










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBotanical Sciences
On-line version ISSN 2007-4476Print version ISSN 2007-4298
Bot. sci vol.91 n.1 México Mar. 2013
Botánica estructural
Viabilidad de polen, densidad y tamaño de estomas en autotetraploides y diploides de Physalis ixocarpa
Pollen viability, density and size of stomata in autotetraploids and diploids of Physalis ixocarpa
Francisca Ramírez-Godina1, Valentín Robledo-Torres2,4, Rahim Foroughbakhch-Pournavab3, Adalberto Benavides-Mendoza2 y Marco Antonio Alvarado-Vázquez3
1 Departamento de Fitomejoramiento, Universidad Autónoma Agraria Antonio Narro, Saltillo, Coahuila, México.
2 Departamento de Horticultura, Universidad Autónoma Agraria Antonio Narro, Saltillo, Coahuila, México.
3 Departamento de Botánica, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, Monterrey, Nuevo León, México.
4 Autor para la correspondencia: varoto@prodigy.net.mx
Recibido: 3 de marzo de 2012
Aceptado: 6 de julio de 2012
Resumen
Con el objetivo de estudiar el impacto de la autopoliploidía en la viabilidad de polen y el tamaño y frecuencia estomática del tomate de cáscara (Physalis ixocarpa) se analizó el polen, tamaño y número de estomas de cinco poblaciones diploides y cinco autotetraploides formadas con colchicina. Las diez poblaciones se establecieron en campo, bajo un diseño de bloques al azar con cuatro repeticiones. Para el estudio de viabilidad de polen, éste fue colectado y coloreado con acetocarmín al 1%. Los granos de polen redondeados y coloreados de rojo se consideraron viables y los constreñidos y sin teñir, no viables. Además, se estimó la densidad e índice estomático, largo y ancho de células oclusivas de estomas de la superficie adaxial y abaxial de la hoja. Los diploides tuvieron valores de viabilidad de polen y densidad estomática, significativamente mayores que los autotetraploides (P ≤ 0.05); sin embargo, los diploides tuvieron granos de polen y estomas significativamente más pequeños que los autotetraploides. Por lo tanto, la poliploidía si afectó la viabilidad del polen, ocasionado pérdida de fertilidad. Sin embargo, esta característica se puede evitar por selección de plantas con estabilidad meiótica, para incrementar la fertilidad de los autotetraploides. El aumento del tamaño celular muestra que los autopoliploides tienen el potencial de generar plantas más vigorosas, pero con una disminución en la fertilidad, aunque esta característica puede mejorarse al paso de varias generaciones, por lo que este tipo de estudios abre la posibilidad de iniciar nuevas estrategias para el mejoramiento del tomate de cáscara.
Palabras clave: abaxial, adaxial, granos de polen, poliploidía, tomate de cáscara.
Abstract
In order to study the impact of autopolyploidy in pollen viability and in stomatal features of husk tomato (Physalis ixocarpa), pollen was analyzed as well as the size and number of stomata of five diploid populations and five colchicines-formed autotetraploids populations. All ten populations were established on open ground, under a randomized block design with four replications. For the study of pollen viability, pollen was collected and stained with acetocarmin 1%. Rounded and red colored pollen grains were considered viable and those constrained and undyed, non-viable. In addition, the stomatal density and index were assessed as well as the length and width of stomata occlusive cells on the adaxial and abaxial leaf surface. The diploids showed significantly higher values of pollen viability and stomatal density than those of autotetraploids (P ≤ 0.05); however, the diploids had pollen grains and stomata significantly smaller than the autotetraploids. Therefore, the polyploidy did affect the viability of pollen, leading to the loss of fertility. However, this feature can be avoided by selecting plants with meiotic stability, to increase the fertility of the autotetraploids. Cell size increment shows that the autopolyploids have the potential to generate more vigorous plants, but with a decline in fertility; although fertility can be improved through several generations. This type of studies opens up the possibility of initiating new strategies for the improvement of husk tomato.
Keywords: abaxial, adaxial, husk tomato, pollen grains, polyploidy.
El tomate de cáscara (Physalis ixocarpa Brot. ex Hornem) se conoce en México desde tiempos precolombinos, los Aztecas lo cultivaban y empleaban como un componente importante de la cocina Mesoamericana, el cual es empleado de manera similar al tomate (Lycopersicon esculentum Mill.). Esta especie es nativa de México y América Central, actualmente es de gran importancia económica en México (Cantwell et al., 1992), en el 2010 fue la cuarta hortaliza en superficie sembrada, con una área de 48,475 ha (SIAP-SAGARPA, 2011). La importancia de esta hortaliza se debe a su alto consumo en México y a su exportación a los Estados Unidos de América y Canadá. También se cultiva en la India, Australia y Sudáfrica, así como en el sureste de los Estados Unidos (Fischer et al., 1990; Peña y Márquez, 1990). El rendimiento medio del tomatillo en México es de 15.58 t ha-1(SIAP-SAGARPA, 2011), el cual es bajo si se considera su potencial estimado en 40 t ha-1 (Peña y Santiaguillo, 1999).
A pesar de existir amplia variabilidad genética tanto en el tomate silvestre como en el domesticado en México (Santiaguillo et al., 2004), el rendimiento medio nacional es bajo, posiblemente como consecuencia del limitado mejoramiento genético de la especie, el uso de variedades nativas y sistemas tradicionales de producción. Sin embargo, esta variabilidad se ha aprovechado por los fitomejoradores mediante métodos de selección masal visual estratificada, la cual ha sido eficiente para incrementar el rendimiento, por lo que se requiere desarrollar métodos de mejoramiento más eficientes que respondan a las necesidades del mercado y de los productores (Peña et al., 2002).
Otra alternativa de introducción de nuevo potencial genético en el tomatillo es la duplicación genómica. La autopoliploidía es un estado biológico inducible caracterizado por redundancia genómica, el cual puede ser aprovechado por los fitomejoradores, además incrementa el tamaño efectivo de la población y la flexibilidad genómica, facilitando así el manejo de la selección artificial. La redundancia genética puede permitir la divergencia adaptativa de genes duplicados (Parisod et al., 2010), además la duplicación cromosómica causa un incremento en la altura de plantas y en la longitud, ancho, espesor y volumen foliar con respecto a plantas diploides (Molero-Paredes y Matos, 2008). Sin embargo, se ha encontrado que los frutos de los poliploides tienen reducida producción de semilla, como consecuencia de irregularidades meióticas, que llevan a la pérdida de cromosomas en anafase I y II, llevando a la planta a tener pérdida de fertilidad (Alfonsi y Cequea, 2000). Por lo tanto, la formación de tetraploides en Physalis ixocarpa abre nuevas posibilidades en la selección y mejora de esta especie, ya que, se sabe que la duplicación del genoma generalmente induce cambios en rasgos fenotípicos y anatómicos, tales como el incremento en el tamaño celular y modificaciones en la tolerancia ecológica (Parisod et al., 2010), aunque se ha encontrado que la pérdida de fertilidad en autopoliploides es relevante, ya que influye significativamente en el desarrollo del fruto, que es el principal órgano de valor económico. La reducción de la fertilidad disminuye la fecundación y, por lo tanto, la producción de semilla, la cual está relacionada con la síntesis de giberelinas y auxinas, reguladores vegetales que promueven la división y elongación celular y el desarrollo del fruto (Bünger-Kibler y Bangerth, 1982). En programas de mejoramiento, el conocimiento de la viabilidad del polen es uno de los factores esenciales, sobre todo cuando se utilizan técnicas de hibridación artificial, como es el caso del tomate de cáscara. La determinación de la viabilidad del polen y su importancia en estudios de la biología de la reproducción y mejoramiento genético de plantas, es fundamental para definir la dirección de una cruza y contar con bases para el éxito de hibridaciones controladas para garantizar la formación de nuevos híbridos e incrementar su viabilidad (Salles et al., 2006). De ahí la necesidad de estimar la viabilidad el polen en autotetraploides de tomate de cáscara. Con el incrementó de ploidia hay modificación en el tamaño celular, por lo que es necesario estudiar estructuras celulares como los estomas que están relacionados con importantes funciones fisiológicas como la fotosíntesis y la respiración, relacionadas con el rendimiento de fruto y acumulación de biomasa en plantas. Costa et al. (2003) indican que los estomas son más grandes y menos frecuentes en cítricos poliploides que en sus progenitores diploides. Considerando lo anterior, los estomas con amplios poros estomáticos facilitan altas tasas de intercambio de CO2 (Franks y Farquhar, 2007).
Dada la importancia que tiene la fertilidad y los estomas en la productividad de los cultivos, así como la variabilidad de los autopoliploides de tomate de cáscara, en el rendimiento del fruto y la sobreexpresión de caracteres relacionados con el rendimiento de fruto (Robledo-Torres et al., 2011), resulta importante estudiar las características de los estomas y viabilidad de polen que están relacionados con el rendimiento y calidad de fruto. Los autotetraploides son utilizados en la reproducción del tomate de cáscara por medio de cruzamientos con diploides para generar triploides con niveles satisfactorios de rendimiento y calidad de fruto, como los obtenidos en mandarina, melón y sandía (Nugent y Adelberg, 1995; Rhodes y Zhang, 2000; Aleza et al., 2009). Sin embargo, hay poca información disponible sobre la viabilidad del polen de materiales poliploides. Por lo tanto el objetivo de esta investigación fue hacer un análisis del polen y de las características (tamaño y número) de los estomas en autotetraploides y diploides de tomate de cáscara para conocer el impacto de la poliploidía en la viabilidad de polen y características estomáticas.
Materiales y métodos
El experimento se llevó a cabo en General Cepeda, Coahuila, México, en el invernadero de la Universidad Autónoma Agraria Antonio Narro y Laboratorio de Citogenética de la misma Universidad en Saltillo, Coahuila, México, en el 2010. Para el estudio de estomas y granos de polen se utilizaron cinco poblaciones tetraploides: 2, 5, 11, 16 y 20, formadas por acción de la colchicina (Robledo-Torres et al., 2011) y cinco diploides originarias de México: (19) Variedad Rendidora (origen Zacatepec, Morelos) de la cual se formaron las poblaciones autotetraploides, (13) Gran Esmeralda (material mejorado, comercializado por la empresa Harris Moran), (1) criollo colectado en Felipe Ángeles (Acatzingo, Puebla), (21) criollo colectado en Palmarito (Quecholac, Puebla) y el (18) criollo morado tamazula colectado en (Arandas, Jalisco). El nivel de ploidía de estos materiales fue analizado con anterioridad por conteos cromosómicos en mitosis y meiosis y análisis por citometría de flujo del contenido de ADN nuclear (Robledo-Torres et al., 2011).
Las poblaciones se establecieron en campo bajo el diseño experimental de bloques al azar con cuatro repeticiones, teniendo como parcela útil tres plantas con competencia completa. Las variables estudiadas fueron; viabilidad (VP) y diámetro de polen (DP), densidad estomática abaxial (DEAb), índice estomático abaxial (IEAb), de acuerdo con el método de Cañizares et al. (2003) se evaluó el largo y ancho de las células oclusivas incluyendo el poro que componen el aparato estomático o estomas de la superficie abaxial (LEAb y AEAb), densidad estomática adaxial (DEAd), índice estomático adaxial (IEAd), largo y ancho de las células oclusivas incluyendo el poro de los estomas de la superficie adaxial (LEAd y AEAd).
Se sembraron 100 semillas por genotipo en charolas de poliestireno con turba canadiense como sustrato. Posteriormente las charolas se llevaron a invernadero para el desarrollo de las plántulas y 30 días después de la siembra, cuando alcanzaron de 10 a 12 cm de altura y dos pares de hojas verdaderas, fueron llevadas a campo para su trasplante en un suelo con acolchado plástico y riego por goteo.
Análisis de polen. Para el análisis de polen, 30 días después del trasplante se tomaron florecillas de tres plantas por población, de cada una de las cuatro repeticiones. La viabilidad de polen, previa a la antesis, fue estimada en los tetraploides y diploides, de la siguiente forma: se colectaron tres flores por planta entre 8:00 y 9:30 de la mañana, se colocaron en bolsas de papel estraza y se llevaron al laboratorio para su análisis. La tinción con carmín acético se realizó de la siguiente forma: se disolvieron 45 ml de ácido acético puro en 55 ml de agua destilada y se vertieron en un matraz, en el que previamente se habían adicionado limaduras de hierro y se mantuvieron en ebullición durante 4 min, se añadió 1 g de carmín por 100 ml de disolución, se continuó con la ebullición durante 1 min y se filtró. Se impregnó un pincel con polen de cada flor y se sacudió sobre un portaobjetos procurando que la distribución fuera uniforme, se depositó una gota de colorante acetocarmín al 1%, encima se colocó un cubreobjetos y después de 5 a 10 seg se procedió a la observación al microscopio. Para el recuento se realizaron dos preparaciones por flor con el polen recién colectado. La viabilidad de polen se estimó considerando los granos de polen redondeados y coloreados de rojo viables (Figura 1) y los constreñidos y sin teñir, no viables de acuerdo con Stone et al. (1995), Lagos et al. (2005) y Srinivasan y Gaur (2012). Los conteos de polen se efectuaron en dos campos del microscopio por preparación, se contabilizó el número de granos de polen viables e inviables y se estimó el porcentaje de viabilidad (Cardoso et al., 2004; Soares et al., 2008). Además se midió el diámetro en (μm) de nueve granos de polen por planta de tres plantas por población en cada una de las cuatro repeticiones, lo que representa un total de 108 granos de polen por población. Todas las observaciones se hicieron con el objetivo 40X en un microscopio compuesto (Carl Zeiss) con cámara digital (Pixera Winder Pro) y un software de medición (Axion Vision Rel. 4.8).
Estudio de estomas. Para el estudio de estomas, se tomaron muestras a los 34 días después del trasplante, se utilizaron tres plantas por población en cada una de las cuatro repeticiones, de las que se tomaron dos hojas maduras, totalmente expandidas de la primera rama y con la misma orientación. En cada hoja, de su parte media, se tomó una impresión epidérmica del haz (adaxial) y otra del envés (abaxial), utilizando poliestireno-xilol en forma líquida, el que se aplicó sobre la superficie foliar con un pincel. Después de que se secó la película, ésta fue removida con un trozo de cinta adhesiva transparente y se montó sobre un portaobjetos. En cada impresión se observaron al azar tres campos microscópicos a 40X, analizando 72 campos del microscopio por población, de la superficie adaxial y 72 de la superficie abaxial. De cada campo se tomó una microfotografía, en la que se contaron los estomas y las células epidérmicas, además se midió el ancho y largo (μm) de las células oclusivas de dos estomas en cada una de los campos del microscopio, de manera que se midieron 144 estomas de la superficie adaxial y 144 de la superficie abaxial por población.
La densidad estomática se obtuvo de la siguiente forma: DE = Número de estomas / 0.02479 mm2 (área de la imagen fotografiada) = estomas por mm2.
Para sacar el índice estomático se utilizó la siguiente fórmula: IE = N° de estomas / células epidérmicas + N° de estomas x 100 (Wilkinson, 1979). Se utilizó un microscopio compuesto (Carl Zeiss) con cámara digital (Pixera Winder Pro) y un software de medición (Axion Vision Rel. 4.8).
Análisis estadístico. Se realizó un análisis de varianza en un diseño de bloques al azar con cuatro repeticiones donde el factor de variación fueron cinco poblaciones de autotetraploides y cinco diploides de tomate de cáscara para la comparación de medias se utilizó la prueba de Tukey (P ≤ 0.05). Se utilizó el programa estadístico de SAS versión 9.0 (Cary, EUA).
Resultados
Análisis de polen. El análisis de varianza aplicado a la viabilidad y diámetro de polen, exhibió diferencias significativas entre poblaciones (F = 24.70, P ≤ 0.01) para VP y (F = 22.09, P ≤ 0.01) en DP, indicando que el tamaño y la fertilidad de polen fueron afectados por el incremento del nivel de ploidía. Al analizar los valores medios de las características estudiadas, se encontró que las poblaciones diploides presentaron; 90.16 a 91.71% de viabilidad de polen, mientras que los tetraploides presentaron; 54.80 a 69.77%. En los diploides se observó aproximadamente 23% más viabilidad de polen que los tetraploides (Cuadro 1). La comparación de medias mostró que en viabilidad de polen, existen diferencias significativas entre poblaciones diploides y autotetraploides (P ≤ 0.05), aunque dentro de las poblaciones autotetraploides no se encontraron diferencias significativas, como tampoco se encontraron diferencias entre poblaciones diploides. Por otra parte las poblaciones diploides presentaron un diámetro de polen de 23.42 a 25.53 μm y los tetraploides de 28.75 a 31.10 μm (Figura 1), mostrando diferencias significativas (P ≤ 0.05) entre poblaciones tetraploides y diploides, sólo la población diploide 13 y la población tetraploide 2 fueron estadísticamente iguales (Cuadro 1).

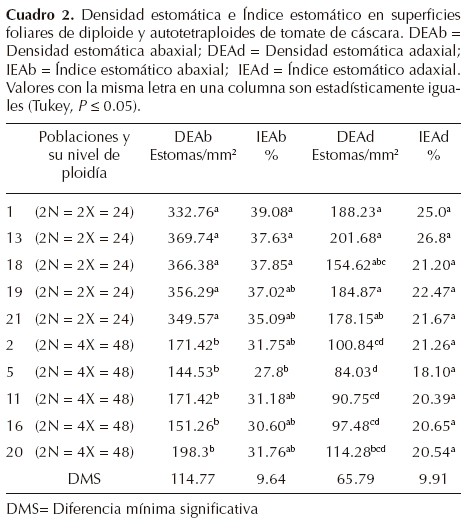
Estudio de estomas. El análisis de varianza aplicado a la densidad estomática abaxial y adaxial de la hoja, mostró diferencias significativas entre poblaciones diploides y tetraploides, en DEAb (F =18.08, P ≤ 0.01) y para DEAd (F = 11.77, P ≤ 0.01). Sin embargo, para el índice estomático sólo hubo diferencias significativas entre poblaciones con un valor de (F = 3.73, P ≤ 0.05), y no se encontraron diferencias significativas entre poblaciones diploides y tetraploides en el índice estomático adaxial (F = 1.47, P = 0.20).
Al analizar los valores medios la DEAb, se encontró que las poblaciones diploides presentaron de 349.57 a 369.74 estomas/mm2 en la superficie abaxial de la hoja, mientras que en las poblaciones tetraploides se observó de 144.53 a 198.31 estomas/mm2. La comparación de medias mostró que entre poblaciones diploides fueron estadísticamente iguales en DEAb (Cuadro 2), pero estadísticamente diferentes a los tetraploides (P ≤ 0.05). En la superficie adaxial de la hoja de los diploides, se encontró de 154.62 a 201.68 estomas/mm2, mientras que en los tetraploides fue de 84.03 a 114.28 estomas/mm2. Los diploides presentaron aproximadamente 112% más estomas en la parte abaxial de la hoja que los tetraploides, mientras que en la parte adaxial los diploides presentaron un 86% más estomas que los tetraploides (Figura 2). Los diploides en la región abaxial de la hoja presentaron un índice estomático de 35.09 a 39.08%, mientras que en los tetraploides fue de 27.81 a 31.76%. El índice estomático adaxial en los diploides fue de 21.20 a 26.81%, mientras que en los tetraploides de 18.10 a 21.26%.
El análisis de varianza aplicado a las variables AEAb, LEAb, AEAd y LEAd, presentó diferencias significativas entre poblaciones con valores de (F = 14.67, 25.29, 7.12, 10.57 todos con P ≤ 0.01) respectivamente, indicando que por lo menos una población es estadísticamente diferente del resto. Las poblaciones diploides mostraron estomas más pequeños que los tetraploides, tanto en la superficie abaxial como adaxial (Figura 4), ya que la longitud media abaxial en los diploides fue de 28.88 a 32.31 μm y de 19.44 a 21.80 μm de ancho, mientras que los tetraploides presentaron una longitud de 42.10 a 46.59 μm y un ancho de 26.77 a 30.02 μm. Las células oclusivas de los estomas de la superficie abaxial de los tetraploides fueron en promedio 46% más largas y 35% más anchas que las células oclusivas de los estomas de los diploides. En la superficie adaxial los diploides presentaron una longitud de 26.32 a 32.03 μm y un ancho de 15.44 a 18.89 μm, mientras que los tetraploides tuvieron estomas con una longitud de 41.25 a 45.04 μm y un ancho de 21.43 a 23.15 μm. Esto indica que la longitud media de los estomas en la superficie adaxial de los tetraploides fue 45% mayor que la de los diploides y los tetraploides fueron 31% más anchos que los estomas adaxiales de los diploides (Figura 3).
La comparación de medias para la longitud de estomas de la superficie abaxial mostró que los estomas de los diploides fueron significativamente más cortos que los estomas de los tetraploides y estadísticamente iguales entre sí, pero estadísticamente diferentes (P ≤ 0.05) de los tetraploides (Cuadro 3). En relación al ancho, de estomas abaxial, todos los tetraploides fueron estadísticamente iguales, aunque el tetraploide 11 fue estadísticamente igual al diploide 13. Los estomas de la superficie adaxial de los tetraploides fueron estadísticamente iguales en longitud. El ancho de estomas de la superficie adaxial separa claramente los diploides de los tetraploides y únicamente el tetraploide 2 fue estadísticamente igual que los diploides.

Discusión
Análisis de polen. Se observó que la duplicación del genoma en tomatillo produjo una reducción en la viabilidad del polen. Estos resultados son consistentes con los obtenidos en otras especies autopoliploides (Arbo y Fernández, 1983; Fernández, 1987). Los porcentajes más bajos de viabilidad del polen observados en algunas poblaciones poliploides son el resultado de irregularidades meióticas que conducen a la producción de gametos no balanceados e inviables. He et al. (2010) mencionan que la duplicación del genoma juega un papel importante en el desarrollo de polen normal en arroz.
De acuerdo con el criterio de Stebbins (1947) los autopoliploides están caracterizados por la presencia de multivalentes en meiosis, dando como consecuencia fertilidad reducida, por lo tanto baja viabilidad de polen. La determinación de la viabilidad del polen permite hacer estimaciones confiables de la fertilidad, además de utilizarse para el estudio de incompatibilidad en los cruzamientos (González et al., 1992).
En relación al diámetro de polen, se observó en los autotetraploides un aumento significativo en el tamaño de granos de polen en comparación con los diploides tal como se esperaba, se pudo corroborar el nivel de ploidía de los materiales diploides y tetraploides ya que de acuerdo con González et al. (2002), esta característica es un indicador de la ploidía en papa y constituye un método indirecto que permite la identificación de materiales diploides y tetraploides, aunque el nivel de ploidía de estos materiales ya se tenía comprobado con anterioridad por análisis citogenético y citometría de flujo (Robledo-Torres et al., 2011).
Estudio de estomas. Se encontró que los autotetraploides presentaron estomas más grandes, pero en menor cantidad por superficie foliar, en comparación con los diploides, por lo tanto, los tetraploides presentaron mayor tamaño de estomas que los diploides, en este sentido se concluye que el tamaño estomático es proporcional al nivel de ploidía, y la densidad estomática es inversamente proporcional al nivel de ploidía, coincidiendo con lo indicado por Borges (1971) quien señala que la densidad estomática en la superficie de las hojas es inversamente proporcional al nivel de ploidía. De igual forma Valerio et al. (2002) y Vandenhout et al. (1995), encontraron que los niveles más bajos de ploidía presentaban densidades estomáticas más altas y estomas pequeños, y a la inversa los niveles de ploidía más altos presentan menor densidad estomática y estomas grandes por superficie foliar. Por otro lado, se menciona que la densidad estomática e índice estomático son afectados por condiciones estresantes, tanto ambientales como nutricionales y que además dependen de la superficie de la hoja y de la especie (Bethke y Drew, 1992; Cañizares et al., 2003). Aunque se indica que la densidad estomática es una característica diagnóstica muy utilizada en sistemática de plantas, dado que generalmente se mantiene sin alteraciones (Wilkinson, 1979).
En este trabajo se observaron diferencias entre diploides y tetraploides a pesar de haberse desarrollado bajo las mismas condiciones ambientales, o bien las condiciones ambientales afectaron de forma diferente a los diploides y tetraploides.
Además se menciona que la longitud de las células oclusivas de los estomas, siempre es un método conveniente y confiable para la clasificación de plantas haploides, diploides, tetraploides y hexaploides (Aryavand et al., 2003; Rêgo et al., 2011) por lo tanto y de acuerdo con los resultados de tamaño y número de estomas por unidad de área, fue posible separar los diploides de los tetraploides en este estudio.
Borges (1971) menciona que el mayor tamaño de las células de los poliploides y de manera específica las células oclusivas tienen núcleos con una dotación duplicada de cromosomas lo cual induce células más largas y más anchas, mientras que la densidad estomática, en general, se reduce así como la transpiración, por lo tanto los poliploides toleran mejor los ambientes más secos que el diploide. Los resultados obtenidos permiten concluir que la duplicación del genoma en tomatillo promovió la reducción de la densidad estomática y el índice estomático, pero aumento el tamaño de estomas en las poblaciones autotetraploides estudiadas. En este caso, los mayores valores de índice estomático tanto de diploides como autotetraploides coincidieron con los mayores valores de densidad estomática.
La autopoloploidia en el tomate de cáscara causó una disminución en la viabilidad de polen y un incremento en el tamaño de granos de polen, en comparación con las plantas diploides. Por lo tanto, la poliploidía si afectó la viabilidad del polen, ocasionado pérdida de fertilidad, característica que podría ser mejorada por selección de plantas con apareamiento estable en meiosis, para incrementar la fertilidad de los autotetraploides.
En los autotetraploides se redujo el número de estomas/ mm2 y aumentó el tamaño de los estomas, demostrándose lo contrario en los diploides. Aunque la variabilidad de la densidad y el tamaño de los estomas de Physalis ixocarpa podrían atribuirse, en parte, al nivel de ploidía, no debería descartarse una posible influencia ambiental. El mayor tamaño celular demuestra que los autopoliploides tienen el potencial de generar plantas más vigorosas, pero una disminución en la viabilidad de polen, característica que puede mejorarse al paso de varias generaciones, por lo que este tipo de estudios abre la posibilidad de iniciar nuevas estrategias para el mejoramiento del tomate de cáscara.
Literatura citada
Aleza G.P., Cuenca J., Juárez R.J., Pina J.A. y Navarro L. 2009. Mandarinos "Garbí" y "Safor", dos nuevos híbridos triploides obtenidos en el IVIA. Levante Agrícola: Revista Internacional de Cítricos 395:85-87. [ Links ]
Alfonsi C. y Cequea H. 2000. Cytogenetic analysis of the artificial tetraploid Lycopersicon esculentum var. ceraciforme. Ciencia. Universidad del Zulia-Venezuela 8:119-126. [ Links ]
Arbo M.M. y Fernández A. 1983. Posición taxonómica, citología y palinología de tres niveles de ploidía de Turnera subulata Sm. Bonplandia 5:211-226. [ Links ]
Aryavand A., Ehdaie B., Tran B. y Waines J.G. 2003. Stomatal frequency and size differentiate ploidy levels in Aegilops neglecta. Genetic Resources and Crop Evolution 50:175-182. [ Links ]
Bethke P.C. y Drew M.C. 1992. Stomatal and nonstomatal competens to inhibition of photosynthesis in leaves of Capsicum annuum during progressive exposure to NaCl salinity. Plant Physiology 99:219-226. [ Links ]
Borges F.O.L. 1971. Tamaño y densidad de estomas en clones cultivados y especies silvestres de Musa. Agronomía Tropical 21:139-143. [ Links ]
Bünger-Kibler S. y Bangerth F. 1982. Relationship between cell number, cell size and fruit size of seeded fruits of tomato (Lycopersicon esculentum Mill.), and those induced parthenocarpically by the application of plant growth regulators. Plant Growth Regulation 1:143-154. [ Links ]
Cantwell M., Flores-Minutti J. y Trejo-González A. 1992. Developmental changes and postharvest physiology of tomatillo fruits (Physalis ixocarpa Brot.). Scientia Horticulturae 50:59-70. [ Links ]
Cañizares A., Sanabria M.E., Rodríguez D.A. y Perozo Y. 2003. Características de los estomas, índice y densidad estomática de las hojas de lima Tahití (Citrus latifoliaTanaka) injertada sobre ocho patrones cítricos. Revista Científica UDO Agrícola 3:59-64. [ Links ]
Cardoso M.B., Kaltchuk-Santos E., Mundstock E.C. y Bodanese-Zanettini M.H. 2004. Initial segmentation patterns of microspores and pollen viability in soybean cultured anthers: indication of chromosome doubling. Brazilian Archives of Biology and Technology 47:703-712. [ Links ]
Costa M.A.P, Mendes B.M.J. y Mourão-Filho F.A.A. 2003. Somatic hybridization for improvement of citrus rootstock: production of five combinations with potential for improvement disease resistance. Australian Journal Experimental Agriculture 43:1151-1156. [ Links ]
Fernández A. 1987. Estudios cromosómicos en Turnera y Piriqueta (Turneraceae). Bonplandia 6:1-21. [ Links ]
Fischer G., Buitrago M. T. y Ludders P 1990. Physalis peruviana L. cultivation and investigation in Colombia. Erwerbs-obstbau 32:229-232. [ Links ]
Franks P.J. y Farquhar G.D. 2007. The mechanical diversity of stomata and its significance in gas-exchange control. Plant Physiology 143:78-87. [ Links ]
González M.E., Estévez A., Rodríguez T. y Álvarez M. 1992. Estudio de la fertilidad del polen en especies de papa. Cultivos Tropicales 13:70-73. [ Links ]
González M.E., Estévez A., Castillo J., Salomón J., Moré O. y Hernández M.M. 2002. La calidad del polen: Requisito indispensable del mejoramiento tradicional de la papa en Cuba. Revista Latinoamericana de la Papa 13:75-94. [ Links ]
He Y., Wei Q., Ge J., Jiang A., Gan L., Song Z. y Cai D. 2010. Genome duplication effects on pollen development and the interrelated physiological substances in tetraploid rice with polyploid meiosis stability. Planta 232:1219-1228. [ Links ]
Lagos T.C., Caetano C.M., Vallejo F.A., Muñoz J.E., Criollo H. y olaya C. 2005. Caracterización palinológica y viabilidad polínica de Physalis peruviana L. y Physalis philadelphica Lam. Agronomía Colombiana 23:55-61. [ Links ]
Molero-Paredes T. y Matos A. 2008. Efectos de la inducción artificial de la poliploidia en plantas de Aloe vera (L.). Boletín del Centro de investigaciones Biológicas, Universidad del Zulia 42:111-133. [ Links ]
Nugent P.E. y Adelberg J. 1995. Fruit characteristics of hybrid triploid melon. HortScience 30:821-822. [ Links ]
Parisod C., Holderegger R. y Brochmann C. 2010. Evolutionary consequences of autopolyploidy. New Phytologist 186:5-17. [ Links ]
Peña L.A. y Márquez S.F. 1990. Mejoramiento genético de tomate de cáscara (Physalis ixocarpa Brot.). Revista Chapingo 71,72:85-88. [ Links ]
Peña L.A. y Santiaguillo H.J.F. 1999. Variabilidad genética de tomate de cáscara en México. Boletín Técnico Núm. 2. Departamento de Fitotecnia, Universidad Autónoma Chapingo, Chapingo. [ Links ]
Peña L.A., Molina G.J.D., Márquez S.F., Sahagún C.J., Ortiz C.J. y Cervantes S.T. 2002. Respuestas estimadas y observadas de tres métodos de selección en tomate de cáscara (Physalis ixocarpa Brot.) Revista Fitotecnia Mexicana 25:171-178. [ Links ]
Rêgo M.M., Rêgo E.R., Bruckner C.H., Finger F.L. y Otoni W.C. 2011. in vitro induction of autotetraploids from diploid yellow passion fruit mediated by colchicine and oryzalin. Plant Cell, Tissue and Organ Culture 107:451-459. [ Links ]
Rhodes B. y Zhang X. 2000. Hybrid seed production in watermelon. Journal of New Seeds 1:69-88. [ Links ]
Robledo-Torres V., Ramírez-Godina F., Foroughbakhch-Pournavab R., Benavides-Mendoza A., Hernández-Guzmán G. y Reyes-Valdés M.H. 2011. Development of tomatillo (Physalis ixocarpa Brot.) autotetraploids and their chromosome and phenotypic characterization. Breeding Science 61:288-293. [ Links ]
Salles L.A., Ramos J.D., Pasqual M., Junqueira K.P. y Bortolotti da Silva A. 2006. Sacarose e pH na germinação in vitro de grãos de pólen de citros. Ciência e Agrotecnológia 30:170-174. [ Links ]
Santiaguillo H.J.F., Cervantes S.T. y Peña L.A. 2004. Selección para rendimiento y calidad de fruto de cruzas planta x planta entre variedades de tomate de cáscara. Revista Fitotecnia Mexicana 27:85-91. [ Links ]
SIAP-SAGARPA. 2011. Servicio de Información y Estadística Agroalimentaria y Pesquera. http://www.siap.gob.mx/index.php?option=com_content&view=article&id=10&Itemid=15 [ Links ]
Srinivasan S. y Gaur P.M. 2012. Genetics and characterization of an open flower mutant in chickpea. Journal of Heredity 103:297-302. [ Links ]
Soares T.L., Oliveira e Silva S., Pereira de Carvalho C.M.A., Almeida dos Santos-Serejo J., Silva S.A., Morais L.L.S., Hilo de Souza E. y Nunes de Jesus O. 2008. In vitro germination and viability of pollen grains of banana diploids. Crop Breeding and Applied Biotechnology 8:111-118. [ Links ]
Stebbins G.L. Jr. 1947. Types of polyploids: their classification and significance. Advances in Genetics 1:403-429. [ Links ]
Stone J.L., Thomson J.D. y Dent-Acosta S.J. 1995. Assessment of pollen viability in hand-pollination experiments: A review. American Journal of Botany 82:1186-1197. [ Links ]
Vandenhout H., Ortiz R., Vuylsteke D., Swennem R. y Bai K.V. 1995. Effect of ploidy on stomatal and other quantitative traits in plantain and banana hybrids. Euphytica 83:117-122. [ Links ]
Valerio R., Lindorf H. y García de G.E. 2002. Anatomía foliar comparada de ocho cultivares de banano con relación a la resistencia o susceptibilidad a la Sigatoka (amarilla y negra). Agronomía Tropical 52:507-521. [ Links ]
Wilkinson H.P 1979. The plant surface (mainly leaf). En: Metcalfe C.R. y Chalk L. Eds. Anatomy of the Dicotyledons, 2nd ed. Vol. 1, pp. 97-165, Clarendon Press, Oxford. [ Links ]

