynodon dactylon, Salsola tragus y Tamarix ramosissima se utilizó el 60% para los datos que construyen el modelo y 40% para los datos de evaluación del modelo. Los porcentajes asignados fueron determinados a partir de ejecutar varias veces el programa MaxEnt para cada especie, y se eligieron los modelos que mejor se ajustaron a cada una de ellas. Los mapas obtenidos fueron exportados al programa ArcGIS 9.2 (ESRI, 2006), y se generaron los mapas finales del modelo potencial donde se muestran valores de baja (0) y alta (1) probabilidad de ocurrencia para cada especie.
Con MaxEnt se obtuvo la prueba Jackknife y la curva ROC/AUC (acrónimo del término en inglés Area Under the Curve) para cada modelo. La prueba Jackknife determina la contribución relativa de cada variable a los modelos generados. Los resultados son expresados con una medida conocida llamada "ganancia", esta información es de suma importancia para evidenciar los requerimientos ecológicos de cada especie. La curva ROC/AUC evalúa la habilidad predictiva de los modelos de distribución generados por medio de un gráfico, indicando el área bajo la curva (AUC), la cual representa la relación entre el porcentaje de presencia correctamente predicha (sensibilidad) y, uno menos el porcentaje de ausencias correctamente predichas (especificidad). El AUC mide la capacidad del modelo para clasificar correctamente a una especie como presente o ausente (Hanley y McNeil, 1982). Los valores de AUC pueden ser interpretados como la probabilidad de que, cuando se seleccione al azar un punto de presencia y uno de ausencia, el punto de presencia tendrá un valor mayor de idoneidad o probabilidad sobre el modelo. De acuerdo a Araújo y Guisan (2006), una guía general para la clasificación de la precisión del modelo tomando en cuenta el AUC es: 0.50-0.60 = insuficiente; 0.60-0.70 = pobre; 0.70-0.80 = promedio; 0.80-0.90 = bueno; 0.90-1 = excelente.
Resultados
Atriplex semibaccata. El área de mayor probabilidad de presencia de esta especie está prácticamente restringida a la parte costera del noroeste del estado hasta los 600 m de altitud, y desde Tijuana hasta el Ejido Eréndira, al sur de Ensenada. Disminuye su probabilidad de presencia a mayor altitud hacia el este (Figura 2A).
Brassica tournefortii. Su alta probabilidad de presencia se encuentra de manera discontinua en la parte costera del Pacífico. Resaltan dos áreas principales: la primera se encuentra en la costa del noroeste del estado hasta los 200 m de altitud y desde Tijuana, hacia el sur, hasta Ensenada, y la segunda en la parte costera central-oeste del estado hasta los 200 m de altitud y desde Camalú, hacia el sur, hasta San Quintín, en el municipio de Ensenada. También destacan dos áreas con una probabilidad media de presencia de esta especie, la primera va desde la costa central-oeste hasta los 400 m de altitud y desde El Rosario, hacia el sur, hasta Punta Prieta, apreciándose una clara reducción del área de probabilidad de presencia conforme disminuye la latitud. La segunda región con alta probabilidad de presencia se extiende desde las faldas de La Rumorosa, municipio de Tecate, hasta Laguna Salada en el municipio de Mexicali (Figura 2B).
Bromus rubens. Esta especie tiene dos áreas en las que su probabilidad de presencia es más alta; la primera se encuentra pegada a la costa noroeste del estado hasta los 400 m de altitud y desde Tijuana, hacia el sur, hasta el Ejido Eréndira, al sur de Ensenada, y la segunda se ubica en la porción central del estado a partir de los 1,600 m de altitud, en la Sierra de San Pedro Mártir. El área con una probabilidad media de presencia de esta especie se presenta como una continuación de la zona de mayor probabilidad en el noroeste hacia el centro norte del estado, cubriendo casi toda la Sierra de Juárez y hacia el sur hasta San Quintín (Figura 2C).
Centaurea melitensis. Su más alta probabilidad de presencia está prácticamente restringida a la parte costera del noroeste del estado hasta los 400 m de altitud y desde Tijuana hasta Camalú, al sur de Ensenada. El área de distribución de probabilidad media de la especie es una extensión de la anterior hasta los 1,000 m de altitud hacia las faldas de las sierras de Juárez y San Pedro Mártir, y hacia el sur hasta San Quintín (Figura 2D).
Cynodon dactylon. Esta especie presenta una distribución potencial de mayor probabilidad en el noroeste del estado, desde la costa hasta los 600 m de altitud y de Tijuana, hacia el sur, hasta el Ejido Eréndira, y en la región central del estado en Sierra de San Pedro Mártir por arriba de los 1,600 m de altitud. También presenta una distribución potencial de mediana probabilidad en el resto del estado de manera dispersa; sin embargo, resaltan las zonas del desierto de San Felipe al noreste, Sierra de Juárez en el centro norte, el área aledaña a la zona de mayor probabilidad en el noroeste, hacia el sur, hasta el Rosario en el municipio de Ensenada y, en el sureste del estado, cerca de Bahía de Los Ángeles (Figura 3A).
Salsola tragus. Tiene una alta probabilidad de presencia en casi todo el noroeste del estado, desde la costa hasta los 1,000 m de altitud en las laderas de Sierra de Juárez y hasta los 1,800 m de altitud en Sierra de San Pedro Mártir, y desde Tijuana, hacia el sur, hasta San Quintín. Las zonas de probabilidad media de presencia de esta especie cubren casi todo el centro norte (por debajo de los 1,600 m de altitud en Sierra de Juárez y por debajo de los 2,400 m de altitud en Sierra de San Pedro Mártir), la parte central y sur del estado (Figura 3B).
Schismus barbatus. En particular, esta especie muestra una gran cobertura de mayor probabilidad de presencia en todo el estado. Inicia en el noroeste, desde la costa hasta los 400 m de altitud, desde Tijuana, hacia el sur, hasta la ciudad de Ensenada, en esta localidad se amplía hasta llegar a los 1,000 m de altitud, aproximadamente en Sierra Juárez, y continua hacia el sur hasta Punta Colonet, ampliándose nuevamente de forma altitudinal hasta los 1,800 m en Sierra San Pedro Mártir, continuando hacia el sur hasta el Rosario, retomando los 1,000 m de altitud, y continua 100 km al sur del Rosario, disminuyendo paulatinamente de altitud conforme disminuye la latitud. Hay algunos manchones dispersos de probabilidad media de presencia de la especie, ubicados en la parte norte y sur del estado (Figura 3C).
Tamarix ramosissima. Las áreas de mayor idoneidad de presencia se encuentran principalmente en las costas y el Valle Mexicali; en el pacífico se extiende desde la costa hasta los 200 m de altitud y desde Tijuana hasta San Quintín y, habiendo una interrupción, continua hacia el sur en la costa de Santa Rosalita. En el Golfo de California se extiende también hasta los 200 m de altitud y desde Puertecitos hasta el paralelo 28° (límite sur del estado; Figura 3D).
Evaluación de la capacidad de predicción de los modelos y variables importantes. La eficiencia de predicción de los modelos muestra que las especies se encuentran en el intervalo del valor de AUC (curva ROC), de 0.744 a 0.946. Así, de acuerdo a los valores considerados por Araújo y Guisan (2006), Brassica tournefortii, Bromus rubens, Cynodon dactylon, Schismus barbatus y Tamarix ramosissima tienen una precisión del modelo clasificada como promedio; Centaurea melitensis y Salsola tragus, con una clasificación de precisión del modelo como buena; y sólo Atriplex semibaccata tiene una clasificación de precisión del modelo excelente (Cuadro 3).

La mayoría de los modelos aumentan su ganancia con variables relacionadas a la temperatura, cuando estas son usadas individualmente. En el caso de Atriplex semibaccata, Centaurea melitensis, Cynodon dactylon y Salsola tragus, la variable climática que más contribuye es la oscilación anual de la temperatura, mientras que para Bromus rubens y Schismus barbatus, la oscilación diurna de la temperatura y la estacionalidad de la temperatura son, respectivamente, las variables más influyentes. Las únicas especies que están más relacionadas a variables de precipitación cuando son usadas individualmente son Brassica tournefortii y Tamarix ramosissima. Contrariamente, las variables climáticas más importantes, que disminuyen la ganancia en los modelos cuando son omitidas, son las relacionadas a la precipitación. Para Atriplex semibaccata, Brassica tournefortii y Centaurea melitensis, resalta la variable de precipitación del cuatrimestre más cálido, y para Schismus barbatus y Tamarix ramosissima sobresale la precipitación del periodo más seco y la estacionalidad de la precipitación, respectivamente. En el resto de las especies, las variables relacionadas a la temperatura cuando son omitidas resultan tener más influencia (Cuadro 4).

Discusión
Las predicciones realizadas a partir del programa MaxEnt utilizando los registros de herbario y las 19 variables climáticas, muestran que la mayoría de las especies estudiadas tienen una mayor probabilidad de presencia en la zona mediterránea del estado, esto puede deberse a que es una zona en donde históricamente el intercambio florístico ha sido más intensivo y donde la actividad antropogénica (urbana y agrícola) es mayor. Lo anterior probablemente ha fomentado el que estas especies hayan encontrado un nicho ecológico que favoreció su naturalización y ahora presentan una mejor adaptación a las condiciones ambientales en esta zona. Pocas especies extienden su distribución potencial hacia las regiones desérticas, quizás porque la actividad humana es más limitada en esta zona y las condiciones climáticas les son menos favorecedoras.
Atriplex semibaccata. De acuerdo con Vibrans (2009), esta especie fue introducida de Australia como forrajera, pero se asilvestró muy exitosamente y ahora es una maleza seria en regiones con suelos salobres (p.ej., El Mezquital en Hidalgo, México), también es encontrada comúnmente en lugares con disturbio, cultivos abandonados y orillas de caminos. En el estado de California se ha convertido en invasora en matorrales y pastizales costeros, así como en la zona supralitoral de marismas (Cal-IPC, 2007). El área costera noroeste del estado parece proporcionarle a A. semibaccata un hábitat ideal para su persistencia y desplazamiento, principalmente por la presencia de suelos salinos.
Brassica tournefortii. De acuerdo con Vibrans (2009), esta planta es originaria del norte de África y es bien conocida por ser una invasora de desiertos, se ha señalado que en México se comporta como una típica arvense. Garcillán et al. (2009) reportaron que tiene preferencia por terrenos abandonados en la ciudad de Ensenada, Baja California. En Cal-IPC (2007), se menciona que B. tournefortii es abundante en ambas costas de Baja California; sin embargo, en el mapa de distribución potencial de esta especie su presencia no se observa en la zona costera del Golfo de California, por lo que es necesario contar con más colectas de herbario para poder tener un mejor diseño de la interpretación de la distribución potencial de esta planta, ya que sólo se encuentra en la costa del Pacífico.
Bromus rubens. Esta especie es originaria de la región mediterránea, se estableció por primera vez en California a mediados del siglo XIX y actualmente se encuentra en todo el Desierto Sonorense. De acuerdo con Winkler (1987), esta especie se ve favorecida en ambientes donde el clima sigue un patrón mediterráneo. La propuesta de Winkler concuerda con el mapa de distribución potencial, en donde se observa su presencia mayormente en la zona mediterránea del estado.
Centaurea melitensis. De acuerdo con Moroney y Rundel (2013), esta especie es nativa a la cuenca mediterránea y tiende a mostrar niveles más altos de dominancia y densidad en áreas invadidas de similar clima que el de su origen. Acorde a lo mencionado anteriormente, el área potencial de mayor probabilidad para C. melitensis en el estado, es la zona mediterránea y muy probablemente, esta especie pueda tener un nivel de dominancia bastante alto en esta región. El modelo de distribución potencial de C. melitensis llevado a cabo en este estudio contrasta con el modelo presentado para esta misma especie por Villaseñor et al. (2011), quienes reportan que prácticamente toda Baja California tiene una alta probabilidad de idoneidad para la presencia de C. melitensis. Sin embargo, es importante señalar que aunque se utilizó el mismo programa de modelaje MaxEnt, el método de trabajo fue diferente al de nuestro estudio. El modelo potencial generado para C. melitensis por Villaseñor et al. (2011) fue diseñado para todo el país, en donde no solo incluyeron registros de colectas para Baja California, ya que agregaron registros de otros dos estados (Sonora y Coahuila), añadieron una variable categórica y mediante un análisis de medias, sólo las variables más relevantes fueron consideradas en el modelo final de distribución potencial de C. melitensis. El uso de un diferente método en estos dos estudios conlleva a tener resultados distintos para esta especie.
Cynodon dactylon. Al igual que Bromus rubens, tiene una distribución potencial en la zona costera del pacífico y en el centro del estado a partir de los 1,600 m de altitud, en la Sierra de San Pedro Mártir, pero el alcance de su distribución también muestra su probabilidad de presencia en el desierto de San Felipe y el sur del estado. Este es un pasto altamente competitivo, muy importante en zonas tropicales y subtropicales, se ha extendido alrededor del mundo desde su origen en África tropical e Indo-Malasia. Probablemente fue introducida en Estados Unidos en el siglo XVII (Bryson y Wills, 1985). Es de los pastos exóticos más comunes en México, encontrándose como dominante sobre todo en sitios un poco más secos. Se comporta como ruderal y arvense en varios cultivos, y se distribuye ampliamente en áreas con disturbio (Vibrans, 2009). Por su origen tropical parece tener una mayor facilidad de establecerse en diferentes tipos de hábitats, quizás por esto, el mapa de distribución potencial muestra que se encuentra en gran parte del estado.
Salsola tragus. Esta especie es originaria de las estepas y cuencas saladas del sur de Rusia, fue introducida en Norteamérica probablemente en la segunda mitad del siglo XIX, y se distribuyó desde Canadá y Estados Unidos hasta estados del norte y centro de México. Se encuentra en zonas áridas, en terrenos salobres, parcelas de cultivo, agostaderos, orillas de carreteras, e invade, sobre todo, lugares en que ha sido removida de su lugar grandes cantidades de tierra (Vibrans, 2009). Salsola tragus es abundante en varias regiones semidesérticas de los Estados Unidos y el centro de Australia (Carnés et al., 2003). De acuerdo con Vibrans (2009), en México esta especie se ha cultivado como forrajera para zonas áridas, lo cual ha ayudado en su expansión. En Baja California se observa que el potencial de presencia es bastante amplio, abarcando la mayor parte del estado.
Schismus barbatus. Esta especie tiene una distribución potencial en el estado parecida a Salsola tragus, y de acuerdo a Randall et al. (1998), esta especie es de origen Mediterráneo. Actualmente está muy extendida en el suroeste de los Estados Unidos y el sur de Baja California (Chambers y Hawkins, 2002). Schismus barbatus es invasiva en el suroeste de Estados Unidos, en los desiertos de Mojave y Sonora, donde es común en áreas con disturbio, también provee comida para tortugas y roedores del desierto, aunque de baja aportación nutricional (GISD, 2010). Los resultados sugieren que tiene un gran potencial invasor en la mayor parte de Baja California, mostrando que toda la zona mediterránea y una gran parte de la zona norte y sur del estado son idóneas para la presencia de esta especie.
Tamarix ramosissima. Esta especie es un arbusto invasivo que ha dominado las zonas riparias de climas áridos (GISD, 2010). De acuerdo con Blackburn et al. (1982), es conocida por ser tolerante a la desecación de los cursos de agua y a la salinización de los suelos superficiales (Busch y Smith, 1993). También está considerada en la lista de las 100 especies exóticas invasoras más dañinas del mundo (GISD, 2010). Es la única que presenta sus áreas más óptimas en ambas costas del estado y en el Valle de Mexicali, lo que remarca sus requerimientos hídricos en su distribución. Lo anterior explica que las áreas más probables para su presencia en el estado sean los lechos de arroyos, áreas costeras salinizadas de ambas costas y zonas agrícolas, como por ejemplo los valles de Mexicali y San Quintín.
De las ocho especies tratadas en este trabajo, siete tienen un ciclo biológico anual de invierno-primavera, con una dependencia de la precipitación para su germinación. En la mayoría de las especies se observa que cuando las variables relacionadas con la precipitación son omitidas, disminuye el ajuste del modelo sobre las muestras de presencia; es decir, estas variables de precipitación contienen información más valiosa, que ayudan a diseñar un modelo más preciso de la distribución potencial de las especies.
En Brassica tournefortii, la variable de la precipitación del cuatrimestre más cálido es la más importante cuando es usada individualmente u omitida, similar con Tamarix ramosissima, aunque de ciclo de vida perenne, ya que la variable estacionalidad de la precipitación es repetida como la más importante cuando es usada de manera individual o cuando es omitida. En estas dos especies el requerimiento hídrico parece jugar un papel muy importante en su distribución.
Solo en el caso de Bromus rubens, la oscilación diurna de la temperatura influye de manera más significativa cuando es usada individualmente o cuando es omitida. De acuerdo a Salo (2004) este pasto anual parece ser capaz de germinar después de eventos de precipitación de 1 cm. Esto explica el por qué esté mayormente condicionada a una variable de temperatura, ya que aprovecha la mínima precipitación para germinar y, aunque es necesaria la precipitación para su desarrollo, no es tan exigente en cantidad, de esta manera, de acuerdo al modelo, la oscilación diurna de la temperatura tiene una influencia mayor sobre esta especie, de acuerdo al modelo.
Algunos autores recomiendan que para generar un mejor modelo de distribución potencial se necesita contar con la mejor información disponible, sugiriendo el uso mínimo de 50 localidades con coordenadas geográficas bien definidas (Peterson et al., 2000). En este trabajo se utilizaron entre 13 y 38 localidades (Cuadro 1); por lo tanto, ninguna especie tiene las condiciones deseables requeridas. Sin embargo, los modelos generados ofrecen hipótesis que pueden ayudar a orientar el trabajo futuro, el cual determinará con una mejor precisión parte del nicho potencial de las especies, representado en su distribución geográfica.
Sería interesante comparar estos modelos con otros programas como (Garp, BioClim, Domain, etc.), y examinar diferencias y/o similitudes entre ellos. Al considerar solo las variables climáticas en la elaboración del modelo, se está expuesto a tener errores en el diseño. Al contar con información como tipos de suelos, influencia antropogénica, interacciones bióticas, entre otras, se podría generar un modelo más completo y cercano a la realidad.
Agradecimientos
Agradecemos a Biól. Héctor Enrique Silvestre Ceballos Alcántara por su asistencia técnica y por su revisión en la traducción del resumen en inglés y a dos revisores anónimos por sus invaluables comentarios. También agradecemos al cuerpo académico Estudios Relativos a la Biodiversidad (Universidad Autónoma de Baja California) y a la Red Temática de Especies Exóticas de México, por el apoyo financiero parcial para la publicación de este trabajo.
Literatura citada
Anderson R.P., Lew D. y Peterson A.T. 2003. Evaluating predictive models of species' distributions: criteria for selecting optimal models. Ecological Modelling 162:211-232. [ Links ]
Araújo M.B. y Guisan A. 2006. Five (or so) challenges for species distribution modeling. Journal of Biogeography 33:1677-1688. [ Links ]
Blackburn W.H., Knight R.W. y Schuster J.L. 1982. Saltcedar influence on sedimentation in the Brazos River. Journal Soil Water Conservation 37:298-301. [ Links ]
Bryson C.T. y Wills G.D. 1985. Susceptibility of Bermudagrass (Cynodon dactylon) biotypes to several herbicides. Weed Science 33:848-852. [ Links ]
Busch D.E. y Smith S.D. 1993. Effects of fire on water and salinity relations of riparian woody taxa. Oecologia 94:186-194. [ Links ]
Cal-IPC. California Invasive Plant Council. 2007. <http://www.cal-ipc.org/> (consultado 14 de marzo 2011).
Carnés J., Fernández-Caldas E., Marina A., Alonso C., Lahoz C., Colás C. y Lezaun A. 2003. Immunochemical characterization of Russian thistle (Salsola kali) pollen extracts. Purification of the allergen Sal k 1. Allergy 58:1152-1156. [ Links ]
Chambers N. y Hawkins T.O. 2002. Invasive Plants of the Sonoran Desert: A Field Guide. Sonoran Institute, Environmental Education Exchange, National Fish and Wildlife Foundation. Tucson. [ Links ]
CONABIO. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. 2010. Sistema de información sobre especies invasoras en México. <http://www.conabio.gob.mx/invasoras> (consultado 14 de enero 2011). [ Links ]
D'Antonio C.M., Dudley T.L. y Mack M. 1999. Disturbance and biological invasions: direct effects and feedbacks. En: Walker L.R. Ed. Ecosystems of Disturbed Ground, pp. 413-452, Elsevier Science, Amsterdam. [ Links ]
Delgadillo J. 1998. Florística y Ecología del norte de Baja California. 2a ed. Universidad Autónoma de Baja California, Mexicali. [ Links ]
ESRI. Environmental Systems Research Institute. 2006. ArcView Geostatistical Analyst. Redlands. [ Links ]
Fielding A.H. y Bell J.F. 1997. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environmental Conservation 24:38-49. [ Links ]
Garcillán P.P., León de la Luz J.L., Rebman J.P. y Delgadillo J. 2013. Plantas no nativas naturalizadas de la Península de Baja California, México. Botanical Sciences 91:461-475. [ Links ]
Garcillán P.P., Rebman J.P. y Casillas F. 2009. Analysis on the non-native flora of Ensenada, a fast growing city in Northwestern Baja California. Urban Ecosystems 12:449-463. [ Links ]
GISD. 2010. Global Invasive Species Database. Invasive Species Specialist Group (ISSG). Species Survival Commission (SSC) de la IUCN (International Union for Conservation of Nature). <http://www.issg.org/database/welcome> (consultado 14 de enero 2011).
Gobierno del Estado de Baja California. 2011. http://www.bajacalifornia.gob.mx/ (consultado 13 de enero 2011).
Guisan A. y Zimmermann N.E. 2000. Predictive habitat distribution models in ecology. Ecological Modelling 135:147-186. [ Links ]
Guisan A. y Thuiller W. 2005. Predicting species distribution: offering more than simple habitat models. Ecology Letters 8:993-1009. [ Links ]
Hanley J.A. y McNeil B.J. 1982. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 143:29-36. [ Links ]
INEGI. Instituto Nacional de Estadística y Geografía. 2011. <http://www.inegi.org.mx/monografias/informacion/bc/territorio/clima.aspx?tema=me> (consultado 14 de enero 2010).
Jarvis A., Williams K., Williams D., Guarino L., Caballero P.J. y Mottram G. 2005. Use of GIS for optimizing a collecting mission for a rare wild pepper (Capsicum flexuosum Sendtn.) in Paraguay. Genetic Resources and Crop Evolution 52:671-682. [ Links ]
Lindenmayer D.B., Nix H.A., McMahon J.P., Hutchinson M.F. y Tanton M.T. 1991. The conservation of Leadbeater's possum, Gymnobelideus leadbeateri (McCoy): a case study of the use of bioclimatic modelling. Journal of Biogeography 18:371-383. [ Links ]
Mack M.C. y D'Antonio C.M. 1998. Impacts of biological invasions on disturbance regimes. Trends in Ecology and Evolution 13:195-198. [ Links ]
Mack R.N. 1991. The commercial seed trade: an early disperser of weeds in the United States. Economic Botany 45:257-273. [ Links ]
Moroney J.R. y Rundel P.W. 2013. Abundance and dispersion of the invasive Mediterranean annual, Centaurea melitensis in its native and non-native ranges. Biological Invasions 15:495-507. [ Links ]
Peterson A.T., Egbert S.L., Sánchez-Cordero V. y Price K.P. 2000. Geographic analysis of conservation priority: endemic birds and mammals in Veracruz, Mexico. Biological Conservation 93:85-94. [ Links ]
Phillips S.J., Anderson R.P. y Schapire R.E. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190:231-259. [ Links ]
Randall M.J., Rejmánek M. y Hunter J.C.1998. Characteristics of the exotic flora of California. Fremontia 26:3-12. [ Links ]
Richardson D.M., Pyšek P., Rejmánek M., Barbour M.G., Panetta F.D. y West C.J. 2000. Naturalization and invasion of alien plants: concepts and definitions. Diversity and Distributions 6:93-107. [ Links ]
Rzedowski J. y Calderón R.G. 1990. Nota sobre el elemento africano en la flora adventicia de México. Acta Botanica Mexicana 12:21-24. [ Links ]
Salo L.F. 2004. Population dynamics of red brome (Bromus madritensis subsp. rubens): times for concern, opportunities for management. Journal of Arid Environments 57:291-296. [ Links ]
Segurado P. y Araújo M.B. 2004. An evaluation of methods for modelling species distributions. Journal of Biogeography 31:1555-1568. [ Links ]
Soto-Hopkins J. 1987. Identificación taxonómica de malezas en cultivos de tomate (Lycopersicum esculentum L.) en la región de San Quintín, Baja California. Tesis licenciatura en Biología, Escuela Superior de Ciencias, Universidad Autónoma de Baja California, Ensenada. 53 pp. [ Links ]
Vibrans H. 2009. Malezas de México. <http://www.conabio.gob.mx/malezasdemexico/2inicio/home-malezas-mexico.htm> (consultado 14 de marzo 2011). [ Links ]
Villaseñor J.L. y Espinosa-García F.J. 2004. The alien flowering plants of Mexico. Diversity and Distributions 10:113-123. [ Links ]
Villaseñor J.L. y Magaña P. 2006. Plantas introducidas en México. Ciencias 82:38-40. [ Links ]
Villaseñor J.L., Ortiz E., Hinojosa-Espinosa O. y Segura-Hernández G. 2011. Especies de la Familia Asteraceae Exóticas a la Flora de México. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación, Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria, Consejo Nacional Consultivo Fitosanitario, Universidad Nacional Autónoma de México, Instituto de Biología, Asociación Mexicana de Ciencia de la Maleza, México, D.F. [ Links ]
Vitousek P.M. 1990. Biological invasions and ecosystem processes: Toward an integration of population biology and ecosystem studies. Oikos 57:7-13. [ Links ]
Weber E.F. 1997. The alien flora of Europe: a taxonomic and biogeographic review. Journal of Vegetation Science 8:565-572. [ Links ]
Williamson M. 1996. Biological Invasions. Chapman y Hall, Londres. [ Links ]
Winkler G. 1987. Bromus rubens. En: Fischer W.C. Compiler. The Fire Effects Information System [Data base]. U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Intermountain Fire Sciences Laboratory, Missoula. http://www.fs.fed.us/database/feis/plants/graminoid/brospp/all.html (consultado 7 de enero de 2009). [ Links ]
Zaniewski A.E., Lehmann A. y Overton J.M. 2002. Predicting species spatial distributions using presence-only data: a case study of native New Zealand ferns. Ecological Modelling 157:261-280. [ Links ]

