










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.28 no.3 Chapingo sep./dic. 2022 Epub 08-Mar-2024
https://doi.org/10.5154/r.rchscfa.2021.05.030
Artículo científico
Nueva ecuación dinámica de altura dominante e índice de sitio para Pinus chiapensis (Martínez) Andresen en Puebla y Veracruz, México
1Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Centro de Investigación Regional Golfo Centro" icountry="MX">Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Centro de Investigación Regional Golfo Centro" icountry="MX">Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Centro de Investigación Regional Golfo Centro, Campo Experimental San Martinito. Carretera Federal México-Puebla km 56.5. C. P. 74100. Tlahuapan, Puebla, México.
2Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Centro de Investigación Regional Golfo Centro, Campo Experimental Ixtacuaco. Carretera Martínez de la Torre-Tlapacoyan km 4.5. C. P. 95600. Tlapacoyan, Veracruz, México.
3Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias, Centro de Investigación Regional Norte Centro, Campo Experimental Valle del Guadiana. Carretera Durango-Mezquital km 4.5. C. P. 34170. Durango, Durango, México.
4Universidad de la Sierra Juárez, Instituto de Estudios Ambientales, División de Estudios de Postgrado. Av. Universidad s/n. C. P. 68725. Ixtlán de Juárez, Oaxaca, México.
Introducción:
Pinus chiapensis (Martínez) Andresen carece en muchas regiones de modelos de índice de sitio (IS) para clasificar la productividad maderable.
Objetivo:
Desarrollar una ecuación dinámica de IS, mediante el enfoque de diferencia algebraica generalizada (DAG), para describir el patrón de crecimiento en altura dominante y clasificar la productividad de rodales naturales de P. chiapensis en Puebla y Veracruz, México.
Materiales y métodos:
Cuatro modelos teóricos de crecimiento se utilizaron para derivar seis ecuaciones en DAG, ajustadas a observaciones de altura dominante-edad procedentes de análisis troncales de 31 árboles. El ajuste se realizó con el método de variables Dummy que es invariante de la edad base; se corrigió la autocorrelación y la heterocedasticidad.
Resultados y discusión:
La evaluación cuantitativa, el análisis gráfico de los residuales y de las tendencias de crecimiento de las ecuaciones permitió la selección de una ecuación derivada del modelo de Levakovic II con capacidad predictiva mayor. Con esta ecuación y una edad base de 50 años se construyeron curvas de IS polimórficas con asíntotas variables para clasificar la productividad en baja, media y alta, correspondientes a IS de 25, 32 y 39 m, respectivamente. El incremento medio anual máximo para el IS de 32 m fue de 1.07 m∙año-1 y ocurrió a los 11.08 años. La ecuación exhibió mejor desempeño con respecto a una ecuación polimórfica previamente reportada.
Conclusiones:
Se recomienda usar la ecuación desarrollada para predecir el crecimiento en altura dominante e IS de rodales de P. chiapensis en Puebla y Veracruz, México.
Palabras clave: diferencia algebraica generalizada; crecimiento en altura; invarianza de edad base; método de variables Dummy; modelo de Levakovic
Introduction:
Pinus chiapensis (Martínez) Andresen lacks in many areas of site index (SI) models to classify timber productivity.
Objective:
To develop a dynamic SI equation, using the generalized algebraic difference approach (GADA) to describe the dominant height growth pattern and classify the productivity of natural stands of P. chiapensis in Puebla and Veracruz, Mexico.
Materials and methods:
Four theoretical growth models were used to derive six equations in GADA, fitted to dominant height-age observations from stem analyses of 31 trees. Fitting was performed using the Dummy variable method, which is invariant to the base age; autocorrelation and heteroscedasticity were corrected.
Results and discussion:
Quantitative evaluation, graphical analysis of residuals and growth trends of equations allowed the selection of an equation derived from the Levakovic II model with higher predictive capacity. With this equation and a base age of 50 years, polymorphic SI curves with variable asymptotes were constructed to classify productivity into low, medium and high, corresponding to SI of 25, 32 and 39 m, respectively. The maximum mean annual increase for the SI of 32 m was 1.07 m∙yr-1 and occurred at 11.08 years. The equation exhibited better performance relative to a previously reported polymorphic equation.
Conclusions:
It is recommended to use the developed equation to predict dominant height growth and SI of P. chiapensis stands in Puebla and Veracruz, Mexico.
Keywords: generalized algebraic difference approach; height growth; base age invariance; Dummy variable method; Levakovic model
Ideas destacadas:
El crecimiento de P. chiapensis se estimó mediante el enfoque de diferencias algebraicas generalizadas.
Una ecuación de Levakovic II tuvo capacidad predictiva mayor de altura dominante e índice de sitio.
La ecuación dinámica genera polimorfismo y asíntotas múltiples de manera concurrente.
Se construyeron curvas de índice de sitio para 25, 32 y 39 m con edad base de 50 años.
La ecuación exhibió mejor desempeño con respecto a una ecuación polimórfica previamente reportada.
Introducción
La estimación del potencial productivo de sitios forestales es importante para el manejo forestal porque ayuda a determinar la cosecha anual, establecer el turno y seleccionar las especies más adecuadas para un área (Burkhart & Tomé, 2012). La productividad forestal como concepto biológico complejo se estima indirectamente por medio de modelos de crecimiento (Tamarit-Urias et al., 2014). La forma habitual para referenciar la productividad es utilizar la relación altura dominante-edad mediante el índice de sitio (IS) que se define como la altura promedio que alcanzan los árboles dominantes de un rodal a una edad base determinada (Quiñonez-Barraza et al., 2015). Con las ecuaciones de IS se construyen curvas que muestran el patrón de crecimiento en altura de los árboles de una especie en una ecorregión (Akbas & Senyurt, 2018).
Las curvas de crecimiento que las ecuaciones dinámicas generan deben cumplir con los atributos siguientes: realismo biológico, crecimiento sigmoide, punto de inflexión, polimorfismo, asíntotas variables, invarianza de la edad base y del camino de simulación, y la altura a la edad base debe ser igual al IS (Guerra-Hernández et al., 2021). La invarianza de la edad base se ha formalizado mediante el enfoque de diferencia algebraica, donde un solo parámetro del modelo base se expresa en función del sitio; sin embargo, este enfoque tiene como principal limitante que las ecuaciones derivadas generan únicamente curvas anamórficas o polimórficas, con lo cual se restringe la posibilidad de describir una variedad mayor de condiciones de crecimiento comunes en suelos forestales (Cieszewski, 2021).
En México, pese a las limitaciones de la diferencia algebraica, en las últimas décadas del siglo pasado y aún en la primera del presente, los manejadores forestales utilizaron este enfoque para desarrollar ecuaciones de IS para especies del género Pinus. Por tanto, es necesario actualizar esos modelos mediante enfoques y técnicas de modelación más flexibles. Con ello, de acuerdo con Seki y Sakici (2017) y Özçelik, Cao, Gómez-García, Crecente-Campo, y Eler (2019), se garantizará mayor fiabilidad y precisión en las estimaciones del crecimiento de la altura dominante e IS. Para lograr este propósito, se cuenta con el enfoque de diferencia algebraica generalizada (DAG). En este enfoque, el modelo base se expande para que dos de sus parámetros se expresen en función del sitio mediante una variable teórica X, la cual se asume que engloba los factores de la productividad del sitio. La DAG permite derivar ecuaciones dinámicas que reproducen curvas de IS polimórficas con múltiples asíntotas de mayor realismo biológico (Cieszewski, 2021; Socha, Tyminska-Czabańska, Grabska, & Orzel, 2020; Sprengel, Spiecker, & Wu, 2022).
Pinus chiapensis (Martínez) Andresen es una conífera subtropical endémica de México y Guatemala que forma parte del bosque mesófilo de montaña en Puebla, Veracruz, Guerrero, Chiapas y Oaxaca. La especie destaca por su crecimiento rápido y potencial maderable alto; se distribuye en forma natural en altitudes de 600 a 2 200 m, con una precipitación anual promedio de 1 500 a 2 000 mm temperaturas de 20 a 22 °C y pendientes pronunciadas con suelos ácidos (Perry, 2009). Esta especie desempeña funciones ecológicas y económicas clave en el neotrópico, es un recurso genético valioso (Del Castillo, Trujillo, & Saénz-Romero, 2009) y está catalogada con estatus de protección especial (Chávez-Pascual et al., 2013). La generación de una nueva función de IS mediante DAG podrá ser de capacidad predictiva mayor que los modelos polimórficos previos derivados de diferencia algebraica.
El objetivo de este trabajo fue desarrollar una ecuación dinámica de IS mediante el enfoque de diferencia algebraica generalizada como una herramienta silvícola para describir el patrón de crecimiento en altura dominante y clasificar la productividad de rodales naturales de P. chiapensis de Puebla y Veracruz, México.
Materiales y métodos
Área de estudio
El área de estudio fue la unidad de manejo forestal (UMAFOR) 2103 “Teziutlán” de Puebla y la 3012 “Valle y Cofre de Perote” de Veracruz, México. La UMAFOR 2103 se localiza a 20° 02’ 34” - 19° 36’ 34” N y 97° 43’ 46” - 97° 22’ 23” O, a una altitud promedio de 2 220 m, con temperatura media anual de 12 a 22 °C y los suelos son de tipo Luvisol. La UMAFOR 3012 se ubica a 19° 56’ 16.80” - 19° 16’ 48.00” N y 96° 53’ 42.00” - 97° 25’ 01.20” O, a una altitud entre 500 y 1 600 m. El clima de esta UMAFOR es templado subhúmedo con lluvias en verano con precipitación media de 600 a 1 000 mm, los suelos son tipo Andosol y Litosol, y relieve con pendientes pronunciadas. El estrato arbóreo en ambas regiones se compone principalmente por especies del género Pinus, entre estas P. chiapensis, además de taxones de Quercus y Liquidambar propias del bosque mesófilo de montaña (Rodríguez-Acosta & Arteaga-Martínez, 2005).
Datos
La base de datos se conformó de una muestra de 31 árboles dominantes y codominantes de P. chiapensis, la cual fue similar a la utilizada por Rodríguez-Acosta y Arteaga-Martínez (2005). En ambas UMAFOR, los ejemplares se colectaron en la vegetación que corresponde a bosque mesófilo de montaña; de acuerdo con Ruiz-Jiménez, Téllez-Valdés, y Luna-Vega (2012), este tipo de ecosistemas mantienen condiciones fisiográficas, climáticas y florísticas semejantes cuando las regiones están próximas entre sí, tal como ocurre en el presente caso. En este estudio se amplió el intervalo de edad observado al agregarse 28 pares de observaciones que correspondieron a edades mayores de 55 años, lo que generó 385 pares de datos de edad y altura dominante, cuyos estadísticos básicos se muestran en el Cuadro 1. Los ejemplares representaron varias condiciones de crecimiento del área de estudio. Cada árbol se derribó y seccionó a lo largo del fuste, con inicio en la altura del tocón y después cada 2.55 m de longitud; del extremo inferior de cada sección se obtuvieron rodajas de 5 cm de espesor. La altura dominante y edad se determinaron con la metodología de análisis troncal (Pretzsch, Dauber, & Biber, 2013); la sobrestimación en las mediciones de la altura se corrigió con base en el método de Carmean y la modificación de Newberry indicados por Cancino, Acuña, y Espinosa (2013) y Subedi y Sharma (2010).
Cuadro 1 Estadísticas descriptivas básicas de las variables analizadas en 31 árboles de Pinus chiapensis pertenecientes a dos unidades de manejo forestal (UMAFOR 2103 y 3012) en Puebla y Veracruz, México.
| Variable | Mínimo | Promedio | Máximo | Desviación estándar | Coeficiente de variación (%) |
|---|---|---|---|---|---|
| Altura dominante (m) | 0.10 | 16.50 | 51.13 | 11.07 | 67.07 |
| Edad (años) | 2.00 | 19.48 | 116.00 | 17.33 | 89.99 |
Ecuaciones dinámicas en diferencias algebraicas generalizadas
Un análisis exploratorio sobre el ajuste de 50 ecuaciones dinámicas, derivadas de la aplicación del enfoque de DAG, permitió la preselección de seis funciones como candidatas para el modelado del crecimiento en altura dominante e IS de P. chiapensis (Cuadro 2). En todos los casos, dos parámetros de cada modelo base fueron específicos de la productividad del sitio. Los modelos de crecimiento teóricos usados como base fueron consignados por Burkhart y Tomé (2012), Panik (2014) y Hernández-Cuevas, Santiago-García, De los Santos-Posadas, Martínez-Antúnez, y Ruiz-Aquino (2018). Dichos modelos son el de Chapman-Richards del cual se generó la ecuación dinámica etiquetada como M1, el de Korf del cual se derivaron las ecuaciones M2 y M3, el modelo de Hossfeld que dio lugar a las ecuaciones M4 y M5, así como el de Levakovic II del cual se obtuvo la ecuación M6.
La estructura general implícita de las ecuaciones dinámicas tiene la forma Y = f(t, t 0 , Y 0 , β i ), donde Y es la altura dominante promedio a estimar a la edad t; Y 0 se define como parámetro durante el ajuste, es la altura dominante observada a la edad t 0 ; β i son los parámetros para estimar; y Y 0 y t 0 son condiciones iniciales (Tamarit-Urias et al., 2014).
Cuadro 2 Ecuaciones dinámicas en diferencias algebraicas generalizadas evaluadas para modelar el crecimiento en altura dominante e índice de sitio de Pinus chiapensis.
| Modelo base | Parámetros específicos | X 0 con valores iniciales en t 0 y Y 0 | Ecuación dinámica | ||
|---|---|---|---|---|---|
| Chapman-Richards |
|
|
|
M1 | |
| Korf |
|
|
|
M2 | |
|
|
|
|
M3 | ||
| Hossfeld |
|
|
|
M4 | |
|
|
|
|
M5 | ||
| Levakovic II |
|
|
|
M6 | |
Y = height at age t, Y 0 = height at age t 0 , A = Age (years); a, b and c = parameter of each basic model; X = theoretical variable representing site productivity factors; F 0 and R 0 = components of the dynamic equation; β i = parameters of each dynamic equation to be estimated
Estrategia de ajuste
Los parámetros de las ecuaciones dinámicas se estimaron con el método de variables Dummy, descrito e ilustrado por Sghaier et al. (2012) y en Pyo (2017), el cual es invariante respecto a la edad base. Este método toma en cuenta los errores de medición y los aleatorios de las observaciones, por lo que, a partir de las series de crecimiento individuales estima parámetros locales para cada árbol y parámetros globales para todos los individuos que corresponden a los β i de las ecuaciones.
Debido a que las observaciones corresponden a mediciones múltiples por árbol con estructura jerárquica y longitudinal, además que, en los análisis troncales, las edades a las que se determinan las alturas dominantes no con equidistantes, se presentan los problemas de autocorrelación y heterocedasticidad (Quiñonez-Barraza, García-Espinoza, & Aguirre-Calderón, 2018; Trim, Coble, Weng, Stovall, & Hung, 2020). Para corregir la autocorrelación, la estructura de covarianza que implica al término del error (e ij ) se modeló mediante una estructura continua autorregresiva de orden ρ (Panik, 2014), con lo que el término de error se expresó en la forma siguiente:
donde,
e ij = j-ésimo residuo ordinario del i-ésimo árbol
e ij-k = j-ésimo residuo ordinario del i-k-ésimo árbol
I k = 1 para j > k y es cero para j ≤ k
ρ k = parámetro autorregresivo de orden k a ser estimado
t ij ‒ t ij-k = distancia que separa la j-ésima de la j-k-ésima observación dentro de cada árbol tij > tij-k
ε ij = término del error bajo la condición de independencia.
La heterocedasticidad se corrigió al asumir que la varianza del error (σ2) puede modelarse como una función potencia de la altura dominante predicha
La estimación de los parámetros locales y globales de cada ecuación dinámica, junto con la estructura de covarianza y varianza del error, se programó para un ajuste simultáneo con el procedimiento MODEL del paquete estadístico SAS/ETS (SAS Institute Inc., 2011). La corrección de la autocorrelación se verificó con el estadístico de Durbin-Watson (DW) (Panik, 2014) y la corrección de la heterocedasticidad se comprobó mediante la inspección gráfica de los residuales contra los valores predichos.
La calidad de ajuste de las ecuaciones se evaluó cuantitativamente mediante comparaciones numéricas de los estadísticos de bondad de ajuste: coeficiente de determinación ajustado por el número de parámetros (
donde,
n = número total de observaciones usadas en el ajuste
p = número de parámetros en cada ecuación dinámica.
Los estadísticos R2 adj, RCME, sesgo, AIC y DW se usaron para implementar un criterio de calificación cuantitativo que, con base en Tewari y Singh (2018), obtiene una calificación total para cada modelo. Este sistema de evaluación permite un orden jerárquico de la calidad y desempeño estadístico de los modelos, donde la calificación total más baja corresponde al mejor modelo y viceversa.
El realismo biológico de las ecuaciones dinámicas se evaluó mediante la comparación de las trayectorias de las curvas de crecimiento que cada una describió para valores lógicos de IS, mismas que se sobrepusieron a las tendencias de crecimiento observadas (Tamarit-Urias et al., 2014). Con la ecuación seleccionada y una edad de referencia o edad base (Eb) de 50 años se construyeron curvas de IS para clasificar la productividad maderable de los rodales en baja, media y alta. La ecuación se comparó con el modelo de IS propuesto por Rodríguez-Acosta y Arteaga-Martínez (2005) para la misma especie y región de estudio. Con base en Pretzsch (2009) se aplicó cálculo diferencial para derivar las expresiones que determinan los incrementos corriente y medio anual (ICA e IMA) en altura dominante, así como los valores máximos respectivos para cada clase de IS.
Resultados y discusión
El ajuste de las ecuaciones dinámicas indicó que todos los parámetros globales y los de autocorrelación (ρ
i
) fueron significativos al nivel significancia del 5 % (Cuadro 3). Los estadísticos de bondad de ajuste, mostrados en el Cuadro 4, revelan que el
Cuadro 3 Parámetros y significancia de las ecuaciones dinámicas para modelar el crecimiento en altura dominante e índice de sitio de Pinus chiapensis.
| Ecuación dinámica | Parámetro | Estimador | Error estándar | Valor t | Significancia |
|---|---|---|---|---|---|
| M1 | β1 | 3.1681 | 0.176 | 18.02 | <0.0001 |
| β2 | 0.0408 | 0.003 | 15.22 | <0.0001 | |
| ρ1 | 0.4582 | 0.023 | 20.05 | <0.0001 | |
| ρ2 | -0.5792 | 0.025 | -22.82 | <0.0001 | |
| M2 | β1 | 0.6234 | 0.037 | 16.92 | <0.0001 |
| β2 | 6.775 | 0.452 | 15 | <0.0001 | |
| ρ1 | 0.5193 | 0.029 | 17.71 | <0.0001 | |
| ρ2 | 0.6576 | 0.031 | 21.49 | <0.0001 | |
| ρ3 | 0.5454 | 0.036 | 15.14 | <0.0001 | |
| M3 | β2 | 27.7074 | 1.534 | 18.06 | <0.0001 |
| β3 | 0.6001 | 0.039 | 15.55 | <0.0001 | |
| ρ1 | 0.5244 | 0.03 | 17.38 | <0.0001 | |
| ρ2 | -0.655 | 0.032 | -20.71 | <0.0001 | |
| ρ3 | 0.5313 | 0.039 | 13.66 | <0.0001 | |
| M4 | β1 | 4 092.5190 | 550.5 | 7.43 | <0.0001 |
| β2 | 1.4722 | 0.062 | 23.75 | <0.0001 | |
| ρ1 | 1.0223 | 0.06 | 17.07 | <0.0001 | |
| ρ2 | 0.8922 | 0.045 | 19.65 | <0.0001 | |
| ρ3 | 0.657 | 0.054 | 12.28 | <0.0001 | |
| M5 | β1 | 3 199.3280 | 343.2 | 9.32 | <0.0001 |
| β2 | 1.3557 | 0.054 | 25.17 | <0.0001 | |
| ρ1 | 0.449 | 0.023 | 19.48 | <0.0001 | |
| ρ2 | -0.5703 | 0.026 | -22.02 | <0.0001 | |
| M6 | β1 | 7.4792 | 0.984 | 7.6 | <0.0001 |
| β2 | 11.9382 | 2.337 | 5.11 | <0.0001 | |
| ρ1 | 0.4474 | 0.023 | 19.54 | <0.0001 | |
| ρ2 | -0.5693 | 0.025 | -22.42 | <0.0001 |
Cuadro 4 Estadísticos de bondad de ajuste de las ecuaciones evaluadas para modelar el crecimiento en altura dominante e índice de sitio de Pinus chiapensis.
| Ecuación dinámica | R2 adj | RCME | Sesgo | AIC | DW | Orden ρ | Calificación total |
|---|---|---|---|---|---|---|---|
| M1 | 0.9808 | 1.54 | -0.004 | 304.11 | 1.61 | AR(2) | 25 |
| M2 | 0.9829 | 1.45 | 0.2032 | 258.09 | 1.82 | AR(3) | 14 |
| M3 | 0.9822 | 1.48 | 0.2042 | 276.07 | 1.84 | AR(3) | 17 |
| M4 | 0.984 | 1.4 | 0.1277 | 239.65 | 1.92 | AR(3) | 8 |
| M5 | 0.9817 | 1.5 | 0.0581 | 287.02 | 1.71 | AR(2) | 21 |
| M6 | 0.9818 | 1.49 | 0.0754 | 284.34 | 1.7 | AR(2) | 20 |
R2 adj = coeficiente de determinación ajustado por el número de parámetros; RCME = raíz del cuadrado medio del error; AIC = criterio de información de Akaike, DW = Durbin-Watson; AR = estructura autorregresiva.
Los residuales estandarizados contra los predichos de las ecuaciones mantuvieron una distribución con tendencia aleatoria en todos los casos (Figura 1), lo que sugiere que la heterocedasticidad se redujo al mínimo para lograr un patrón homocedástico; por lo tanto, se infiere que las ecuaciones evaluadas se ajustan de forma aceptable a los datos observados. Bajo este escenario, ambas correcciones permiten que los estimadores obtenidos en el ajuste sean de varianza mínima, insesgados y más eficientes, lo que mejora sustancialmente las propiedades estadísticas y predictivas de las ecuaciones. De este modo, es posible estimar intervalos de confianza realistas y efectuar los contrastes estadísticos de significación sobre los parámetros con propiedad.

Figura 1 Comportamiento gráfico de los residuales estandarizados con respecto a los valores predichos de la altura dominante de Pinus chiapensis para las ecuaciones evaluadas.
La Figura 2 muestra las tendencias de crecimiento en altura dominante que las ecuaciones dinámicas reproducen para valores de IS de 22, 27, 32, 37 y 42 m (Eb = 50 años), sobrepuestas a las trayectorias de crecimiento observadas. Una inspección detallada muestra que las funciones M2 y M3 tienden a sobrestimar las alturas dominantes en la mejor calidad de sitio en edades superiores a 60 años; antes de esta edad, la tendencia sugiere ligera subestimación. Por su parte, las ecuaciones M4 y M5 tienden a ligeras subestimaciones en las mejores calidades de IS para todo el intervalo de edad. La ecuación M1 exhibió el comportamiento más deficiente, ya que subestima en todos los IS a edades tempranas. Esta tendencia de subestimación se mantiene para los mejores sitios a edades maduras y longevas.
En contraparte, la ecuación dinámica M6 (modelo base de Levakovic II) mostró mejor realismo biológico, porque las tendencias de crecimiento que describe se sobreponen de mejor manera a las trayectorias observadas en todos los niveles de IS y en todo el intervalo de edad analizado. Por esta razón, la ecuación M6 se seleccionó para describir el patrón de crecimiento en altura dominante y para clasificar la productividad de rodales naturales de P. chiapensis en la zona de estudio. Esta selección ratifica lo señalado por Özçelik et al. (2019), en el sentido de que las funciones con los mejores estadísticos de ajuste no siempre reproducen de mejor manera las tendencias observadas. En este caso, estadísticamente, la ecuación M6 se posicionó en el antepenúltimo lugar; sin embargo, dado el realismo biológico que recrea, es claro que tiene la capacidad predictiva más alta. Esta elección más que un ejercicio puro de inferencia estadística fue producto de un compromiso y balance entre desempeño estadístico y razones biológicas. La ecuación M6 tiene parsimonia con solo dos parámetros, suficientes para revelar las verdaderas tendencias de crecimiento en las alturas dominantes observadas.

Figura 2 Comportamiento de las curvas de índice de sitio (IS) reproducidas por las ecuaciones dinámicas evaluadas sobrepuestas a las trayectorias de crecimiento observadas en Pinus chiapensis.
Con los valores de los parámetros de la ecuación dinámica M6 y una Eb = 50 años, se construyeron curvas de IS para tres niveles de productividad etiquetadas como baja, media y alta, cuyos valores medios corresponden a los IS de 25, 32 y 39 m, respectivamente (Figura 3a). Estas curvas junto con sus límites respectivos, en particular el inferior y superior de las clases baja y alta, cubrieron la amplitud y las tendencias de todas las series de crecimiento en el intervalo de edad analizado. Por esta razón, las clases de IS establecidas se consideraron suficientes para la clasificación de la productividad de los rodales naturales de P. chiapensis en la zona de estudio.
El enfoque de DAG para generar la ecuación M6 en combinación con la técnica de variables Dummy para ajustarla, aunado a las correcciones de autocorrelación y heterocedasticidad, logran una ecuación dinámica robusta para realizar predicciones fiables. Este aspecto se aprecia mejor al contrastar las curvas de IS con las que reproduce el modelo de IS propuesto por Rodríguez-Acosta y Arteaga-Martínez (2005) (Figura 3b), que corresponde a un modelo polimórfico, cuyo modelo base es el de Chapman-Richards. Se evidencia que para una Eb = 50 años y los valores de IS referidos, el modelo polimórfico no describe adecuadamente el crecimiento en altura dominante de P. chiapensis. La mayor deficiencia se aprecia para la clase de IS alta, donde las tendencias de crecimiento son ilógicas porque las curvas colapsan, debido a que el valor de la asíntota horizontal del modelo es menor que la altura real que los datos observados sugieren. Las diferencias pueden explicarse desde el punto de vista matemático-estadístico, porque para generar el modelo polimórfico usaron el enfoque de diferencias algebraicas, en el que solo el parámetro de forma fue dependiente del sitio y, además, no se corrigieron la autocorrelación ni la heterocedasticidad.
Las alturas dominantes de 25, 32 y 39 m estimadas por la ecuación nueva, que equivalen a los valores de IS (Eb = 50 años), son ligeramente superiores a 24.2, 29.7 y 35.5 m (Eb = 50 años), alturas determinadas mediante el modelo propuesto por Sánchez y Del Castillo (2001). Estos autores utilizaron datos de sitios temporales y ajustaron modelos lineales por condición de crecimiento en la estimación del IS para rodales naturales coetáneos de P. chiapensis en la Sierra Norte de Oaxaca, México. Esta discrepancia sugiere que las condiciones de crecimiento dadas por condiciones físicas y ambientales en los sitios de Veracruz y Puebla son comparativamente mejores. En este sentido, Del Castillo et al. (2009) refieren que árboles individuales longevos de esta especie, en las mejores calidades de estación, pueden alcanzar alturas de hasta 50 m, tal como se evidencia en las observaciones del presente estudio para la clase alta de IS.
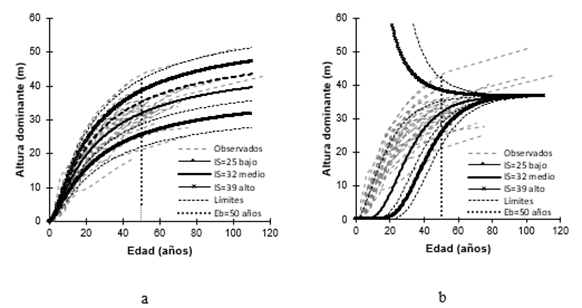
Figura 3 Comparativo gráfico de las curvas de índice de sitio (IS) generadas por (a) la nueva ecuación dinámica y (b) el modelo de Rodríguez-Acosta y Arteaga-Martínez (2005).
Un análisis de la función polimórfica derivada mediante diferencias algebraicas con el modelo base de Hossfeld IV, desarrollado por Fierros-Mateo, De los Santos-Posadas, Fierros-González, y Cruz-Cobos (2017) para la misma especie, mostró que con una Eb = 8 años y clases de IS de 6, 10 y 14 m, se pudo modelar el crecimiento en altura dominante de los datos observados en forma aceptable hasta los 20 años; posteriormente, tiende a un techo asintótico máximo de 33 m, el cual es menor que las alturas máximas observadas. Esta disparidad, aun cuando la zona de estudio fue similar (UMAFOR 2103), puede explicarse porque en dicho estudio se procesó información de plantaciones comerciales jóvenes con un intervalo de edad de 1 a 8 años.
Con valores de IS de 25, 32 y 39 m y Eb = 50 años, la ecuación dinámica en DAG del modelo base de Chapman-Richards [
Las expresiones 1 y 2, cuyo componente X 0 está indicado en el Cuadro 2, derivadas de la ecuación dinámica M6, son útiles para construir curvas de IMA e ICA por clase de IS. En tanto que las expresiones 3 y 4, derivadas de la misma ecuación, sirven para determinar el incremento medio anual máximo (IMAmáx) y el incremento corriente anual máximo (ICAmáx) por nivel de productividad.
Las determinaciones de ICA, IMA y los incrementos máximos evidencian que la ecuación M6 satisface consistentemente el supuesto lógico de que la edad a la que se alcanzan tales incrementos decrece conforme el IS aumenta (Cuadro 5). También se cumple que los incrementos son mayores a medida que el valor del IS es mayor. De acuerdo con Seki y Sakici (2017) y Socha et al. (2020), se ratifica que la ecuación dinámica desarrollada cumple a cabalidad con las leyes de crecimiento biológico.
Cuadro 5 Edades a las que suceden los incrementos medio y corriente anual (IMA e ICA) máximos e incrementos de la altura dominante por clase de índice de sitio (IS) para Pinus chiapensis.
| Clase de IS (m) | Edad IMAmáx(años) | IMAmáx (m∙año-1) | Edad ICAmáx (años) | ICAmáx (m∙año-1) |
|---|---|---|---|---|
| 25 | 12.4 | 0.8 | 6.2 | 0.95 |
| 32 | 11.08 | 1.07 | 5.54 | 1.25 |
| 39 | 10.11 | 1.35 | 5.05 | 1.57 |
El hecho de que se cumplan estos supuestos relevantes coincide con lo referido por Ercanli, Kahriman, y Yavuz (2014) y por Seki y Sakici (2017), en el sentido de que las ecuaciones dinámicas derivadas mediante DAG poseen mayor flexibilidad para modelar una variedad amplia de tendencias de crecimiento. Tales ecuaciones cumplen con todos los atributos deseables que se traducen en consistencia y realismo biológico, por lo que deben de ser utilizadas para modelar el crecimiento de bosques naturales y artificiales. En este contexto, se considera que, para el desarrollo de nuevas ecuaciones dinámicas de IS en México, se debe priorizar el uso del enfoque de DAG y que sean dos los parámetros específicos del sitio en cada modelo de crecimiento base que se utilice. Además, se debe usar algún método de ajuste que sea invariante de la edad base, tal como el de variables Dummy (Sghaier et al., 2012), el procedimiento iterativo anidado (Sharma, Brunner, Eid, & Øyen, 2011) o bien la técnica de modelo de efectos mixtos (Tamarit-Urias et al., 2014). También se debe corregir la autocorrelación y la heterocedasticidad para obtener modelos más robustos que cumplan con los supuestos teóricos del análisis de regresión. Esto permitirá estar al nivel de la tendencia internacional ya que, como refieren Özçelik et al. (2019), es común que cuando se disponga de datos provenientes de análisis troncales o de parcelas permanentes con remediciones se usen los refinamientos matemáticos y estadísticos referidos.
Mediante el contraste de las edades en que ocurren los turnos técnicos (IMA = ICA) en la altura dominante, por clase de IS de la ecuación dinámica M6, se infiere que P. chiapensis tiende a tasas constantes de crecimiento en la zona de estudio. Lo anterior debido a que las diferencias en la ocurrencia son pequeñas, efecto que conduce a que las curvas de IS pudieran considerarse de tipo cuasianamórficas. Esta tendencia de crecimiento también fue observada por Calegario, Daniels, Maestri, y Neiva (2005) en clones del género Eucalyptus en los estados Bahía y Espírito de Brasil.
Conclusiones
La ecuación dinámica desarrollada de índice de sitio, derivada del modelo de crecimiento de Levakovic II, tiene base biológica y rigor matemático-estadístico sólido; además, genera curvas polimórficas con asíntotas variables que se apegan con fidelidad a la realidad del crecimiento observado. Asimismo, la ecuación cumple con atributos deseables lógicos entre los que destacan realismo biológico, invarianza con respecto a la edad base y de ruta, flexibilidad, parsimonia y consistencia. Se recomienda su uso como una herramienta actual y fiable para la gestión silvícola en términos de la predicción del crecimiento en altura dominante y determinación del potencial productivo de rodales naturales de Pinus chiapensis en Puebla y Veracruz, México. La ecuación podrá utilizarse como componente básico de sistemas de crecimiento y rendimiento maderable. Esta ecuación nueva presentó mejor desempeño dado por una capacidad predictiva superior, comparada con un modelo polimórfico de índice de sitio previo para la misma especie y región.
References
Akbas, U., & Senyurt, M. (2018). Site quality estimations based on the generalized algebraic difference approach: a case study in Çankiri forests. Revista Árvore, 42(3), e420311. doi: 10.1590/1806-90882018000300011 [ Links ]
Burkhart, H. E., & Tomé, M. (2012). Modeling forest trees and stands. New York, NY, USA: Springer-Verlag. [ Links ]
Calegario, N., Daniels, R. F., Maestri, R., & Neiva, R. (2005). Modeling dominant height growth based on nonlinear mixed-effects model: a clonal Eucalyptus plantation case study. Forest Ecology and Management, 204(1), 11-21. doi: 10.1016/j.foreco.2004.07.051 [ Links ]
Cancino, J., Acuña, E., & Espinosa, M. (2013). Combining ring counting and ring width for estimating height in stem analysis. Forest Science, 59(6), 599‒609. doi: 10.5849/forsci.12-028 [ Links ]
Chávez-Pascual, E. Y., Rodríguez-Ortiz, G., Carrillo-Rodríguez, J. C., Enríquez-del Valle, J. R., Chávez-Servia, J. L., & Campos-Ángeles, G. V. (2013). Factores de expansión de biomasa aérea para Pinus chiapensis (Mart.) Andresen. Revista Mexicana de Ciencias Agrícolas, 6, 1273‒1284. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-09342013001000018 [ Links ]
Del Castillo, R. F., Trujillo, A. S., & Saénz-Romero, C. (2009). Pinus chiapensis, a keystone species: Genetics, ecology, and conservation. Forest Ecology and Management, 257(11), 2201‒2208. doi: 10.1016/j.foreco.2009.03.004 [ Links ]
Ercanli, İ., Kahriman, A., & Yavuz, H. (2014). Dynamic base-age invariant site index models based on generalized algebraic difference approach for mixed Scots pine (Pinus sylvestris L.) and Oriental beech (Fagus orientalis Lipsky) stands. Turkish Journal of Agriculture and Forestry, 38(1), 134‒147. doi: 10.3906/tar-1212-67 [ Links ]
Fierros-Mateo, R., De los Santos-Posadas, H. M., Fierros-González, M. A., & Cruz-Cobos, F. (2017). Crecimiento y rendimiento maderable en plantaciones de Pinus chiapensis (Martínez) Andresen. Agrociencia, 51(2), 201‒214. Retrieved from https://agrociencia-colpos.mx/index.php/agrociencia/article/view/1287/1287 [ Links ]
Guerra-Hernández, J., Arellano-Pérez, S., González-Ferreiro, E., Pascual, A., Altelarrea, V. S., Ruiz-González, A. D., & Álvarez-González, J. G. (2021). Developing a site index model for P. pinaster stands in NW Spain by combining bi-temporal ALS data and environmental data. Forest Ecology and Management, 481, 118690. doi: 10.1016/j.foreco.2020.118690 [ Links ]
Hernández-Cuevas, M., Santiago-García, W., De los Santos-Posadas, H. M., Martínez-Antúnez, P., & Ruiz-Aquino, F. (2018). Modelos de crecimiento en altura dominante e índices de sitio para Pinus ayacahuite Ehren. Agrociencia, 52(3), 437‒453. Retrieved from https://agrociencia-colpos.mx/index.php/agrociencia/article/view/1680/1680 [ Links ]
Özçelik, R., Cao, V. Q., Gómez-García, E., Crecente-Campo, F., & Eler, Ü. (2019). Modeling dominant height growth of cedar (Cedrus libani A. Rich) stands in Turkey. Forest Science, 65(6), 725‒733. doi: 10.1093/forsci/fxz038 [ Links ]
Panik, M. J. (2014). Growth curve modeling: Theory and applications. Hoboken, NJ, USA: Wiley. [ Links ]
Perry, J. P. (2009). The pines of Mexico and Central America. Portland, OR, USA: Timber Press. [ Links ]
Pretzsch, H. (2009). Forest dynamics, growth and yield. From measurement to model. Berlin, Germany: Springer. [ Links ]
Pretzsch, H., Dauber, E., & Biber, P. (2013). Species-specific and ontogeny-related stem allometry of european forest trees: Evidence from extensive stem analyses. Forest Science, 59(3), 290‒302. doi: 10.5849/forsci.11-102 [ Links ]
Pyo, J. (2017). Developing the site index equation using a generalized algebraic difference approach for Pinus densiflora in central region, Korea. Forest Science and Technology, 13(2), 87-91. doi: 10.1080/21580103.2017.1308889 [ Links ]
Quiñonez-Barraza, G., De los Santos-Posadas, H. M., Cruz-Cobos, F., Velázquez-Martínez, A., Ángeles-Pérez, G., & Ramírez-Valverde, G. (2015). Site index with complex polymorphism of forest stands in Durango, Mexico. Agrociencia, 49(4), 439-454. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-31952015000400007 [ Links ]
Quiñonez-Barraza, G., García-Espinoza, G. G., & Aguirre-Calderón, O. A. (2018). How to correct the heteroscedasticity and autocorrelation of residuals in taper and height growth models? Revista Mexicana de Ciencias Forestales, 9(49), 28-59. doi: 10.29298/rmcf.v9i49.151 [ Links ]
Rodríguez-Acosta, M., & Arteaga-Martínez, B. (2005). Índice de sitio para Pinus chiapensis (Martínez) Andresen, en los estados de Veracruz y Puebla, México. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 11(1), 39‒44. Retrieved from https://revistas.chapingo.mx/forestales/revista/articulos/viewer.html?file=rchscfaXI428.pdf [ Links ]
Ruiz-Jiménez, C. A., Téllez-Valdés, O., & Luna-Vega, I. (2012). Clasificación de los bosques mesófilos de montaña de México: afinidades de la flora. Revista Mexicana de Biodiversidad, 83, 1110‒1144. doi: 10.7550/rmb.29383 [ Links ]
Ryan, T. P. (2009). Modern regression methods. New York, USA: Wiley. [ Links ]
Sánchez, V. N. M., & Del Castillo, S. R. F. (2001). Calidad de estación para Pinus chiapensis (Mart.) Andresen en El Rincón, Oaxaca, México. Foresta Veracruzana, 3(2), 9‒12. Retrieved from https://www.redalyc.org/articulo.oa?id=49703202 [ Links ]
Seki, M., & Sakici, O. E. (2017). Dominant height growth and dynamic site index models for Crimean pine in Kastamonu-Taşköprü region of Turkey. Canadian Journal of Forest Research, 47(11), 1441‒1449. doi: 10.1139/cjfr-2017-0131 [ Links ]
Sghaier, T., Palahi, M., Garchi, S., Bonet, J. A., Ammari, Y., & Pique, M. (2012). Modeling dominant height growth in planted Pinus pinea stands in Northwest of Tunisia. International Journal of Forestry Research, ID 902381. doi: 10.1155/2012/902381 [ Links ]
Sharma, R. P., Brunner, A., Eid, T., & Øyen, B. H. (2011). Modelling dominant height growth from national forest inventory individual tree data with short time series and large age errors. Forest Ecology and Management, 262(12), 2162-2175. doi: 10.1016/j.foreco.2011.07.037 [ Links ]
Socha, J., Tyminska-Czabańska, L., Grabska, E., & Orzel, S. (2020). Site index models for main forest-forming tree species in Poland. Forests, 11, 301. doi: 10.3390/f11030301 [ Links ]
Sprengel, L., Spiecker, H., & Wu, S. (2022). Two subject specific modelling approaches to construct base-age invariant polymorphic site index curves with varying asymptotes. Silva Fennica, 56(1), 10544. doi: 10.14214/sf.10544 [ Links ]
Statistical Analysis System (SAS Institute Inc.) (2011). SAS/STAT User’s Guide, version 9.3. Cary, NC, USA: Author. [ Links ]
Subedi, N., & Sharma, M. (2010). Evaluating height-age determination methods for Jack pine and black spruce plantations using stem analysis data. Northern Journal of Applied Forestry, 27(2), 50‒55. doi: 10.1093/njaf/27.2.50 [ Links ]
Tamarit-Urias, J. C., De los Santos-Posadas, H. M., Aldrete, A., Valdez-Lazalde, J. R., Ramírez-Maldonado, H., & Guerra-De la Cruz, V. (2014). Ecuaciones dinámicas de índice de sitio para Tectona grandis en Campeche, México. Agrociencia, 48(2), 225‒238. Retrieved from https://www.agrociencia-colpos.mx/index.php/agrociencia/article/view/1077/1077 [ Links ]
Tewari, V. P., & Singh, B. (2018). Total wood volume equation for Tectona grandis Linn F. stands in Gujarat, India. Journal of Forest and Environmental Science, 34(4), 313‒320. doi: 10.7747/JFES.2018.34.4.313 [ Links ]
Trim, K. R., Coble, D. W., Weng, Y., Stovall, J. P., & Hung, I. K. (2020). A new site index model for intensively managed loblolly pine (Pinus taeda) plantations in the west Gulf Coastal Plain. Forest Science, 66(1), 2‒13. doi: 10.1093/forsci/fxz050 [ Links ]
Vargas-Larreta, B., Corral-Rivas, J. J., Aguirre-Calderón, O. A., López-Martínez, J. O., de los Santos-Posadas, H. M., Zamudio-Sánchez, F. J.,. . . Aguirre-Calderón, C. G. (2017). SiBiFor: Forest Biometric System for forest management in Mexico. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 23(3), 437-455. doi: 10.5154/r.rchscfa.2017.06.040 [ Links ]
Recibido: 11 de Mayo de 2021; Aprobado: 06 de Mayo de 2022
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
