










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkIngeniería agrícola y biosistemas
versão On-line ISSN 2007-4026versão impressa ISSN 2007-3925
Ing. agric. biosist. vol.11 no.1 Chapingo Jan./Jun. 2019 Epub 24-Fev-2020
https://doi.org/10.5154/r.inagbi.2018.05.011
Artículo científico
Comportamiento agronómico de ecotipos de chile piquín bajo cubiertas fotoselectivas
1Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro núm. 1923, Saltillo, Coahuila, C. P. 25315, MÉXICO.
2Universidad Autónoma de Nuevo León. San Nicolás de los Garza, Nuevo León, C. P. 66455, MÉXICO.
Introducción:
El uso de malla sombra de colores e invernaderos es una estrategia para aprovechar la radiación solar y regular la temperatura, además de incrementar la productividad de los cultivos hortícolas.
Objetivo:
Evaluar el efecto de diferentes cubiertas fotoselectivas sobre variables agronómicas en ecotipos de chile piquín silvestre.
Metodología:
Las variables microclimáticas y radiación fotosintéticamente activa (RFA) se midieron desde el trasplante de la planta, y las variables agronómicas a los 40 días después del trasplante y al último corte del ciclo (160 días).
Resultados:
La temperatura, humedad relativa y RFA cambiaron con el uso de las diferentes cubiertas. La malla azul redujo 50.10 % la RFA en comparación con la incidente en campo abierto. El análisis de componentes principales agrupó a las plantas cultivadas sin cubierta con los menores resultados en las variables evaluadas, y el ecotipo RTZ reveló mayor tendencia hacia el cuadrante 1 en la mayoría de las variables. El rendimiento con malla blanca incrementó 1 290.9 % en comparación con el obtenido en campo.
Limitaciones del estudio:
El estudio únicamente genera información de los recursos fitogenéticos propios de la Región Noreste de México.
Originalidad:
El cultivo no está domesticado, por lo que este trabajo aporta el estudio bajo diferentes cubiertas de malla sombra y un invernadero.
Conclusiones:
La malla blanca favoreció el desarrollo de los ecotipos, siendo RTZ el que presentó los mejores valores, por lo que se considera viable esta combinación para la preservación y producción a gran escala.
Palabras clave malla sombra; rendimiento; Capsicum annuum
Introduction:
The use of colored shading nets and greenhouses is a strategy to improve the use of solar radiation and regulate temperature, in addition to increasing productivity in horticultural crops.
Objective:
To evaluate the effect of different photoselective covers on agronomic variables in wild piquin pepper ecotypes.
Methodology:
Microclimatic variables and photosynthetically active radiation (PAR) were measured from transplanting seedlings, and agronomic variables at 40 days after transplant and at the last cut of the cycle (160 days).
Results:
Temperature, relative humidity and PAR changed with the use of the different covers. Blue netting reduced PAR by 50.10 % compared to the open field. Principal component analysis grouped the uncovered cultivated plants with the lowest results in the evaluated variables, and the RTZ ecotype revealed the greatest tendency towards quadrant 1, with most of the variables. The yield in the white netting increased 1 290.9 % compared to that obtained in the field.
Study limitations:
The study generated information only for the plant genetic resources of the Northeast Region of Mexico.
Originality:
The crop is not domesticated, so this work was done under different shade netting covers and a greenhouse.
Conclusions:
White netting favored the development of the ecotypes, with RTZ being the one that presented the best values, which is why this combination is considered viable for the preservation and production of this species on a large scale.
Keywords shade netting; yield; Capsicum annuum
Introducción
El chile piquín es una especie silvestre de gran importancia desde el punto de vista económico y alimenticio. Su preferencia se debe a su agradable sabor y picor, comparado con el de otros chiles como el serrano y el jalapeño (Villalón-Mendoza, Medina-Martínez, Ramírez-Meraz, Solis-Urbina, & Maiti, 2014). Esta planta se encuentra distribuida de forma silvestre en México y en el sur de Estados Unidos de América, lo que demuestra su amplia adaptabilidad ambiental, probablemente asociada con su diversidad genética. De acuerdo con su naturaleza, los frutos se obtienen por recolección en poblaciones silvestres, y no de plantaciones comerciales, lo que disminuye las poblaciones naturales y amenaza la diversidad genética de esta especie (Rueda-Puente et al., 2010).
Por otro lado, las condiciones ambientales modifican la fenología de las plantas; específicamente, las poblaciones silvestres de chile piquín son perennes y comúnmente se asocian a condiciones parcialmente sombreadas en el sotobosque, pero cuando se siembran en campo abierto se pueden considerar cultivos anuales debido a las heladas, las enfermedades y la susceptibilidad a plagas (Rodríguez-del Bosque et al., 2003). Además, en los últimos años, el calentamiento global ha ocasionado cambios en las condiciones ambientales, como la temperatura y la precipitación (Conde-Álvarez & Saldaña-Zorrilla, 2007). Para mitigar dichos cambios se pueden utilizar mallas sombra o cubiertas plásticas que, además de generar cierta cantidad de sombra, simulan su hábitat silvestre y propician el desarrollo normal de la planta, lo que incrementa el rendimiento y la calidad del fruto en comparación con la producción en campo abierto (Ayala-Tafoya et al., 2011).
Lo anterior es de suma importancia ya que las plantas que no están adaptadas a la luz solar intensa pueden desarrollar estrés por calor. La mayoría de los efectos negativos pueden evitarse con la protección de un invernadero o con estructuras de sombreado (Castronuovo, Statuto, Muro, Picuno, & Candido, 2017).
Con la finalidad de preservar esta especie e incrementar su cultivo comercial, se utilizan sistemas de producción protegida como alternativa viable para la siembra de Capsicum annuum var. Glabriusculum (Kitta, Katsoulas, Kandila, González-Real, & Baille, 2014); ya que las plantas cultivadas bajo sombra presentan cambios morfológicos como mecanismos de adaptación a la baja disponibilidad de luz. Dicha adaptación incluye cambios en el índice de área foliar, temperatura y humedad relativa, así como mejor distribución del área foliar en altura, lo que puede tener efectos sobre las respuestas productivas y fisiológicas de la planta (Li, Chen, & Li, 2012).
Las mallas fotoselectivas de uso agrícola filtran la radiación solar interceptada (Shahak, Gal, Offir, & Ben-Yakir, 2008), además de estar diseñadas para detectar varias bandas espectrales de la radiación solar y transformar la luz directa a luz difusa. En cultivos como pepino (Cucumis sativus) y pimiento (Capsicum annuum), al utilizar cubiertas de malla blanca, azul, verde y polietileno blanco se incrementó la temperatura, humedad relativa y la evapotranspiración, con lo que se obtuvo mayor crecimiento, rendimiento y contenido de compuestos bioquímicos por planta (Casierra, Matallana, & Zapata, 2014; Hashem, Medany, Abd, & Abdallah, 2011).
Los cambios en la luz espectral promueven diferentes respuestas morfogenéticas y fotosintéticas (Fu, Li, & Wu, 2012). Cada red de plástico modifica la radiación solar que llega al cultivo, lo que reduce el flujo de luz y varía el espectro radiante disponible. Además de la estructura de la red, el espectro de transmitancia también está influenciado por el diámetro del hilo, el color y el grosor de la red, y las propiedades de absorbancia, transmitancia y reflectancia del material plástico (Sica & Picuno, 2008). Con base en lo anterior, este estudio tuvo como objetivo evaluar el efecto de diferentes cubiertas fotoselectivas (mallas e invernadero) sobre variables agronómicas en ecotipos de chile piquín silvestre. Se plantea como hipótesis que el uso de diferentes cubiertas incrementará el rendimiento y los caracteres agronómicos, al menos en uno de los seis ecotipos evaluados.
Materiales y métodos
La investigación se llevó a cabo durante el ciclo agrícola primavera-verano de 2016 en el Departamento de Horticultura de la Universidad Autónoma Agraria Antonio Narro en Saltillo, Coahuila (25° 35’ 63” latitud norte y 101° 03’ 49” longitud oeste, a 1 581 m de altitud). Se utilizaron semillas de chile piquín silvestre recolectadas en los estados de Zacatecas, Nuevo León y Coahuila, México, durante octubre y noviembre de 2015 (Cuadro 1).
Cuadro 1 Colecta y ubicación de ecotipos de chile piquín silvestre.
| Abreviación | Localidad-Estado | Altitud (msnm) | Coordenadas de recolección |
|---|---|---|---|
| RTZ | Río Tuxpan, Zacatecas | 2 060 | 22° 39’ 10.5’’ LN, 102° 42’ 16.2’’ LO |
| PTZ | Puente Tepetatilla, Zacatecas | 1 888 | 22° 47’ 14.1’’ LN, 103° 35’ 11.4’’ LO |
| MZC | Múzquiz, Coahuila | 499 | 28° 00’ 02.9’’ LN, 101° 44’ 45.8’’ LO |
| SAC | San Alberto, Coahuila | 365 | 27° 42’ 83.1’’ LN, 101° 38’ 16.1’’ LO |
| SNL | Santiago, Nuevo León | 480 | 25° 23’ 53.7’’ LN, 100° 06’ 22.8’’ LO |
| LNL | Linares, Nuevo León | 356 | 24° 50’ 14.5’’ LN, 99° 35’ 21.8’’ LO |
Las semillas, tratadas previamente con 500 ppm de ácido giberélico (Biogib®), se sembraron en charolas de poliestireno de 200 cavidades con el fin de romper la testa y obtener mayor germinación. Como sustrato se utilizó turba de sphagnum (Pro Mix®) y perlita (Hortiperl®) en una proporción 2:1 (v/v). Las plantas comenzaron a emerger a los 25 y 38 días después de la siembra. Cuando las plántulas alcanzaron 15 cm de altura, se trasplantaron en bolsas de polietileno con capacidad de 10 L y se colocaron a una distancia de 40 cm entre plantas y 1 m entre surcos (a una densidad de 20 750 plantas·ha-1).
El cultivo se desarrolló en macro túneles (4 m de ancho, 6 m de largo y 2.30 m de alto) con malla “raschel” con 30 % de sombreo y tamaño de orificio de 6 x 8 mm, y un macro túnel cubierto con polietileno blanco lechoso con 20 % de sombra, mientras que la producción en campo abierto tuvo el 100 % de transmisión de luz. Se utilizó la solución nutritiva de Steiner (1976) al 25 % en plántula, al 50 % durante el desarrollo vegetativo, al 75 % en floración y al 100 % en fructificación. El aporte de agua fue de 0.50 a 2.50 L·planta-1·día-1 mediante un sistema de fertirriego por estacas.
Se estableció un diseño experimental con arreglo en parcelas divididas; en donde la parcela grande incluye las cubiertas y la parcela chica los ecotipos (Cuadro 2), lo que dio un total de 36 tratamientos. El análisis de varianza y la comparación de medias de Tukey (P ≤ 0.05) de las variables evaluadas se realizaron con el programa Statistical Analysis System (SAS Institute Inc., 2004). Por su parte, el análisis de componentes principales y las gráficas biplot se llevaron a cabo con la función prcomp en R versión 3.4.3.
Cuadro 2 Descripción de factores evaluados.
| Parcela | Factores | |||||
|---|---|---|---|---|---|---|
| Chica (Ecotipos) | RTZ: Rio Tuxpan, Zacatecas | PTZ: Puente Tepetatilla, Zacatecas | MZC: Múzquiz, Coahuila | SAC: San Alberto, Coahuila | LNL: Linares, Nuevo León | SNL: Santiago, Nuevo León |
| Grande (Cubiertas) | CA: Campo abierto | INV: Invernadero | MN: Malla negra | MR: Malla roja | MA: Malla azul | MB: Malla blanca |
Las variables microclimáticas registradas fueron temperatura ambiental y humedad relativa con un termohigrómetro digital (1452, Taylor®, China), y la radiación fotosintéticamente activa (RFA) se registró con un sensor portátil Quantum (SM-700, Apogee®, EUA). Todas las mediciones se realizaron entre las 7:00 y 19:00 h en el centro de cada macrotunel y en condiciones de cielo despejado. Por su parte, las variables morfológicas evaluadas fueron: altura de planta (AP) y longitud de entrenudos (LE) con cinta métrica, diámetro basal de tallo (DBT) y diámetro de fruto (DF) con un vernier digital (HER-411, Digital Caliper®, China), y rendimiento promedio de fruto (RPF). Esta última se estimó mediante el peso de los frutos por planta con una báscula electrónica (BABOL-100G, Rhino®, China) con capacidad máxima de 100 g y resolución mínima de 0.01 g. Adicionalmente, se cuantificó el número de frutos por planta (NFP), número de semillas (NS), número de hojas (NH) y días a floración (DAF).
Resultados y discusión
Efecto de las mallas en el ambiente del cultivo
La RFA, la temperatura ambiental y la humedad relativa fueron diferentes en cada cubierta con respecto a campo. Solo las mediciones en campo y bajo invernadero superaron los 1 000 μmol·m-2·s-1 (Figura 1), y hubo reducciones entre 43 y 50 % de la RFA con las diferentes cubiertas en comparación con la presentada en campo (Cuadro 3). Los índices máximos de RFA ocurrieron de manera consistente entre las 13:00 y 15:00 h. Con las mallas de colores dicho índice osciló entre 500 y 700 μmol·m-2·s-1, siendo la malla roja la que registró los mayores valores y la malla azul los menores. La RFA por encima de 1 000 μmol·m-2·s-1 tiene un efecto directo en la tasa de fotosíntesis, lo cual afecta negativamente la productividad, el crecimiento y el rendimiento (Kitta et al., 2014). Los resultados de la disminución de la RFA fueron notables debido a que la malla sombra provocó un efecto de amortiguamiento en los cambios de este parámetro en el transcurso del día (Figura 1).

Figura 1 Comportamiento de la radiación fotosintéticamente activa (RFA) bajo las diferentes cubiertas en comparación con el campo abierto.
Cuadro 3 Comparación de medias de la radiación y temperatura registradas durante el ciclo de producción de chile piquín bajo los diferentes ambientes.
| Ambiente | Reducción de RFA1 respecto a CA (%) | Temperatura (°C) | HR (%) |
|---|---|---|---|
| Malla roja | 43.59 | 31.23 bz | 31.84 c |
| Malla azul | 50.10 | 30.18 d | 33.36 a |
| Malla blanca | 49.14 | 30.01 d | 32.83 b |
| Malla negra | 48.86 | 29.07 e | 32.96 b |
| Invernadero | 7.16 | 31.68 b | 31.63 d |
| Campo abierto | 0 | 33.94 a | 32.72 b |
1RFA = radiación fotosintéticamente activa; CA = campo abierto; HR = humedad relativa. zMedias con la misma letra dentro de cada columna no difieren estadísticamente (Tukey, P ≤ 0.05)
De acuerdo con Retamales, Montecino, Lobos, y Rojas (2008), en el cultivo de arándano se ha observado que la RFA, transmitida por mallas de color blanco y rojo con 35 % de sombra, y con malla gris con 50 % de sombra, se redujo hasta un 29 %; mientras que la malla de color rojo con 50 % de sombra y la malla negra con 35 % de sombra disminuyeron la RFA en 41 y 47 %, respectivamente, en comparación con el campo. Por ello, el uso de malla sombra resulta relevante para reducir el impacto de la alta intensidad de luz en condiciones de campo abierto y transformar la radiación directa en dispersa. Lo anterior permite que la luz penetre en el dosel interno de la planta, lo que previene quemaduras y da un efecto moderado de enfriamiento (Ilic et al., 2017).
Shahak (2014) señala que con cualquier cubierta que se coloque sobre las plantas, además de los efectos sobre la intensidad y la calidad de la luz, se pueden modificar los elementos climáticos (radiación solar, velocidad del viento, temperatura y humedad relativa) (Arthurs, Stamps, & Giglia, 2013), alterando el metabolismo y el consumo de agua de las plantas, lo que repercute positivamente en el rendimiento y la calidad del fruto (Tanny, 2012). La temperatura y la humedad relativa juegan un papel importante en la conductancia estomática foliar y, por ende, en la tasa de transpiración y fotosíntesis de la planta (Righi, Buriol, Angelocci, Heldwein, & Tazzo, 2012).
Efecto de las mallas sobre el cultivo
Se encontraron efectos altamente significativos (P ≤ 0.01) entre cubiertas en todas las variables de crecimiento estudiadas (Cuadro 4). Las plantas cultivadas bajo malla blanca presentaron la mayor diferencia estadística en siete de las nueve variables estudiadas (AP, DBT, RPF, NFP, DF, NS y NH), y las plantas cultivadas bajo malla azul tuvieron los mejores valores en cuatro de las variables (DAF, DF, NS y LE); por lo que esta última se puede considerar como la segunda mejor cubierta para cultivo de chile piquín (Cuadro 5).
Cuadro 4 Cuadrados medios del análisis de varianza de las variables de los diferentes ecotipos de chile piquín desarrollados bajo diferentes cubiertas.
| Fuente de variación | GL1 | AP | DBT | DAF | NFP | RPF | DF | NS | LE | NH |
|---|---|---|---|---|---|---|---|---|---|---|
| Cubierta | 5 | 1 192.4** | 12.3** | 1 001.7** | 720.6** | 103.8** | 154.5** | 548.4** | 432.6** | 162 629.7** |
| Ecotipo | 5 | 1 333.4** | 14.9** | 633.0** | 5 328.7** | 312.4** | 58.0** | 125.2** | 40.0** | 23 717.3** |
| Cubierta x ecotipo | 25 | 104.1** | 2.1** | 68.2** | 383.0* | 84.3** | 4.4** | 6.1** | 4.1** | 5 035.7** |
| CV (%) | 5.29 | 3.08 | 2.75 | 8.50 | 16.94 | 1.87 | 7.55 | 8.15 | 6.42 |
1GL = grados de libertad; AP = altura de planta; DBT = diámetro basal de tallo; DAF = días a floración; NFP = número de frutos por planta; RPF = rendimiento promedio de fruto; DF = diámetro de fruto; NS = número de semillas; LE = longitud de entrenudos; NH = número de hojas; CV = coeficiente de variación. * = significativo (P ≤ 0.05); ** = altamente significativo (P ≤ 0.01).
Cuadro 5 Comparación de medias de las variables agronómicas de los diferentes ecotipos de chile piquín y ambientes evaluados.
| Factores | Variables evaluadas | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ecotipo | AP1 (cm) | DBT (mm) | DAF (días) | RPF (g) | NFP | DF (mm) | NS | LE (cm) | NH |
| RTZ | 36.95 az | 6.15 a | 43.66 a | 9.29 a | 40.04 a | 11.11 a | 14.79 a | 12.45 a | 193.58 c |
| PTZ | 34.70 b | 6.01 b | 53.54 b | 0.98 b | 8.79 b | 6.46 b | 11.50 c | 9.75 c | 152.16 e |
| LNL | 25.75 c | 5.11 c | 58.62 d | 0.16 b | 1.95 d | 6.29 b | 10.79 c | 9.66 cd | 219.95 a |
| MZC | 23.54 d | 4.92 d | 54.12 b | 0.51 b | 4.62 d | 8.30 ab | 14.66 ab | 10.91 b | 206.66 b |
| SNL | 23.25 d | 4.71 e | 56.29 c | 0.35 b | 2.79 d | 7.02 b | 12.04 bc | 9.25 cd | 166.50 d |
| SAC | 17.12 e | 4.09 f | 54.37 b | 0.26 b | 2.20 d | 6.60 b | 7.91 d | 9.04 d | 141.37 f |
| Cubiertas | |||||||||
| MB | 36.41 a | 6.29 a | 47.29 b | 5.68 a | 19.33 a | 10.77 a | 18.87 a | 13.66 b | 303.08 a |
| MA | 32.58 b | 5.39 b | 45.12 a | 3.10 b | 13.95 b | 12.17 a | 17.66 a | 15.67 a | 175.12 c |
| IN | 27.83 c | 5.37 b | 55.95 d | 0.86 b | 8.58 c | 6.02 b | 7.37 cd | 6.62 e | 139.12 d |
| MR | 24.45 d | 4.76 d | 55.62 d | 0.70 c | 6.50 d | 6.86 b | 12.83 b | 7.87 d | 248.79 b |
| MN | 23.04 e | 4.15 e | 53.66 c | 0.69 c | 6.87 cd | 5.39 b | 8.95 c | 12.16 c | 135.91 d |
| CA | 17.00 f | 5.03 c | 62.95 e | 0.44 c | 5.16 d | 4.56 b | 6.00 d | 5.08 f | 78.20 e |
| ** | ** | ** | ** | ** | ** | * | ** | ** | |
| CV (%) | 5.29 | 3.08 | 2.75 | 16.94 | 8.5 | 1.87 | 7.55 | 8.15 | 6.42 |
1AP = altura de planta; DBT = diámetro basal de tallo; DAF = días a floración; RPF = rendimiento promedio de fruto; NFP = número de frutos por planta; DF = diámetro de fruto; NS = número de semillas; LE = longitud de entrenudos; NH = número de hojas; RTZ = Rio Tuxpan, Zacatecas; PTZ = Puente Tepetatilla, Zacatecas; LNL= Linares, Nuevo León; MZC = Múzquiz, Coahuila; SNL = Santiago, Nuevo León; SAC = San Alberto, Coahuila; MB = malla blanca; MA = malla azul; IN = invernadero; MR = malla roja; MN = malla negra; CA = campo abierto; CV = coeficiente de variación. ** = altamente significativo (P ≤ 0.01). zMedias con la misma letra dentro de cada columna no difieren estadísticamente (Tukey, P ≤ 0.05).
Por su parte, las plantas cultivadas en campo abierto mostraron los peores resultados en ocho de las nueve variables estudiadas (excepto en DBT) (Cuadro 5). Asimismo, las plantas cultivadas bajo malla de color negro mostraron los valores más bajos en cuatro de las variables (DBT, RPF, NFP y DF), lo cual soporta la necesidad de buscar las mejores condiciones para el desarrollo de chile piquín silvestre; en este caso mediante el uso de mallas que proporcionen los rangos óptimos en las variables microclimáticas.
En la malla blanca se produjo un mayor NFP y RPF (374.61 y 1 290.90 %, respetivamente), en comparación con la producción en campo abierto (Cuadro 5). Igualmente, los frutos cultivados bajo malla blanca y azul produjeron los mayores DF (236.18 y 266.88 %, respectivamente) y NS (314.50 y 294.33 %, respectivamente), ambos con respecto a la producción en campo abierto.
La disminución de la disponibilidad de la RFA puede inducir respuestas morfogenéticas como incremento del área foliar, y alargamiento del vástago y entrenudos, debido a que las células se expanden más con bajas intensidades de radiación solar para captar la luz y llevar a cabo la fotosíntesis. Lo anterior está asociado con el fenómeno conocido como “evitación de la sombra”, ya que está influenciado por la actividad fitocromática que regula el alargamiento del tallo, al transformar plastidios no fotosintéticos (etioplastos) en cloroplastos completamente desarrollados con función fotosintética, y el área foliar, con los consiguientes efectos sobre el reparto de fotoasimilación y el crecimiento del fruto (Bastías, Manfrini, & Grappadelli, 2012).
En este estudio, las mallas e invernadero propiciaron un incremento en la altura final de las plantas de chile piquín (Cuadro 5), probablemente en respuesta a la reducción de la luz (Salisbury & Ross, 2000). Se ha reportado que al usar mallas cambia la AP dependiendo del color de la malla; en pimiento morrón la mayor AP se obtuvo con mallas azules (Ayala-Tafoya et al., 2015) y en albahaca con malla negra (Martínez-Gutiérrez, Nicolás-Santana, Ortiz-Hernández, Morales, & Gutiérrez-Hernández, 2016).
La radiación es uno de los factores capaces de producir mayor actividad fotosintética en las plantas; además, la calidad y la distribución del espectro de luz pueden influir en la longitud del entrenudo y el desarrollo de la floración. (Runkle & Heins, 2006). En este caso, dicho efecto sobresalió en las mallas blanca y azul, donde se registró un aumento en la AP de 114.05 y 91.53 %, respectivamente, en comparación con campo abierto. Adicionalmente, la malla blanca incrementó 34.1 % el DBT en contraste con la malla negra, donde se obtuvo el menor resultado. Esto coincide con lo reportado por Ayala-Tafoya et al. (2011), quienes mencionan que la malla sombra de color aumentó el DBT en el cultivo de tomate.
El análisis de componentes principales (ACP) reveló una agrupación de las plantas en el segundo cuadrante (Figura 2), lo cual indica que éstas mostraron menor AP, NFP y DF, y mayor DAF (CP1), además de un mayor DBT y peso del fruto, y menor NS (CP2); lo anterior considerando la relevancia de esas variables para cada uno de los componentes (Cuadro 6). Sin embargo, en general, los materiales se comportaron de modo disperso en las condiciones analizadas, lo cual revela que el ecotipo jugó un papel determinante en las variables estudiadas. Es importante resaltar que un mayor DBT permite a la planta tener mejor desarrollo de su sistema vascular, mediante el cual conduce el agua y los nutrimentos, lo que mejora los procesos fisiológicos (Bahena-Delgado, Bustos-Rangel, Broa-Rojas, & Jaime-Hernández, 2012).
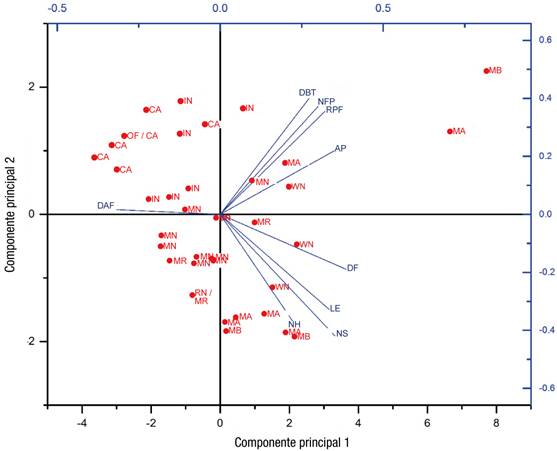
Figura 2 Gráfica de componentes principales de las variables analizadas y tipo de cubierta. AP = altura de planta; DBT = diámetro basal de tallo; DAF = días a floración; RPF = rendimiento promedio de fruto; NFP = número de frutos por planta; DF = diámetro de fruto; NS = número de semillas; LE = longitud de entrenudos; NH = número de hojas; MB = malla blanca; MA = malla azul; IN = invernadero; MR = malla roja; MN = malla negra; CA = campo abierto.
Cuadro 6 Vectores característicos y proporción de la varianza explicada por los dos primeros componentes principales.
| Variable evaluada | CP11 | CP2 |
|---|---|---|
| Altura de planta | 0.33247 | 0.15243 |
| Diámetro basal de tallo | 0.26704 | 0.33062 |
| Días a floración | -0.35263 | 0.05962 |
| Número de frutos por planta | 0.32774 | 0.36468 |
| Rendimiento promedio de fruto | 0.31251 | 0.30546 |
| Diámetro de fruto | 0.35866 | -0.22458 |
| Número de semillas | 0.31634 | -0.45157 |
| Longitud de entrenudos | 0.30149 | -0.36418 |
| Número de hojas | 0.22385 | -0.4196 |
| Varianza explicada | 63.33 | 15.15 |
| Varianza acumulada | 63.33 | 78.48 |
1CP1 y CP2 = coeficientes de los componentes principales 1 y 2.
Díaz-Pérez (2014) encontró un aumento en el número y el tamaño de frutos en pimiento morrón bajo condiciones de sombreo. Shahak et al. (2008) al evaluar mallas de colores “raschel” (rojo, amarillo y perla, con 30 a 40 % de sombra) en pimiento obtuvieron un RPF de 115 a 135 % más en comparación con los resultados obtenidos con malla negra y en campo abierto. En contraste, Ayala-Tafoya et al. (2015) no encontraron diferencias significativas en el NFP en pimiento, esto al utilizar diferentes mallas de colores y producción sin malla.
Efecto de los ecotipos
Todas las variables de crecimiento mostraron diferencias altamente significativas (P ≤ 0.01) entre los ecotipos analizados (Cuadro 4). El ecotipo RTZ tuvo el mejor desempeño en ocho de las nueve variables (AP, DBT, DAF, RPF, NFP, DF, NS y LE), mientras que SAC tuvo el peor desempeño en casi todas las variables. La comunidad de procedencia de este último se encuentra a 365 m de altitud, la cual es mucho menor que la del lugar donde se estableció el experimento (1 580 m de altitud); en tanto que el lugar de procedencia del ecotipo RTZ está a 2 060 m de altitud, lo que pudo influir en la adaptación a las condiciones del experimento. No obstante, esto no coincide con lo reportado por Martínez-Sánchez, Pérez-Grajales, Rodríguez-Pérez, y Moreno-Pérez (2010), quienes recolectaron especies silvestres de Capsicum annuum en altitudes de 940 a 1 600 m, y las evaluaron a 2 247 m sin encontrar diferencias estadísticas significativas entre especies en las variables AP, DBT, NFP y tamaño de estos.
El ACP no reveló agrupaciones concretas de los ecotipos, lo cual sustenta la idea de que el ambiente también juega un papel relevante en la dispersión de los datos; sin embargo, se observaron algunas tendencias, por ejemplo, el ecotipo RTZ se agrupó en mayor medida en el cuadrante 1 (Figura 3), lo cual indica que esas plantas tuvieron un mejor desempeño en cuanto a DF, NFP, AP y DBT. Además, este ecotipo presentó mayor AP (46.5 %) y DBT (34.5 %) en comparación con el ecotipo SAC (Cuadro 5), y exhibió mayor precocidad (DAF), 14 días antes que el ecotipo más tardío (LNL), y 95 % más frutos por planta en comparación con el mismo ecotipo (Cuadro 5).
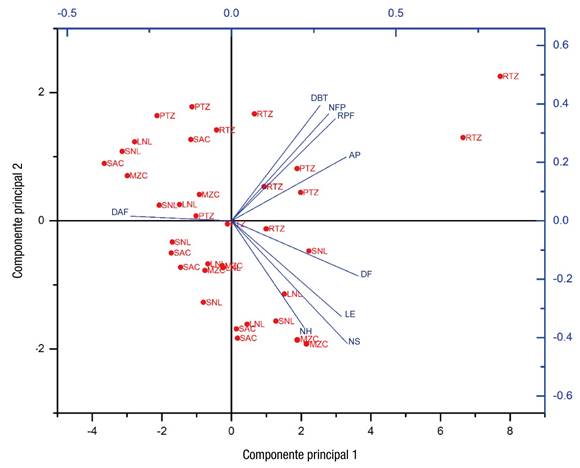
Figura 3 Gráfica de componentes principales de las variables analizadas y ecotipos. AP = altura de planta; DBT = diámetro basal de tallo; DAF = días a floración; RPF = rendimiento promedio de fruto; NFP = número de frutos por planta; DF = diámetro de fruto; NS = número de semillas; LE = longitud de entrenudos; NH = número de hojas; RTZ = Rio Tuxpan, Zacatecas; PTZ = Puente Tepetatilla, Zacatecas; LNL= Linares, Nuevo León; MZC = Múzquiz, Coahuila; SNL = Santiago, Nuevo León; SAC = San Alberto, Coahuila.
Por otro lado, la malla azul aumentó la precocidad a la floración en casi 18 días con respecto al campo abierto, además de ser la mejor cubierta en cuanto a LE con 62.81 % más que la producción sin cubierta. En esta misma variable, el ecotipo RTZ fue mayor (27.38 %) al ecotipo SAC (Cuadro 5).
El rendimiento, tamaño y calidad en Capsicum, provocados por el efecto de sombreado, dependen en gran medida del área geográfica y la tecnología de producción (Zhu, Peng, Liang, Wu, & Hao, 2012). En general, los ecotipos RTZ y MZC presentaron el mayor DF (40.59 y 20.48 %, respectivamente) y NS (46.52 y 46.04 %, respectivamente), en comparación al ecotipo SAC (Cuadro 5). Salinas-Hernández, Liévano-Liévano, Ulín-Montejo, Mercado, y Petit-Jiménez (2010) también obtuvieron mayor longitud, DF y NS en dos tipos de chile amashito silvestre (“Pico de paloma” y “Garbanzo”), al compararlos con los demás tipos de chile evaluados.
Santos y Salame-Donoso (2012) encontraron que el sombreo al 35 % sobre el cultivo de arándano (Vaccinium oxycoccus) incrementa la floración, el peso del fruto y el rendimiento, en contraste con la producción en cielo abierto. Por otro lado, Nooryazdan, Serieys, Baciliéri, David, y Bervillé (2010) reportan que las diferencias en características morfológicas, al evaluar 77 poblaciones de Helianthus annuus silvestre, colectadas en Estados Unidos, se correlacionaron con las variables climáticas de los sitios de origen de las poblaciones, lo cual sugiere adaptación a las condiciones locales de las especies evaluadas.
Efecto de la interacción
El análisis de varianza de dos vías reveló que la interacción entre las cubiertas utilizadas y los ecotipos presenta un efecto significativo en todas las variables. Lo anterior coincide con lo mencionado por Hernández-Verdugo et al. (2015), quienes reportan diferencias significativas en todas las características fenotípicas evaluadas en chile silvestre bajo diferentes niveles de sombra. En términos generales, se puede señalar que el uso de malla sombra blanca proporciona mejores resultados en las variables analizadas en todos los ecotipos (Cuadro 5), y que el ecotipo RTZ presenta los mejores valores en todas las cubiertas utilizadas.
El análisis de los vectores reveló que cierto grupo de variables presentaron alta correlación entre sí (DBT, RPF, AP y NFP) (Figuras 2 y 3). Estos resultados son similares a los reportados por Meena-Prakash y Bahadur (2015), en AP y NFP, y por Ogwulumba y Ugwuoke (2013), para NH y NFP. Por su parte, las variables DF, LE, NS y NH formaron otro grupo, el cual presentó poca correlación con el grupo anterior.
Meena-Prakash y Bahadur (2015) indican que la AP mostró una correlación negativa significativa con respecto al peso del fruto y el diámetro polar de la fruta, lo que revela que a mayor AP, el peso y el DF disminuyen; mientras que la variable DAF presentó una correlación negativa con todas las variables, lo que indica que entre mayor sea el número de DAF, el desempeño en las otras variables es menor. En este estudio, el análisis biplot reveló que todas las variables, excepto DAF, están relacionadas positivamente con los cultivos bajo malla blanca o azul (Figura 2); es decir, los cultivos bajo esas cubiertas tuvieron los mejores resultados en casi todas las variables. En cuanto a los ecotipos, el RTZ se relacionó de modo positivo con las variables DBT, RPF, AP y NFP (Figura 3).
La variable DAF en cielo abierto e invernadero (Figura 2), revela que las plantas cultivadas bajo estas condiciones tardan más en presentar flores y tienen un desempeño menor en el resto de las variables, lo que se observó en todos los ecotipos cuando se cultivaron a cielo abierto (Figura 3).
Conclusiones
El uso de cubiertas redujo la RFA, lo que tuvo un efecto positivo en la morfología de las plantas. Las mallas de color blanco y azul incrementaron el rendimiento, el número de frutos y el tamaño de estos, y el ecotipo RTZ fue el que presentó mejores resultados en todas las variables estudiadas.
La combinación del ecotipo RTZ con malla de color blanco mejoró los caracteres agronómicos y el rendimiento en chile piquín, por lo que se considera una alternativa viable para la producción de Capsicum annuum var. Glabriusculum, bajo condiciones similares a las de este estudio.
Agradecimientos
Los autores agradecen a la Universidad Autónoma Agraria Antonio Narro (UAAAN), al Consejo Nacional de Ciencia y Tecnología (CONACYT) y a la Secretaría de Educación Pública (SEP) a través del Programa para el Desarrollo Profesional Docente (PRODEP) por su apoyo brindado en el financiamiento de este proyecto.
REFERENCIAS
Arthurs, S. P., Stamps, R. H., & Giglia, F. F., (2013). Environmental modification inside photoselective shade houses. HortScience, 48(8), 975-979. Retrieved from http://hortsci.ashspublications.org/content/48/8/975.full.pdf [ Links ]
Ayala-Tafoya, F., Sánchez-Madrid, R., Partida-Ruvalcaba, L., Yáñez-Juárez, M., Ruiz-Espinosa, F., Velázquez Alcaraz, T., .& Parra-Delgado, J. (2015). Producción de pimiento morrón con mallas sombra de colores. Revista Fitotecnia Mexicana, 38(1), 93-99. Retrieved from http://www.redalyc.org/articulo.oa?id=61035375012 [ Links ]
Ayala-Tafoya, F., Zatarain-López, D., Valenzuela-López, M., Partida-Ruvalcaba, L., Velázquez-Alcaraz, T., Díaz-Valdés, T., & Osuna-Sánchez, J. (2011). Crecimiento y rendimiento de tomate en respuesta a radiación solar transmitida por mallas sombra. Terra Latinoamericana, 29(4), 403-410. Retrieved from http://www.redalyc.org/articulo.oa?id=57322342006 [ Links ]
Bahena-Delgado, G., Bustos-Rangel, A. J., Broa-Rojas, E., & Jaime-Hernández, M. A. (2012). Agronomic behavior of creole chili (Capsicum annuum L.) in fertirrigation with plastic mulch and row cover in Xalostoc, Morelos. Ingeniería Agrícola y Biosistemas, 4(1), 19-24. doi: 10.5154/r.inagbi.2011.11.11014 [ Links ]
Bastías, R. M., Manfrini, L., & Grappadelli, L. C. (2012). Exploring the potential use of the photo-selective nets for fruit growth regulation in apple. Chilean Journal of Agricultural Research, 72(2), 224-231. Retrieved from http://www.bioline.org.br/pdf?cj12036 [ Links ]
Casierra, P. F., Matallana, D., & Zapata, E. (2014). Growth of bell pepper plants (Capsicum annuum) affected by coloured covers. Healthy plants, 66(4), 149-155. doi: 10.1007/s1034 [ Links ]
Castronuovo, D., Statuto, D., Muro, N., Picuno, P., & Candido, V. (2017). The use of shading nets for the greenhouse cultivation of sweet pepper in the Mediterranean area. Acta Horticulturae 1170, 373-380. doi: 10.17660/ActaHortic.2017.1170.46 [ Links ]
Conde-Álvarez, C., & Saldaña-Zorrilla, S. O. (2007). Cambio climático en América Latina y el Caribe: Impactos, vulnerabilidad y adaptación. Ambiente y Desarrollo, 23(2), 23-30. Retrieved from http://www.keneamazon.net/Documents/Publications/Virtual-Library/Economia-Desarrollo/39.pdf [ Links ]
Díaz-Pérez, J. C., (2014). Bell pepper (Capsicum annum L.) crop as affected by shade level: Fruit yield, quality, and postharvest attributes and incidence of Phytophthora blight (caused by Phytophthora capsici Leon.). HortScience , 49(7), 891-900. Retrieved from http://hortsci.ashspublications.org/content/49/7/891.full.pdf [ Links ]
Fu, W., Li, P., & Wu, Y. (2012). Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Scientia Horticulturae, 135, 45-51. doi: 10.1016/j.scienta.2011.12.004 [ Links ]
Hashem, F. A., Medany, M. A., Abd, E. M., & Abdallah, M. M. F. (2011). Influence of green-house cover on potential evapotranspiration and cucumber water requirements. Annals of Agricultural Science, 56(1), 49-55. doi: 10.1016/j.aoas.2011.05.001 [ Links ]
Hernández-Verdugo, S., González-Sánchez, R. A., Porras, F., Parra-Terraza, S., Valdez-Ortiz, A., Pacheco-Olvera, A., & López-España, R. G. (2015). Phenotypic plasticity of populations of wild chile (Capsicum annuum var. glabriusculum) in response to light availability. Botanical Sciences, 93(2), 231-240. doi: 10.17129/botsci.237 [ Links ]
Ilic, S. Z., Milenkovic, L., Dimitrijevic, A., Stanojevic, L., Cvetkovic, D., Kevresan, Z., Fallik, E., & Mastilovic, J. (2017). Light modification by color nets improve quality of lettuce from summer production. Scientia Horticulturae , 226, 389-397. doi: 10.1016/j.scienta.2017.09.009 [ Links ]
Kitta, E., Katsoulas, N., Kandila, A., González-Real, M. M., & Baille, A. (2014). Photosynthetic acclimation of sweet pepper plants to screenhouse conditions. HortScience , 49(2), 166-172. Retrieved from http://hortsci.ashspublications.org/content/49/2/166.full.pdf [ Links ]
Li, X., Chen, W., & Li, Y. (2012). Study on photosynthetic characteristics of blueberry in greenhouse. Acta Horticulturae, 926, 315-320. doi: 10.17660/ActaHortic.2012.926.43 [ Links ]
Martínez-Gutiérrez, G. A., Nicolás-Santana, L., Ortiz-Hernández, Y. D., Morales, I., & Gutiérrez-Hernández, G. F. (2016). Crecimiento y contenido de aceite en albahaca (Ocimum basilicum L.) bajo mallas sombra de colores. Interciencia, 41(6), 428-432. Retrieved from http://www.redalyc.org/articulo.oa?id=33945816010 [ Links ]
Martínez-Sánchez, D., Pérez-Grajales, M., Rodríguez-Pérez, J. E., & Moreno-Pérez, E. C. (2010). Colecta y caracterización morfológica de ‘chile de agua’ (Capsicum annuum L.) en Oaxaca, México. Revista Chapingo Serie horticultura, 16(3), 169-176. doi: 10.5154/r.rchsh.2010.16.021 [ Links ]
Meena-Prakash, O., & Bahadur, V. (2015). Genetic associations analysis for fruit yield and its contributing traits of indeterminate tomato (Solanum lycopersicum L.) germplasm under open field condition. Journal of Agricultural Science, 7(3), 148-164. doi: 10.5539/jas.v7n3p148 [ Links ]
Nooryazdan, H., Serieys, H., Baciliéri, R., David, J., & Bervillé, A. (2010). Structure of wild annual sunflower (Helianthus annus L.) accessions based on agro-morphological traits. Genetic Resources and Crop Evolution, 57(1), 27-39. doi: 10.1007/s10722-009-9448-9 [ Links ]
Ogwulumba, S. I., & Ugwuoke, K. I. (2013). Coefficient and path analyses of the impact of root galls caused by Meloidogyne javanica on some growth and yield parameters of tomato (Solanum lycopersicum). International Journal of Plant and Soil Science, 2(2), 222-229. doi: 10.9734/IJPSS/2013/3510 [ Links ]
Retamales, J. B., Montecino, J. M., Lobos, G. A., & Rojas, L. A. (2008). Colored shading nets increase yields and profitability of highbush blueberries. Acta horticulturae, 770, 193-197. doi: 10.17660/ActaHortic.2008.770.22 [ Links ]
Righi, E. Z., Buriol, G. A., Angelocci, L. R., Heldwein, A. B., & Tazzo, I. F. (2012). Relationships of photosynthetic photon flux density, air temperature and humidity with tomato leaf diffusive conductance and temperature. Brazilian Archives of Biology and Technology, 55(3), 359-370. doi: 10.1590/S1516-8913201200030000 [ Links ]
Rodríguez-del Bosque, L. A., Pozo-Campodónico, O., Ramírez-Meraz, M., Silva-Cavazos, F. J., Zúñiga-Estrada, R., Sánchez-de la Cruz, R., Medina-Martínez, T., & Villalón-Mendoza, H. (2003). Efecto del sombreado sobre el desarrollo y rendimiento de 10 colectas de chile piquín (Capsicum annuum var. aviculare) en cuatro localidades del noreste de México. México: Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Retrieved from http://biblioteca.inifap.gob.mx:8080/jspui/bitstream/handle/123456789/3482/1773%201er%20Simposio%20de%20Chile%20Piqu%C3%ADn%2C%20Memoria.pdf?sequence=1 [ Links ]
Rueda-Puente, E. O., Murillo-Amador, B., Castellanos-Cervantes, T., García-Hernández, J. L., Tarazòn-Herrera, M. A., Moreno-Medina, S., & Gerlach-Barrera, L. E. (2010). Effects of plant growth promoting bacteria and mycorrhizal on Capsicum annuum L. var. aviculare ([Dierbach] D'Arcy and Eshbaugh) germination under stressing abiotic conditions. Plant Physiology and Biochemistry, 48(8), 724-730. doi: 10.1016/j.plaphy.2010.04.002 [ Links ]
Runkle, E. S., & Heins, R. D. (2006). Manipulating the light environment to control flowering and morphogenesis of herbaceous plants. Acta Horticulturae , 771, 51-60. doi: 10.17660/ActaHortic.2006.711.4 [ Links ]
Salinas-Hernández, R., Liévano-Liévano, E., Ulín-Montejo, F., Mercado, J., & Petit-Jiménez, D. (2010). Caracterización morfológica y cambios durante la vida postcosecha de cuatro tipos de chile amashito (Capsicum annuum L.) variedad glabriusculum (Dunal) Heiser & Pickersgill. Revista Iberoamericana de Tecnología Postcosecha, 11(1), 92-100. Retrieved from http://www.redalyc.org/articulo.oa?id=81315093012 [ Links ]
Salisbury, F., & Ross, F. (2000). Fisiología de las plantas: Desarrollo de las plantas y fisiología ambiental. Spain: S.A. Ediciones Paraninfo. [ Links ]
Santos, B. M., & Salame-Donoso, T. P. (2012). Performance of southern highbush blueberry cultivars under high tunnels in Florida. HortTechnology, 22(5), 700-704. doi: 10.21273/HORTTECH.22.5.700 [ Links ]
Shahak, Y., Gal, E., Offir, Y., & Ben-Yakir, D. (2008). Photoselective shade netting integrated with greenhouse technologies for improved performance of vegetable and ornamental crops. Acta Horticulturae , 797, 75-80. doi: 10.17660/ActaHortic.2008.797.8 [ Links ]
Shahak, Y. (2014). Photoselective netting: An overview of the concept, R & D and practical implementation in agriculture. Acta Horticulturae , 1015, 155-162. doi: 10.17660/ActaHortic.2014.1015.17 [ Links ]
Sica, C., & Picuno, P. (2008). Spectro-radiometrical characterization of plastic nets for protected cultivation. Acta Horticulturae , 801, 245-252 doi: 10.17660/ActaHortic.2008.801.23 [ Links ]
Statistical Analysis System (SAS Institute Inc.). (2004). Software product support manual , ver. 9.2. Cory, N. C.: Author. [ Links ]
Steiner, A. A. (1973). The selective capacity of tomato plants for ions in a nutrient solution. In: Proceedings 3rd International Congress on Soiles Culture. [ Links ]
Tanny, J. (2012). Microclimate and evapotranspiration of crops covered by agricultural screens: A review. Biosystems Engineering, 114(1), 26-43. doi: 10.1016/j.biosystemseng.2012.10.008 [ Links ]
Villalón-Mendoza, H., Medina-Martinez, T., Ramírez-Meraz, M., Solis-Urbina, S. E., & Maiti, R. (2014). Factors influencing the price of chile piquin wild chili (Capsicum annuum L. var. Glabriusculum) of North-east Mexico. International Journal of Bio-resource and Stress Management, 5(1), 128-131. doi: 10.5958/j.0976-4038.5.1.025 [ Links ]
Zhu, J. J., Peng, Q., Liang, Y. L., Wu, X., & Hao, W. L. (2012). Leaf gas exchange, chlorophyll fluorescence, and fruit yield in hot pepper (Capsicum annuum L.) grown under different shade and soil moisture during the fruit growth stage. Journal of Integrative Agriculture 11(6), 927-937. doi: 10.1016/S2095-3119(12)60083-5 [ Links ]
Recibido: 27 de Mayo de 2018; Aprobado: 13 de Diciembre de 2018
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
