










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBotanical Sciences
versión On-line ISSN 2007-4476versión impresa ISSN 2007-4298
Bot. sci vol.91 no.2 México jun. 2013
Ecología
Reforestación con Cupressus lusitanica y su influencia en la diversidad del bosque de pino-encino en Los Altos de Chiapas, México
Reforestation with Cupressus lusitanica and its influence on the diversity of pine-oak forest in the Highlands of Chiapas, Mexico
Laura Fernández-Pérez1,2, Neptalí Ramírez-Marcial1,3 y Mario González-Espinosa1
1 Departamento de Conservación de la Biodiversidad, El Colegio de la Frontera Sur (ECOSUR), San Cristóbal de Las Casas, Chiapas, México.
2 Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental Chetumal, Chetumal, Quintana Roo, México.
3 Autor para la correspondencia: nramirezm@ecosur.mx
Recibido: 25 de junio de 2012
Aceptado: 24 de octubre de 2012
Resumen
El uso extendido de Cupressus lusitanica para reforestar áreas degradadas puede reducir la riqueza y diversidad de bosques húmedos de montaña. Se comparó la composición y estructura de la vegetación leñosa de un área reforestada con C. lusitanica con la de fragmentos de bosque de Pinus-Quercus y bosques secundarios de pino-encino en Los Altos de Chiapas. Los tres tipos de bosque presentaron diferencias significativas (P ≤ 0.01) en cobertura, riqueza y diversidad de especies leñosas. La mayor área basal de árboles grandes se registró en el bosque de Pinus-Quercus (38.7 m2ha-1) seguida del bosque secundario (16.5 m2ha-1) y la más baja en el sitio reforestado con Cupressus (12.7 m2ha-1). Aunque la menor riqueza (P ≤ 0.05) de especies arbóreas se registró en esta última condición (16 especies), comparada con el bosque de Pinus-Quercus (18) y el bosque secundario (30 especies), la diversidad de árboles (> 10 cm de diámetro a la altura del pecho) resultó mayor en el sitio reforestado con Cupressus (P ≤ 0.05). Las principales variables edáficas que explicaron las diferencias entre las parcelas mediante un análisis de escalamiento no métrico multidimensional fueron el contenido de fósforo y arcilla (P < 0.05). Se concluye que la reforestación después de 28 años ha sido efectiva para recuperar la cobertura del sitio; además de permitir la presencia de varias de las especies encontradas en los bosques aledaños no reforestados.
Palabras clave: bosque de Pinus-Quercus, Ciprés mexicano, plántulas, reforestación, sucesión.
Abstract
The widespread use of Cupressus lusitanica to recover degraded forest areas may reduce species richness and diversity of montane forests. Woody composition and structure in a reforested area with C. lusitanica were compared with fragments of Pinus-Quercus forest and secondary pine-oak forests in the Highlands of Chiapas. The three forest types differ significantly (P ≤ 0.01) in coverage, richness and diversity of woody species. The highest basal area of large trees occurred in the Pinus-Quercus forest (38.7 m2ha-1), followed by secondary forest (16.5 m2ha-1) and the Cupressus reforested forest (12.7 m2ha-1). Although the lowest tree richness (P ≤ 0.05) was recorded in the latter forest (16 species), compared with Pinus-Quercus forest (18) and secondary forest (30 species), diversity of trees (> 10 cm diameter at breast height) was greater under Cupressus reforested site (P ≤ 0.05). According to the analysis of non-metric multidimensional scaling major soil variables that explained the differences between the plots were the phosphorus and clay content (P < 0.05). It is concluded that reforestation after 28 years has been effective to retrieve site cover and allowed the presence of several species in the surrounding forests not reforested, although it has not yet reached the values of basal area.
Key words: Mexican Cypress, plant succession, reforestation, seedlings.
Los bosques contribuyen a la provisión de bienes y servicios ambientales, tales como la conservación de diversidad biológica, captura de carbono y a la regulación del ciclo hidrológico y el clima (Rey Benayas et al., 2009; Seppelt et al., 2011). A pesar de esta importancia, los bosques del mundo se encuentran altamente fragmentados o degradados en su composición, estructura y funcionamiento (Price et al., 2011).
En particular, los bosques de montaña albergan una alta diversidad biológica y cumplen funciones ecológicas como la regulación climática, conservación de biodiversidad y provisión de agua, además de servir como elemento cultural integrador para numerosas poblaciones humanas dispuestas en sus áreas circundantes (Konijnendijk, 2008). Sin embargo, la misma presencia humana ha significado una amenaza continua a la permanencia del bosque (Webster, 2002; Konijnendijk, 2008). El crecimiento demográfico y la expansión de la agricultura intensiva han obligado a los pequeños agricultores a moverse hacia zonas periurbanas, normalmente con fuertes pendientes, y por lo tanto, se han deforestado o empobrecido las zonas forestales. Aunque el cambio más perceptible de la deforestación por cambio de uso del suelo es la pérdida de la cubierta vegetal, también se aprecian procesos de degradación de la calidad de los fragmentos remanentes por pérdida de especies del interior (especies tolerantes a la sombra; Cayuela et al., 2006).
Actualmente, persiste el debate respecto a si las actividades de reforestación -con un número relativamente bajo de especies- son efectivas para recuperar las diversos atributos y valores del bosque. Por ejemplo, hay evidencia que las plantaciones de árboles en sitios altamente degradados pueden acelerar la sucesión vegetal, en comparación con el tiempo requerido por los procesos de regeneración natural (Lugo et al., 1993; Guariguata et al., 1995). En tales escenarios de degradación, la opción de elegir especies de rápido crecimiento y tolerantes a condiciones extremas de degradación ha sido una práctica común en numerosos bosques neotropicales. El supuesto es que estas especies modifican algunas condiciones desfavorables del suelo y microclima, para crear nuevas condiciones que promueven el arribo paulatino de las especies locales, lo que conlleva a una recuperación de la estructura y función del bosque (Parrotta et al., 1997).
La reforestación con especies forestales maderables, normalmente persigue objetivos precisos para recuperar la cobertura vegetal en poco tiempo y a la postre, obtener un beneficio económico a partir del aprovechamiento de la madera (Guariguata et al., 1995). Sin embargo, ante la ausencia de un manejo silvícola apropiado durante el desarrollo de la plantación, se crean oportunidades para el enriquecimiento natural de especies a partir de factores tales, como la presencia de áreas vecinas de vegetación original que puedan funcionar como fuente potencial de nuevos propágulos, de las condiciones de degradación del sitio y del tiempo transcurrido desde el inicio de la plantación.
Actualmente, las acciones de restauración o rehabilitación forestal en México se realizan exclusivamente para lograr la repoblación de los sitios que de manera natural o por influencia humana han perdido su cobertura vegetal. Pocas veces se privilegia el uso de especies arbóreas nativas propias de cada región y, cuando ocurre, está restringido a un número mínimo de especies. Esta actividad ha sido fomentada mediante diversos programas de apoyo a la reforestación, pero con nulo o escaso seguimiento de las condiciones de supervivencia y crecimiento de las plantas (Cervantes et al., 2008).
En varias localidades del estado de Chiapas y en otros estados del país, se han realizado planes de reforestación utilizando especies de rápido crecimiento como el ciprés mexicano (Cupressus lusitanica Mill. var. lusitanica, Farjon, 1993). Debido a que esta especie es maderable (elaboración de polines y tablas), su uso promovido por programas locales y regionales de reforestación no consideran los impactos ecológicos favorables o nocivos que puedan tener sobre la estructura y el funcionamiento del bosque (Cervantes et al., 2008). Bajo ciertas circunstancias, una reforestación monoespecífica puede considerarse análoga a una plantación forestal con fines productivos. Varios estudios han indicado las ventajas de las plantaciones para la restauración de algunos atributos del suelo (p. ej., Parrotta, 1992, Lugo, 1997), y otros que indican que las plantaciones tienden a acelerar la degradación del suelo y otros procesos biológicos (Richter et al., 1999).
Una de las formas de evaluar el éxito de las acciones de reforestación y restauración forestal es analizar la estructura y diversidad del sitio intervenido con relación a otro sitio de referencia (Ruiz-Jaen y Aide, 2005). El objetivo de este estudio fue evaluar las condiciones de estructura y composición de especies leñosas en un sitio de reforestación con Cupressus lusitanica var. lusitanica establecida desde 1984 en La Milpoleta, un área periférica a la ciudad de San Cristóbal de Las Casas, Chiapas en comparación con las condiciones presentes en un bosque de Pinus-Quercus aledaño y otro sitio de vegetación secundaria. Se propone que el tiempo desde la reforestación y, ante la ausencia de un manejo silvícola del área, se espera que la riqueza y diversidad de plantas leñosas serán inferiores con respecto a los bosques aledaños que han seguido una dinámica de regeneración y crecimiento natural.
Materiales y métodos
Área de estudio. Los tres sitios de estudio se localizan en la Altiplanicie Central de Chiapas dentro de la cuenca de San Cristóbal de Las Casas, en un intervalo altitudinal de 2,080 a 2,430 m, en los límites de los municipios de San Juan Chamula y San Cristóbal de Las Casas (Cuadro 1). El área se caracteriza por presentar un paisaje heterogéneo que incluye fragmentos de bosques, áreas de producción de cultivos anuales (maíz, frijol, calabaza, flores de ornato, hortalizas) y perennes (cultivos de pera, manzana, durazno, ciruela), y ganadería de ovinos y bovinos. Los niveles de erosión hídrica calculados en la cuenca de San Cristóbal de Las Casas desde 1975 al 2000, debido a la pérdida de cubierta forestal y al aumento de zonas agrícolas y urbanas en ese periodo, indican volúmenes de escurrimiento medio anual del orden de 77 a 82 millones m3 (Figueroa-Jáuregui et al., 2011).
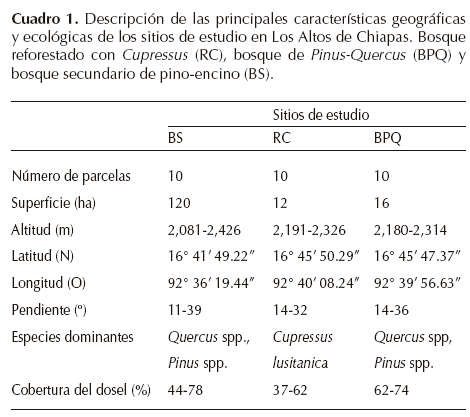
El clima es templado subhúmedo, con temperatura media anual de 14.7 °C y promedio de precipitación total anual de 1,104 mm, en la cual, más de 80% de la precipitación se presenta de mayo a octubre. Las heladas nocturnas son frecuentes de diciembre a marzo, principalmente en las partes planas y abiertas. Los suelos son arcillosos, moderadamente profundos, de color café oscuro y derivados de rocas calizas. La vegetación incluye diversas etapas sucesionales derivadas de bosque de pino-encino (González-Espinosa et al., 2006).
Específicamente, se eligió como área de estudio un rodal de 12 ha en La Milpoleta (en lo sucesivo, bosque reforestado con Cupressus lusitanica, RC, Figura 1A), debido a que en este sitio ocurrió un incendio severo en mayo de 1983, año que coincidió con una temporada seca muy prolongada según los registros de la estación meteorológica "La Cabaña", que se ubica en el valle de San Cristóbal. En julio de 1984 se realizó la reforestación con el ciprés mexicano (Cupressus lusitanica var. lusitanica), con plantas de dos años producidas en un vivero local de la Secretaría de Agricultura y Recursos Hidráulicos (SARH), en San Cristóbal de Las Casas. La plantación se realizó con cepellón sobre líneas en curvas de nivel a una densidad entre 625-1,000 plantas por hectárea. El segundo sitio de estudio (bosque de Pinus-Quercus, BPQ) es un área de 16 ha aledaña al RC, pero que no fue afectada por el incendio y aún conserva la fisonomía de los bosques de Pinus-Quercus típicos de la región (Figura 1B). El tercer sitio (bosque secundario, BS) se ubica en la localidad de El Aguaje (120 ha en el extremo sureste de la microcuenca de San Cristóbal de Las Casas) que ha estado bajo un régimen de extracción continua de leña y madera, además de que fue afectado por un incendio superficial de baja intensidad en el año de 1998 (Figura 1C, Cuadro 1). La razón de incorporar este sitio fue contrastar un sitio forestal perturbado, pero de edad más reciente, y donde nunca se ha realizado la reforestación.
Condiciones edáficas. En cada tipo de vegetación se tomaron 10 muestras de suelo superficial a una profundidad de 0-20 cm (3 kg de suelo por muestra aproximadamente). Las muestras se pusieron a secar al aire, se molieron y tamizaron, para posteriormente realizarles por triplicado los siguientes análisis físicos y químicos en el laboratorio de suelos de ECOSUR: textura (método de Bouyoucos), densidad aparente (método gravimétrico), pH (relación 1:2 con H2O), contenido de fósforo extraíble (método de olsen), nitrógeno total (método semi-microKjeldhal), materia orgánica (método de Walkley y Black) y capacidad de intercambio catiónico (método de acetato de amonio 1N pH 7).
Composición y estructura. En cada tipo de bosque (BS, RC y BPQ) se establecieron y marcaron diez parcelas circulares de 0.1 ha, siguiendo la metodología de Ramírez-Marcial et al. (2001). En cada parcela se contó y midió el diámetro a la altura del pecho (DAP, a 130 cm de altura) de todos los individuos de especies leñosas con un DAP mayor a 5 cm. El muestreo se estratificó de acuerdo con el tamaño diamétrico de los individuos. Los individuos "grandes" (DAP > 20 cm) se midieron en un área circular de 1,000 m2 (radio, r = 17.84 m), los individuos "medianos" (10.1-20 cm de DAP) dentro de una subparcela de 500 m2 (r = 12.61 m) y los individuos "chicos" (5-10 cm de DAP) dentro de una subparcela de 100 m2 (r = 5.64 m). De cada especie se obtuvo una muestra botánica con la que se elaboró un catálogo de referencia para su identificación. Las especies se determinaron en el campo con la ayuda de un técnico experto en la flora local, y cuando no se reconocieron se colectaron algunos individuos para su posterior determinación en el herbario de El Colegio de La Frontera Sur en San Cristóbal de Las Casas, donde se encuentran depositados los especímenes. Para cada categoría de tamaño, se calculó la densidad, frecuencia y dominancia (área basal) de todas las especies (riqueza) y con ello se elaboró el Índice de Valor de Importancia Relativa (VIR), considerando los valores relativos de la densidad y dominancia (área basal) (Mueller-Dombois y Ellenberg, 1974). Con la densidad de individuos se calculó el índice de diversidad de Simpson, expresado como -Ln (índice de concentración de Simpson; Rosenzweig, 1995). La cobertura del dosel se obtuvo con un densiómetro convexo (Lemmon, 1956), estimada como el porcentaje de cobertura aérea reflejada por los árboles del dosel (Cook et al., 1995). Se tomaron cinco lecturas en cada parcela de 1,000 m2 y se obtuvo un valor promedio por parcela.
Regeneración natural y tocones. Se trazaron aleatoriamente tres transectos de 30 x 1 m (un total de 90 m2 de muestreo por cada tipo de bosque), para registrar la presencia de regeneración natural expresada como el número de individuos con una altura inferior a 1 m. Para registrar la ocurrencia de extracción de madera (actual y pasada), se cuantificó dentro de las parcelas circulares de 1,000 m2 la presencia de tocones y rebrotes a los que se les midió el diámetro a la base del tronco principal para determinar el área basal y calcular el VIR. En la medida de lo posible, los tocones fueron identificados hasta especie y cuando no, al menos hasta género. La abundancia y riqueza de plántulas y de tocones se analizaron con un análisis de varianza (ANOVA), entre las tres condiciones de bosque.
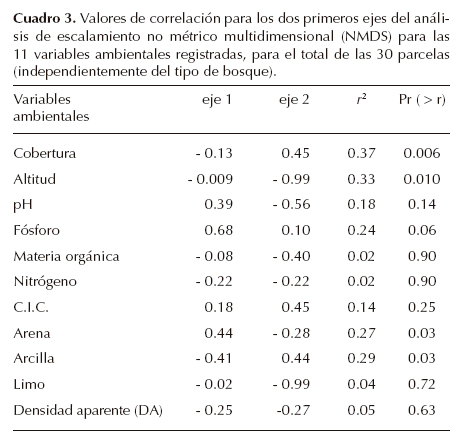
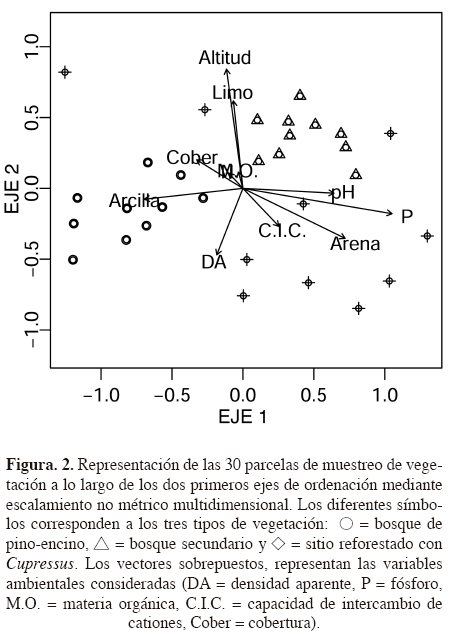
Análisis. Debido a la heterogeneidad dentro y entre parcelas detectada durante el muestreo, se realizó un primer análisis de ordenación indirecta de las parcelas con las variables ambientales; para ello se construyó una primera matriz de datos con los inventarios de las 30 parcelas que incluyó la densidad de todos los individuos > 5 cm de DAP; la segunda matriz consistió de 11 variables (nueve variables edáficas más la altitud y cobertura del dosel). El método de ordenación fue el de escalamiento no métrico multidimensional, NMDS (Non-metric Multidimensional Scaling) que utiliza la matriz de disimilitud entre parcelas; este método se considera efectivo para ordenar datos ecológicos, al no requerir el supuesto de relación lineal entre las variables, y por ello se utilizó un procedimiento automatizado para encontrar el valor de estrés más pequeño a partir de un procedimiento de 1,000 reiteraciones, incluido en la función metaMDS (Oksanen, 2011) operada desde el programa R, versión 2.12 (R Development Core Team, 2012). El método de ajuste de las variables ambientales sobre la ordenación de las parcelas consistió en sobreponer los vectores de las variables ambientales sobre los ejes de ordenación con la función envfit. Para ello, se usó la segunda matriz con las 11 variables ambientales; el análisis proporcionó un valor de correlación de cada variable y significancia de cada una a lo largo de los dos primeros ejes de la ordenación.
En un segundo nivel de análisis, los valores de riqueza de especies, abundancia y diversidad se analizaron para cada categoría de tamaño (plántulas, árboles chicos, medianos y grandes) dentro de cada tipo de vegetación mediante un análisis de varianza (AVOVA), o la prueba de Wilcoxon cuando no se satisficieron los supuestos de normalidad y/o homogeneidad de varianzas. Cuando alguna de estas variables resultó significativa, se utilizó la prueba de comparación de medias entre condiciones con la prueba de Tukey y la prueba U de Mann-Whitney. Todos los análisis se realizaron con el programa R, versión 2.12. (R Development Core Team, 2012).
Resultados
Caracterización física y edáfica de los sitios. Del total de 11 variables ambientales analizadas entre los tres tipos de vegetación, cinco de ellas resultaron significativamente diferentes (Cuadro 2). La cobertura de la vegetación en el sitio reforestado con Cupressus fue significativamente inferior con respecto a los otros dos sitios. Se observó un incremento significativo (P ≤ 0.03) en el valor de pH, fósforo y contenido de arena en el sitio reforestado (RC) con respecto a los otros dos tipos de bosque, mientras que el de arcilla fue significativamente más bajo (Cuadro 2).
La ordenación de todas las parcelas (N = 30) mediante el análisis NMDS, mostró una relación significativa con cinco variables ambientales (cobertura, altitud, contenido de fósforo, arcilla y arena), que explicaron entre 27-33% de la variación entre las parcelas (Cuadro 3). El coeficiente de bondad de ajuste (estrés) para los tres primeros ejes de la ordenación fue de 11.31, y permitió visualizar la relativa correspondencia de los dos primeros ejes con la variación ambiental entre las parcelas para los tres tipos de vegetación. Lo más notable, es que las parcelas del sitio reforestado con Cupressus lusitanica mostraron una alta variación respecto a la posición de sus parcelas a lo largo de los dos ejes de la ordenación (Figura 2).
Composición y estructura forestal. La mayor cobertura del dosel se registró en el BPQ (70.6 %), seguida de BS (64.2 %) y RC (55%; Cuadro 1). Esta condición de dosel más abierto, registrada en el sitio RC, resultó significativamente diferente del BPQ (P ≤ 0.001), pero no entre BS y RC, ni entre BS y BPQ. La riqueza total de especies leñosas registrada en las 30 parcelas fue de 35 especies, totalizando 2,266 individuos censados. de este total, 30 especies estuvieron presentes en el BS (85.7%), 18 especies en el BPQ (51.4%) y 16 especies en el sitio reforestado con Cupressus (45.7%; Cuadro 4).
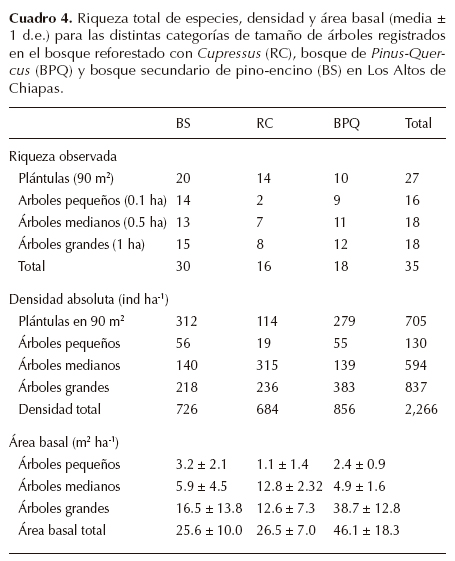
El promedio de la riqueza de especies por parcela fue similar entre BS (4.1 ± 2.3 d.e.) y BPQ (4.4 ± 1.3 d.e.), ambas condiciones resultaron significativamente diferentes con respecto al sitio RC (2.7 ± 1.3 d.e.; X2 =13.24, g.l. = 2, P ≤ 0.001). La riqueza observada fue significativamente diferente en tres de las cuatro categorías de tamaño consideradas (excepto árboles medianos), con una mayor riqueza de especies en el BS con respecto al BPQ y RC (Figura 3).
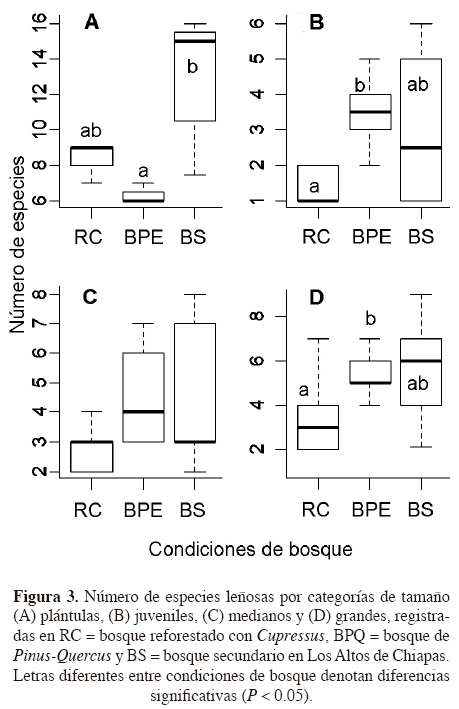
La mayor densidad de árboles pequeños se registró en el BS, la de los árboles medianos en el sitio RC y la de árboles grandes en el BPQ (Cuadro 4). La prueba U-Mann-Whitney evidenció diferencias significativas en la densidad absoluta de los árboles grandes entre BS y RC (P ≤ 0.05) y entre BS y BPQ (P ≤ 0.05). La mayor acumulación de área basal (P ≤ 0.01) se registró en el BPQ (46.1 m2ha-1), seguido de valores significativamente más bajos en el BS (25.6 m2ha-1) y el sitio RC (26.5 m2ha-1, Cuadro 4).
Al considerar los valores de importancia (VIR) por tipo de bosque y categorías de tamaño dentro del sitio RC, Cu- pressus lusitanica representó el 68.8, 75.4 y 53.9%, para las categorías de árboles chicos, medianos y grandes, respectivamente. La segunda especie más importante fue Pinus pseudostrobus (31.2, 13.9 y 19.8%). En el BS las especies más importantes en la categoría de árboles pequeños fueron Cleyera theoides (15.5%), Quercus crispipilis (12.4%) y Q. segoviensis (12.4%); en la de tamaño mediano fueron Q. rugosa (28.6%), C. theoides (21.1%) y Q. segoviensis (12.6%). En la categoría de árboles grandes fueron Q. rugosa (32.9%), P. pseudostrobus (21.9%) y Q. crispipilis (18.2%). En el BPQ las siguientes especies representaron el 66.9% del VIR en la categoría de árboles pequeños: Crataegus mexicana (24.6%), Q. segoviensis (21.6%) y Garrya laurifolia (20.7%). En árboles medianos, Q. segoviensis (39.6%), seguido de Q. crispipilis (23.7%) y C. mexicana (9.7%). Para los árboles grandes, P. pseudostrobus fue la más importante (37.4%), seguida de Q. segoviensis (30.6%) y Q. crispipilis (19.6%).
Diversidad de especies. El índice de concentración de Simpson (-Ln) resultó significativamente diferente entre los tipos de vegetación y entre las categorías de tamaño (Figura 4). Para todos los casos (excepto árboles chicos), la diversidad más alta se registró en el RC, aunque únicamente se detectaron diferencias significativas entre los tres tipos de bosque para los árboles medianos (F2,26 = 4.64, P ≤ 0.05) y grandes (F2,26 = 3.50, P ≤ 0.05). En ambos casos, las diferencias correspondieron al significativamente mayor valor de diversidad del sitio RC respecto a las otras dos condiciones de bosque.
Regeneración y tocones. La riqueza de especies registradas como plántulas fue de 27 especies, 20 de las cuales se distribuyeron en el BS, 14 en sitio RC y diez en el BPQ. De igual manera, la densidad de plántulas varió entre los tres tipos de bosque (P ≤ 0.001). Las especies más abundantes en RC fueron Prunus serotina y Cornus excelsa; en el BS Quercus crispipilis y P. serotina, y en el BPQ, Q. crispipilis y P. serotina. El mayor número de tocones (110 tocones) se registró en el sitio RC, seguido de BPQ (84) y BS (55), cuya diferencia fue significativa (X2 = 4.116, g.l. = 2, P < 0.05). El área basal del total de tocones fue similar (X2 =1.039, g.l. = 2, P > 0.5) entre los tipos de bosque: BS (7.4 ± 0.19 m2ha-1), RC (6.6 ± 0.08 m2ha-1) y BPQ (4.9 ± 0.09 m2ha-1), y correspondió principalmente a individuos de Quercus y Pinus (61-79% del VIR total de los tocones).
Discusión
El análisis de ordenación mostró que no todas las características edáficas del bosque reforestado con Cupressus (RC) difieren respecto a los otros dos tipos de vegetación. Aunque se aprecian diferencias en las concentraciones de fósforo y en la mayor predominancia de arena en el suelo bajo la cobertura de Cupressus, la ordenación indicó un mayor peso a la cobertura y a la altitud para diferenciar la estructura actual de los tres tipos de bosque. En estudios previos ya se ha documentado que las plantaciones de C. lusitanica tienen pocos efectos sobre las propiedades del suelo. Por ejemplo, Cavelier y Tobler (1998) al comparar plantaciones de Pinus y de Cupressus (21 años de establecidas), con un bosque mesófilo de montaña en Los Andes colombianos, encontraron diferencias sólo en el contenido de carbono y la relación C/N. En otro estudio realizado en México, Bautista-Cruz y del Castillo (2005) no encontraron una relación directa entre la pérdida de suelo y la deforestación de un bosque mesófilo en Oaxaca hasta después de 45 años. una posible explicación podrá atribuirse a que el tiempo desde el establecimiento de la reforestación es aún reciente como para modificar las propiedades físicas y químicas del suelo con respecto a las características de los bosques de referencia (en este caso el BPQ).
A pesar de que, numéricamente, el pH del suelo en el sitio reforestado con Cupressus fue mucho más básico (7.1) con respecto al BPQ (5.8) y BS (6.1), estas diferencias no fueron significativas y contradice los hallazgos de otros autores que señalan que el pH tiende a reducirse en plantaciones de Cupressus (Lemenih et al., 2004). Probablemente la mayor dominancia de especies latifoliadas en BPQ y BS proporcionan un alto contenido de materia orgánica que tiende a mantener los suelos neutros o ácidos (Ramírez-Marcial et al., 2001; Galindo-Jaimes et al., 2002), aunque tampoco se detectaron diferencias significativas en el contenido de materia orgánica entre los tres tipos de bosque. Por otro lado, se ha documentado que las plantaciones de Cupressus en El Salvador (Price, 1982) presentan una mayor compactación del suelo y reducen significativamente la cantidad de agua disponible para la recarga de agua en el suelo y de escorrentía en comparación con plantaciones de Pinus pseudostrobus y de un bosque de pino-encino. Debido a que en el presente estudio no se evaluó el gasto o recarga de agua, la información de la textura más arenosa registrada en el sitio RC sugiere un proceso de arrastre de partículas finas con un posible efecto de menor retención de humedad.
En los tres tipos de bosque coexisten diversas especies de Quercus, con una mayor abundancia en el BPQ (369 individuos), seguido de BS (226 individuos) y el sitio RC (33 individuos), que en su conjunto representan el 39% de la abundancia total. Lo anterior indica que el 61% restante de la flora leñosa corresponde a otras especies de latifoliadas y coníferas. La mayor diversidad de árboles registrada en el BPQ refleja la condición sucesional más avanzada de esta comunidad, a pesar de tener evidencia de extracción forestal. Las 20 especies de árboles registradas en el BPQ representan sólo la mitad de lo que pueden albergar los bosques maduros de la región (35-40 especies; González-Espinosa etal., 2006).
La escasez de plántulas en el sitio RC puede deberse a la falta de individuos reproductivos de otras especies de árboles diferentes de Cupressus, y por el posible efecto alelopático de C. lusitanica sobre la germinación de semillas o crecimiento de plántulas (Lines y Fournier, 1979; Pérez-Salicrup et al., 2006). Tampoco se observaron plántulas de C. lusitanica en ninguno de los tres bosques, a pesar de que la edad reproductiva de esta especie es entre los 10-12 años, aunque no se regeneran bajo su propia sombra y requiere de luz para su establecimiento exitoso (Chaves y Fonseca, 1991).
La estructura de tamaños observada en los tres tipos de vegetación sugiere que hay una amplia utilización de las especies para leña y madera y, que hay variaciones en intensidad y frecuencia. En los bosques de encino en México, incluyendo los Altos de Chiapas, la madera de Quercus se ha utilizado para la elaboración de carbón, por lo que muchas extensiones de bosques de encino de la región han disminuido por la tala selectiva (González-Espinosa et al., 2007, 2009). Ante la mayor ausencia de árboles grandes de encino en RC, es natural que el aprovechamiento sea más intensivo en el BPQ y BS.
Es importante señalar el valor de la reforestación de Cupressus para propósitos de recuperación y mantenimiento de la riqueza y diversidad de árboles. Contrario a lo esperado, no todas las condiciones estructurales y de calidad del sitio son inferiores a los otros dos bosques comparados. Una posible explicación es que el sitio RC nunca ha tenido un manejo silvícola para promover el crecimiento exclusivo de Cupressus. Después del establecimiento de la plantación, en 1984, sólo se han dado podas esporádicas en algunos sectores de La Milpoleta, pero se ha permitido el proceso de regeneración natural. Actualmente no está claro el destino que tendrá este sitio reforestado, y es necesario abrir la discusión ante los distintos sectores interesados (pobladores locales, autoridad municipal, organismos no gubernamentales), para decidir conjuntamente las acciones futuras para la conservación y manejo del área. Si se decide para propósitos productivos, es necesario aplicar un manejo silvícola conveniente; si es para propósitos de conservación, se pueden recomendar varias acciones. Por ejemplo, la escasa regeneración de encinos dentro del sitio reforestado con Cupressus implicaría repoblar con plántulas o semillas para incrementar su densidad y crecimiento en el área. En las áreas más densas, podrían crearse claros de copa para promover la liberación y crecimiento de los encinos ya establecidos, pero también mediante la repoblación con nuevas plántulas. Este sistema de enriquecimiento ya ha dado resultados favorables en algunas localidades de Chiapas (Ramírez-Marcial et al., 2006).
El análisis de la diversidad de árboles entre los tres tipos de bosque confirmó la tendencia general de un empobrecimiento florístico en el sitio reforestado con Cupressus respecto al BS y BPQ. Se ha reportado que en Los Altos de Chiapas, los bosques de encino análogos al BPQ pueden contener una riqueza estimada de 27 especies por hectárea y puede incrementarse conforme aumenta la edad sucesional de un bosque (González-Espinosa et al., 2006). Aunque el BPQ se consideró un bosque sin perturbación aparente, los resultados del presente estudio señalan que se trata de un bosque intervenido. La fisonomía del BPQ muestra una cobertura continua, pero la información de la estructura y diversidad indican una merma considerable respecto a los bosques con mayor estado de conservación en la región. Sin embargo, las condiciones actuales en el sitio reforestado con Cupressus constatan que la riqueza y diversidad de árboles es inferior a los bosques originales de pino-encino. Debido a que la reforestación con el ciprés tuvo como objetivo inicial conservar el suelo, propósito que hasta ahora parece haberse logrado parcialmente, es necesario incrementar la riqueza de especies bajo el dosel del sitio reforestado con Cupressus mediante prácticas de enriquecimiento con plántulas de otras especies tolerantes a la sombra. Asimismo, se propone que en futuros esfuerzos de reforestación se fomente el empleo de otras numerosas especies de árboles nativas de rápido crecimiento que contribuyan a mantener la estructura y diversidad de los bosques típicos de la región.
Conclusiones
Después de 28 años la reforestación de Cupressus lusitanica en La Milpoleta, ha provocado cambios notables en la composición y estructura en comparación con los bosques de pino-encino sin reforestación. Aunque se aprecian algunas diferencias en las propiedades del suelo en el sitio reforestado (mayor concentración de fósforo y suelos más arenosos), las condiciones parecen ser propicias para el crecimiento y desarrollo de especies arbóreas nativas. La ausencia de un plan de manejo del área reforestada señala la oportunidad para discutir las posibles rutas de desarrollo del área para propósitos de producción de madera o de conservación de suelos y biodiversidad. A la luz de las tendencias actuales de empobrecimiento florístico por deforestación en la región de estudio, se sugiere que la estrategia de conservación podría ser la más apropiada y, para ello, se propone la necesidad de sustituir algunos individuos del ciprés por otras especies arbóreas nativas de rápido crecimiento o que permitan el reclutamiento natural de otras especies.
Agradecimientos
Agradecemos a Miguel Martínez-Icó por su apoyo en el trabajo de campo y en la determinación de ejemplares botánicos y a los comentarios de dos revisores anónimos por sus atinadas observaciones a una versión previa. A las autoridades de La Milpoleta y El Aguaje por el acceso a las áreas de estudio. Este trabajo fue posible gracias al apoyo del Fondo Institucional de Fomento Regional para el Desarrollo Científico, Tecnológico y de Innovación (FORDECYT-CONACyT) por medio de los convenios 116306: Innovación socioambiental para el desarrollo en áreas de alta pobreza y biodiversidad de la frontera sur de México, y 143303: Gestión y estrategias de manejo sustentable para el desarrollo regional en la cuenca hidrográfica transfronteriza Grijalva.
Literatura citada
Bautista-Cruz A. y del Castillo R.F. 2005. Soil changes during secondary succession in a tropical montane cloud forest area. Soil Science Society of American Journal 69:906-914. [ Links ]
Cavelier J. y Tobler A. 1998. The effect of abandoned plantations of Pinus patula and Cupressus lusitanica on soils and regeneration of a tropical montane rain forest in Colombia. Biodiversity and Conservation 7: 335-347. [ Links ]
Cayuela L., Golicher D.J., Rey-Benayas J.M., González-Espinosa M. y Ramírez-Marcial N. 2006. Fragmentation, disturbance and tree diversity conservation in tropical montane forests. Journal of Applied Ecology 43:1172-1181. [ Links ]
Cervantes V, Carabias J. y Arriaga V. 2008. Evolución de las políticas públicas de restauración ambiental. En: CONABIO. Ed. Capital Natural de México. Vol. Ill: Políticas Públicas y Perspectivas de Sustentabilidad, pp: 155-226, Comisión Nacional para el Conocimiento y uso de la Biodiversidad, México, D.F. [ Links ]
Chaves E. y Fonseca W. 1991. Ciprés. Cupressus lusitanica Mill. Especie de Árbol de Uso Múltiple en América Central. Centro Agronómico Tropical para la Investigación y Enseñanza, Turrialba. [ Links ]
Cook J.G., Stutzman T.W., Bowers C.W., Brenner K.A y Irwin L.L.1995. Spherical densiometers produce biased estimates of forest canopy cover. Wildlife Society Bulletin 23:711-717. [ Links ]
Farjon A. 1993. Nomenclature of the Mexican Cypress or "Cedar of Goa", Cupressus lusitanica Mill. (Cupressaceae). Taxon 42:81-84. [ Links ]
Figueroa-Jáuregui M.L., Ibáñez-Castillo L.A., Arteaga-Ramírez R., Arellano-Monterrosas J.L. y Vázquez-Peña M. 2011. Cambio de uso del suelo en la cuenca de San Cristóbal de Las Casas, México. Agrociencia 45:531-544. [ Links ]
Galindo-Jaimes L. González-Espinosa M., Quintana-Ascencio P y García-Barrios L. 2002. Tree composition and structure in disturbed stands with varying dominant by Pinus spp. in the highlands of Chiapas, Mexico. Plant Ecology 162:259-272. [ Links ]
González-Espinosa M., Ramírez-Marcial N. y Galindo-Jaimes L. 2006. Secondary succession in montane pine-oak forests of Chiapas, Mexico. En: Kappelle M. Ed. Ecology and Conservation of Neotropical Montane Oak Forest. Ecological Studies Vol. 185, pp 209-221, Springer Verlag, Heidelberg. [ Links ]
González-Espinosa M., Ramírez-Marcial N., Camacho-Cruz A., Holz S.C, Rey-Benayas J.M. y Parra-Vázquez M.R. 2007. Restauración de bosques en territorios indígenas de Chiapas: modelos ecológicos y estrategias de acción. Boletín de la sociedad Botánica de México 80:11-23. [ Links ]
González-Espinosa M., Ramírez-Marcial N., Galindo-Jaimes L., Camacho-Cruz A., Golicher D., Cayuela L. y Rey-Benayas J.M. 2009. Tendencias y proyecciones del uso del suelo y la diversidad florística en Los Altos de Chiapas, México. Investigación Ambiental 1:40-53. [ Links ]
Guariguata M.R., Rheingans R. y Montagnini F. 1995. Early woody invasion under tree plantations in Costa Rica: implications for forest restoration. Restoration Ecology 3:252-260. [ Links ]
Konijnendijk C. 2008. The Forest and the City: The Cultural Landscape of Urban Woodland. Springer, Copenhagen. [ Links ]
Lemenih M., Olsson M. y Karltun E. 2004. Comparison of soil attributes under Cupressus lusitanica and Eucalyptus saligna established on abandoned farmlands with continuously cropped farmlands and natural forest in Ethiopia. Forest Ecology and Management 195:57-67. [ Links ]
Lemmon P.E. 1956. A spherical densiometer for estimating forest overstory density. Forest Science 2:314-320. [ Links ]
Lines M.N. y Fournier O.L.A. 1979. Efecto alelopático de Cupressus lusitanica Mill. sobre la germinación de las semillas de algunas hierbas. Revista de Biología Tropical 27:223-229. [ Links ]
Lugo A.E. 1997. The apparent paradox of reestablishing species richness on degraded lands with tree monocultures. Forest Ecology and Management 99:9-19. [ Links ]
Lugo A.E., Parrotta J.A. y Brown S. 1993. Loss of species caused by tropical deforestation and their recovery through management. Ambio 22:106-109. [ Links ]
Mueller-Dombois D. y Ellenberg H. 1974. Aims and Methods of Vegetation Ecology. Wiley, Nueva York. [ Links ]
Oksanen J. 2011. Multivariate analysis of ecological communities in R: vegan tutorial. <cc.oulu.fi/~jarioksa/opetus/metodi/vegantutor.pdf> (consultado 17 de octubre de 2012). [ Links ]
Parrotta J.A. 1992. The role of plantation forests in rehabilitating degraded tropical ecosystems. Agriculture Ecosystems and Environment 41:115-133. [ Links ]
Parrotta J.A., Turnbull J.W. y Jones N. 1997. Catalyzing native forest regeneration on degraded tropical lands. Forest Ecology and Management 99:1-7. [ Links ]
Pérez-Salicrup D., Peñaloza-Guerrero C. y Aguiar-Eleutério A.A. 2006. Regeneration of Styrax argenteus in natural forest and in plantations of Cupressus lindleyi in Michoacan, Mexico. New Forest 32:231-241. [ Links ]
Price M.F., Gratzer G., Alemayehu-Duguma L., Kohler T., Maselli D. y Romeo R. Eds. 2011. Mountain Forests in a Changing World - Realizing Values, Addressing Challenges. Food and Agriculture Organization of the United Nations-Swiss Agency for Development and Cooperation, Roma. [ Links ]
Price N.W. 1982. A comparison of water balance components in natural and plantation forests in El Salvador, Central America. Turrialba 32:399-416. [ Links ]
R Development Core Team. 2012. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna <www.r-project.org> (consulta: 17 de octubre 2012). [ Links ]
Ramírez-Marcial N., González-Espinosa M. y Williams-Linera G. 2001. Anthropogenic disturbance and tree diversity in montane rain forest in Chiapas, Mexico. Forest Ecology and Management 154:311-326. [ Links ]
Ramírez-Marcial N., Camacho-Cruz A., González-Espinosa M. y López-Barrera F. 2006. Establishment, survival and growth of tree seedlings under successional montane oak forests in Chiapas, Mexico. En: Kappelle M. Ed. Ecology and Conservation of Neotropical Montane Oak Forest, Ecological Studies Vol. 185, pp. 177-189, Springer Verlag, Heidelberg. [ Links ]
Rey-Benayas J.M., Newton A.C., Díaz A. y Bullock J.M. 2009. Enhancement of biodiversity and ecosystem services by ecological restoration: A meta-analysis. Science 325:1121-1124. [ Links ]
Richter D.D., Markewitz D., Trumbore S.E. y Wells C.G. 1999. Rapid accumulation and turnover of soil carbon in a re-establishing forest. Nature 400:56-58. [ Links ]
Rosenzweig M.L. 1995. Species Diversity in Space and Time. Cambridge university Press, Nueva York. [ Links ]
Ruiz-Jaen M.C. y Aide T.M. 2005. Restoration success: How is it being measured? Restoration Ecology 13:569-577. [ Links ]
Seppelt R., Dormann C.F., Eppink F.V. Lautenbach S. y Schmidt S. 2011. A quantitative review of ecosystem service studies: approaches, shortcomings and the road ahead. Journal of Applied Ecology 48:630-636. [ Links ]
Webster D.R. 2002. On the Edge: Shaping the Future of Periurban East Asia. Asia/Pacific Research Center, Stanford University, Stanford. [ Links ]

