










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versão On-line ISSN 2007-4018versão impressa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.28 no.1 Chapingo Jan./Abr. 2022 Epub 02-Fev-2024
https://doi.org/10.5154/r.rchscfa.2021.07.046
Artículo científico
Producción y viviparidad de nogal pecanero (Carya illinoinensis [Wangenh.] K. Koch) en relación con la humedad del suelo
1 Universidad Autónoma Chapingo, Unidad Regional Universitaria de Zonas Áridas. Carretera Gómez Palacio-Ciudad Juárez, km 40. C. P. 35230. Bermejillo, Durango, México.
2 Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias, Campo Experimental La Laguna. Bulevar José Santos Valdez núm. 1200, col. Centro. C. P. 27440. Matamoros, Coahuila, México.
3 Instituto Nacional de Investigaciones Forestales Agrícolas, Campo Experimental Zacatecas. Carretera Zacatecas-Fresnillo km 24.5. C. P. 98500. Calera de Víctor Rosales, Zacatecas, México.
4 Universidad Autónoma Chapingo, División de Ciencias Forestales. Carretera México-Texcoco km 38.5. C. P. 56230. Texcoco, Estado de México, México.
Introducción:
La viviparidad en nogal pecanero (Carya illinoinensis [Wangenh.] K. Koch) ha incrementado considerablemente. Este fenómeno genético-ambiental puede ser controlado con el manejo de humedad del suelo y otras variables de calidad.
Objetivo:
Determinar el efecto del contenido de humedad del suelo sobre el rendimiento, tamaño de nuez, porcentaje de almendra y nuez germinada en el árbol.
Materiales y métodos:
Árboles de nogal de 40 años fueron regados por 40 emisores con gasto de 3.2 L∙h-1. A partir de la etapa fenológica de endurecimiento de cáscara, el tiempo de riego fue de 4, 6 y 8 h, correspondientes a niveles de humedad de 0.257, 0.327 y 0.380 m3 de agua por m3 de suelo, a profundidad de 40 a 80 cm. El rendimiento por árbol, porcentaje de nuez germinada, tamaño de fruto y porcentaje de almendra se evaluaron durante los ciclos productivos 2016 y 2017.
Resultados y discusión:
Durante los dos ciclos de producción, cuando el nivel de humedad en el suelo aumentó de 0.257 a 0.380 m3∙m-3, la producción de nuez por árbol, la longitud y diámetro del fruto, y el porcentaje de almendra incrementaron en promedio 23.8 %, 25.3 y 10 %, y 1.8 %, respectivamente; sin embargo, la viviparidad incrementó 11.2 %, lo cual se vio reflejado en la disminución del porcentaje de nuez comercial. Los tratamientos 0.327 y 0.380 m3∙m-3 tuvieron un efecto similar (P > 0.05) en dichas variables.
Conclusiones:
El menor nivel de humedad (0.257 m3∙m-3), a partir del llenado de almendra, disminuyó la viviparidad y aumentó el porcentaje de nuez comercial.
Palabras clave: nuez; variedad Western; germinación prematura, tamaño de fruto, porcentaje de almendra
Introduction:
Viviparity in pecan nut (Carya illinoinensis [Wangenh.] K. Koch) has increased considerably. This genetic-environmental phenomenon can be controlled with soil moisture management and other quality variables.
Objective:
To determine the effect of soil moisture content on yield, nut size, kernel percentage and germinated nut on the tree.
Materials and methods:
Forty-year-old pecan nut trees were irrigated by 40 emitters with an output of 3.2 L∙h-1. From the phenological stage of shell hardening, irrigation time was 4, 6 and 8 h, corresponding to moisture levels of 0.257, 0.327 and 0.380 m3 of water per m3 of soil, at depths of 40 to 80 cm. Yield per tree, germinated nut percentage, fruit size and kernel percentage were evaluated during the production cycles of 2016 and 2017 production cycles.
Results and discussion:
During the two production cycles, when soil moisture level increased from 0.257 to 0.380 m3∙m-3, nut yield per tree, fruit length and diameter, and kernel percentage increased on average 23.8 %, 25.3 and 10 %, and 1.8 %, respectively; however, vivipary increased 11.2 %, which was reflected in the decrease of commercial pecan nut percentage. Treatments 0.327 and 0.380 m3∙m-3 had a similar effect (P > 0.05) on these variables.
Conclusions:
The lower moisture level (0.257 m3∙m-3), starting at the kernel filling stage, decreased vivipary and increased commercial kernel percentage.
Keywords: nut; Western variety; premature germination, fruit size, kernel percentage
Introducción
La viviparidad o germinación prematura de la nuez es un fenómeno que consiste en la continuación del crecimiento del embrión durante la maduración del fruto, cuando este se encuentra aún en el árbol (Wood, 2015). Este fenómeno recurrente se ha manifestado considerablemente tanto en árboles nativos que crecen en poblaciones espontáneas (Sparks, 2005) como en las huertas comerciales (León, 2014). La viviparidad es resultado de un efecto genético-ambiental (Wells, 2017) relacionado con el contenido hormonal (Wood, 2015), las temperaturas cálidas de otoño (León, 2014) y el contenido elevado de humedad en el suelo (Godoy & López, 2000; Thompson, 2005), previo al inicio de apertura del ruezno o madurez de la nuez.
El grado de germinación prematura también está relacionado con la susceptibilidad de la variedad (Wells, 2017). Wichita, Mahan, Shawnee y Cheyenne son variedades muy susceptibles; Western presenta un grado moderado y otras como Caddo y Sioux tienen baja susceptibilidad (Aguilar et al., 2015). Las regiones con mayor incidencia de nuez germinada incluyen la costa de Hermosillo, en la que se ha observado 31 % para la variedad Wichita y 27 % para la variedad Western (García-Moreno, Báez-Sañudo, Mercado-Ruiz, García-Robles, & Núñez-Moreno, 2020). La región norte de Coahuila, Comarca Lagunera y algunas áreas de Nuevo León y Tamaulipas (Aguilar et al., 2015) presentan el mismo problema. Este fenómeno también se manifiesta en los estados productores del sureste de Estados Unidos con germinación que supera 50 % (Ou et al., 1994; Smith, 2012).
La germinación prematura de la nuez incrementa cuando las temperaturas son de 30 a 35 °C durante septiembre y octubre, que coincide con la maduración de la nuez y apertura del ruezno (León, 2014; Thompson, 2005). Igualmente, la viviparidad incrementa en años de producción elevada, la cual está estrechamente relacionada con la humedad del suelo (Garrot, Kilbby, Fangmeier, Husman, & Ralowics, 1993; Sparks, Reid, Yates, Smith, & Stevenson, 1995; Wood, 2015) y duración del periodo en el que las nueces permanecen en el árbol antes de cosechar (Stein, 1985).
Muchos cultivos presentan viviparidad (Farnsworth, 2000), aunque el problema se exhibe en nogal pecanero (Carya illinoinensis [Wangenh.] K. Koch) no sucede en otras especies productoras de nuez (Cohen et al., 1997; Wood, 2015). En esta especie, las evidencias indican que la predisposición de las nueces para germinar se debe a una concentración baja de ácido abscísico (León, 2014) y su interacción con las giberelinas (León, 2014; White, Proebsting, Hedden, & Rivin, 2000). Cuando no hay pérdida de humedad en el ruezno, la semilla continuará su desarrollo y germinará en el árbol antes de ser cosechada (Wells, 2017). Si la humedad elevada del suelo induce la viviparidad, entonces, la disminución en el contenido de humedad podría reducir la germinación de la nuez en el árbol; sin embargo, su efectividad debe ser estudiada y comprobada. Por lo tanto, el objetivo del estudio fue evaluar el efecto de contenidos de humedad en el suelo sobre el rendimiento de C. illinoinensis, la germinación prematura de la nuez, el tamaño del fruto y contenido de almendra en nogal pecanero, durante los ciclos productivos 2016 y 2017.
Materiales y métodos
Descripción del área de estudio
El estudio se realizó en árboles de nogal de 40 años, variedad Western, durante el periodo julio-octubre de 2016 y 2017. Los árboles están situados en el municipio de Viesca, Coahuila, México, con coordenadas geográficas de 25° 20’ 28’’ LN y 102° 10’ y 48º 16’ LO, a una altitud de 1 100 m. El clima pertenece a un tipo BWhw (e), el cual se interpreta como muy árido, semicálido con lluvias en verano y de amplitud térmica extremosa con una temperatura media anual entre 18 y 22 °C. La precipitación promedio anual varía entre 200 y 300 mm con régimen de lluvias en los meses de abril a noviembre y escasas en el resto del año (Servicio Meteorológico Nacional [SMN], 2010). El suelo es de textura franco-arcillosa, con capacidad de campo de 0.35 m3∙m-3 (valores de peso en contenido hídrico, calculados a volumen de agua/suelo) y punto de marchitez permanente de 0.19 m3∙m-3. El esparcimiento entre árboles es de 12 m x 12 m y están plantados bajo un diseño de marco real. El sistema de riego es por goteo con dos líneas a cada lado del árbol colocadas a 2 y 3 m a partir del tronco a una profundidad de 40 cm.
Unidades experimentales y tratamientos
Se seleccionaron árboles representativos de la huerta con circunferencia promedio de tronco de 90 cm, medida a 60 cm a partir del suelo, con una altura aproximada de 15 m y una densidad foliar de 3.5 m2∙m-3. Cada árbol se regó con 40 emisores con gasto de 3.2 L∙h-1. Los riegos con 3.5 h diarias iniciaron en marzo, durante la etapa fenológica de brotación. El tiempo de riego incrementó con el avance de la estación, hasta 6 h diarias. En la etapa fenológica del llenado de almendra, el tiempo de riego fue de 4, 6 y 8 h en tres secciones de riego, respectivamente, con el fin de establecer los niveles de humedad (tratamientos) de 0.257, 0.327 y 0.380 m3 de agua por m3 de suelo. Las unidades de muestreo (tres árboles) se seleccionaron en la parte media de cada sección de riego, la cual incluía 50 árboles respectivamente, considerando el criterio sugerido por Prodan (1968) para muestreo en especies arbóreas. El nivel de humedad en el suelo se registró con base en el contenido volumétrico de agua, expresado en metros cúbicos de agua por metro cúbico de suelo, a profundidad de 80 cm, donde se colocó cada sensor de humedad. Los sensores fueron de tipo capacitivo (EC-5 DECAGON DEVICES Inc., Pullman WA, EUA) debidamente calibrados, los cuales miden la constante dieléctrica del suelo, conectados a un datalogger (Em50 DECAGON DEVICES Inc., Pullman WA, EUA) para registrar las lecturas que, posteriormente, se correlacionaron con el contenido de humedad. El registro se hizo diariamente y los datos se almacenaron en archivo de formato de hoja electrónica, durante el periodo julio-octubre del 2016 y 2017.
Variables respuesta y análisis estadístico
Las variables evaluadas fueron producción de nuez (kg) por árbol al momento de la cosecha, la cual se llevó a cabo con vibradora mecánica el 11 y 16 de septiembre de 2016 y 2017, respectivamente; tamaño de fruto, expresado en longitud y diámetro, medido con vernier en una muestra de 50 nueces por árbol; porcentaje de nuez germinada y nuez comercial resultado del cociente del número de nueces para cada condición (n) y el total de las nueces colectadas (N), multiplicado por 100; porcentaje de almendra de una muestra de 20 nueces por árbol, las cuales se rompieron con un cascanueces y se separó la almendra de la cáscara. El peso (g) de la almendra se dividió entre el peso total de la muestra y se multiplicó por 100. El diseño experimental fue completamente al azar en medidas repetidas en el tiempo; el factor de estudio correspondió al contenido de humedad disponible en el suelo (0.380, 0.327 y 0.257 m3∙m-3). Un árbol se consideró como unidad experimental. La distribución normal de los datos se analizó utilizando la prueba de Shapiro-Wilk; posteriormente, un análisis de varianza se realizó para determinar la significancia de los tratamientos y la prueba de Tukey (P ≤ 0.05) para la separación de las medias. Para ello se usaron los programas SAS versión 9.0 (SAS Institute, 2002) y Microsoft Excel versión 2013.
Resultados
Dinámica de la humedad del suelo
La Figura 1 muestra el comportamiento de la humedad del suelo bajo tres contenidos volumétricos de agua, a profundidad de 80 cm del suelo durante el periodo julio-octubre de 2016. El suelo regado con 0.380 m3∙m-3 tuvo humedad hídrica superior que a capacidad de campo (CC), a partir de la etapa fenológica de endurecimiento de cáscara que ocurre en la última semana de julio (Figura 1). El contenido de humedad en este tratamiento no presentó valores de saturación durante la estación; sin embargo, a finales de septiembre se observó una tendencia hacia un contenido hídrico mayor. El área regada con 0.327 m3∙m-3, que correspondió al riego aplicado por el productor, conservó la humedad disponible cercana a capacidad de campo durante julio y agosto; a partir de septiembre, disminuyó conforme avanzó el ciclo productivo. Por otra parte, el suelo con menor volumen de agua aplicado (0.257 m3∙m-3) mostró los valores más bajos durante la estación y disminuyó a partir de septiembre, como sucedió en el tratamiento de 0.327 m3∙m-3.
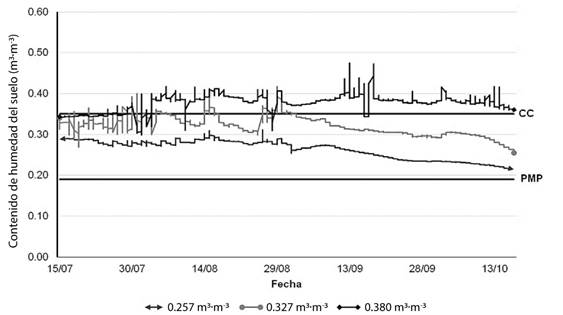
Figura 1 Dinámica de humedad del suelo bajo tres contenidos volumétricos de agua a la profundidad de 80 cm en nogal pecanero (Carya illinoinensis) durante el ciclo productivo 2016. CC: capacidad de campo, PMP: punto de marchitez permanente.
En 2017 se observaron curvas de humedad similares a las del 2016. La precipitación acumulada en 2016 fue 352 mm; la cantidad de agua registrada fue 114 mm de enero a junio y 238 mm de julio a octubre. En el 2017, la cantidad de agua acumulada fue 200 mm, muy inferior a la del 2016, obteniendo un registro de 26 mm en el periodo enero a junio y 131 mm de julio a septiembre, mientras que el resto, equivalente a 43 mm, se registró en diciembre.
Rendimiento de nuez
De acuerdo con el Cuadro 1, los tratamientos con mayor humedad incrementaron la producción de nuez por árbol (P ≤ 0.05). Este resultado se debió principalmente al incremento en el tamaño de la nuez y mayor porcentaje de almendra, pero no por el número de nueces. En 2016, considerado año productivo, el incremento fue 12.7 % cuando el nivel de humedad en el suelo aumentó de 0.257 a 0.327 m3∙m-3, y 24.1 % cuando aumentó a 0.380 m3∙m-3. Una respuesta similar se obtuvo en 2017, en el cual se observó menor producción debido a la alternancia que es característica en esta especie. La producción por árbol incrementó 12.7 % cuando la humedad disponible en el suelo aumentó de 0.257 a 0.327 m3∙m-3, y 23.6 % en los árboles con la mayor humedad disponible.
Cuadro 1 Producción total de nuez comercial por árbol (Carya illinoinensis), bajo tres contenidos volumétricos de agua, durante dos ciclos productivos.
| Humedad edáfica (m3∙m-3) | Producción total (kg∙árbol-1) | |
|---|---|---|
| 2016 | 2017 | |
| 0.38 | 21.46 ± 2.57 a | 19.83 ± 1.67 a |
| 0.327 | 19.48 ± 2.95 ab | 18.08 ± 2.63 ab |
| 0.257 | 17.29 ± 6.72 b | 16.04 ± 3.29 b |
± Desviación estándar de la media (n = 3). En cada ciclo, los valores medios seguidas de la misma letra no son significativamente diferentes entre sí, según la prueba de Tukey (P = 0.05).
Germinación prematura de nuez
En los dos ciclos productivos (2016 y 2017), la producción de nuez germinada prematuramente en los árboles (Figura 2) incrementó significativamente (P < 0.05) bajo los contenidos de humedad de 0.327 m3∙m-3 y 0.380 m3∙m-3. Acorde con el Cuadro 2, en el 2016, los árboles con mayor humedad en el suelo tuvieron 12 % más producción de nuez germinada que los árboles con el nivel más bajo. En el 2017, a partir de la etapa del endurecimiento de cáscara (finales de julio) hasta el final del ciclo productivo, el mayor nivel de humedad en el suelo provocó el aumento significativo (P < 0.05) de 10.5 % de nuez germinada. El incremento en viviparidad afectó negativamente el porcentaje de nuez comercial (Cuadro 2); por lo tanto, el nivel de humedad en el suelo de 0.257 m3∙m-3 resultó ser el más efectivo en la disminución de la viviparidad sin afectar el porcentaje de nuez comercial.

Figura 2 Nuez germinada dentro del ruezno de Carya illinoinensis mostrando la radícula durante el periodo de maduración (A) y nuez cosechada (B).
Cuadro 2 Nuez germinada prematuramente y nuez comercial de Carya illinoinensis en función de la humedad disponible en el suelo durante dos ciclos de producción.
| Humedad edáfica (m3∙m-3) | Año 2016 | Año 2017 | ||
|---|---|---|---|---|
| Nuez germinada (%) | Nuez comercial (%) | Nuez germinada (%) | Nuez comercial (%) | |
| 0.38 | 17.4 ± 3.3 a | 82.6 ± 3.8 a | 14.6 ± 1.4 a | 85.4 ± 4.7 a |
| 0.327 | 14.2 ± 0.9 a | 85.8 ± 4.1 a | 12.8 ± 3.2 a | 87.2 ± 5.9 a |
| 0.257 | 5.2 ± 1.4 b | 94.8 ± 6.9 b | 4.1 ± 2.3 b | 95.9 ± 7.6 b |
| CV (%) | 8.7 | 20.2 | 7.2 | 17.4 |
± Desviación estándar de la media (n = 3). En cada columna, los valores medios seguidos por las mismas letras no son significativamente diferentes entre sí, según la prueba de Tukey (P = 0.05).
Tamaño del fruto
En el ciclo productivo 2016, la longitud y el diámetro del fruto incrementaron significativamente (P < 0.05) en los árboles con los dos niveles de humedad superiores (Cuadro 3). En este ciclo, año de mayor producción, la longitud y el diámetro de nuez en los árboles con mayor contenido de humedad (0.38 m3 ∙m-3) fueron 30 % y 12 % mayores que en los árboles sometidos a niveles menores de humedad. En 2017 se observó un efecto similar; el tratamiento con menor contenido de humedad disponible en el suelo (0.257 m3 ∙m-3) ocasionó una reducción de 1.0 cm (20.6 %) y 0.23 cm (8 %) en la longitud y diámetro del fruto, respectivamente, con respecto al nivel alto de humedad. A partir del endurecimiento del fruto, la longitud y diámetro final al momento de la cosecha se redujo en los árboles con suministro hídrico menor.
Cuadro 3 Longitud y diámetro de nuez pecanera (Carya illinoinensis) en función de tres contenidos de humedad del suelo durante dos ciclos productivos.
| Humedad edáfica (m3∙m-3) | Año 2016 | Año 2017 | ||
|---|---|---|---|---|
| Longitud de fruto(cm) | Diámetro de fruto(cm) | Longitud de fruto(cm) | Diámetro de fruto(cm) | |
| 0.38 | 5.07 ± 0.22 a | 2.90 ± 0.24 a | 5.17 ± 0.18 a | 2.90 ± 0.19 a |
| 0.327 | 4.93 ± 0.24 a | 2.81 ± 0.26 a | 4.95 ± 0.2 a | 2.85 ± 0.17 a |
| 0.257 | 3.89 ± 0.27 b | 2.58 ± 0.18 b | 4.16 ± 0.32 b | 2.67 ± 0.22 b |
| CV (%) | 4.69 | 7.54 | 8.54 | 5.61 |
± Desviación estándar de la media (n = 50). En cada columna, los valores medios seguidos por las mismas letras no son significativamente diferentes entre sí, según la prueba de Tukey (P = 0.05).
Porcentaje de almendra
El Cuadro 4 indica que, en el 2016, el porcentaje de almendra aumentó significativamente (P < 0.05) en los tratamientos con mayor humedad disponible en el suelo; los porcentajes de almendra fueron 1.95 y 0.62 % mayores que en los nogales sometidos a niveles inferiores de humedad. En el 2017, los tratamientos de humedad tuvieron un efecto similar sobre esta variable; en el nivel de humedad del suelo de 0.257 m3∙m-3 se observó el menor porcentaje de almendra, el cual fue superado en 0.58 y 1.82 puntos porcentuales por los árboles con niveles de humedad de 0.327 y 0.380 m3∙m-3, respectivamente.
Cuadro 4 Porcentaje de almendra bajo tres niveles de humedad en el suelo en dos ciclos de producción de nogal pecanero (Carya illinoinensis).
| Humedad edáfica (m3∙m-3) | Año 2016 | Año 2017 |
|---|---|---|
| Almendra (%) | Almendra (%) | |
| 0.38 | 58.85 ± 3.50 a | 58.82 ± 3.33 a |
| 0.327 | 57.52 ± 1.50 ab | 57.58 ± 2.87 ab |
| 0.257 | 56.90 ± 1.28 b | 57.00 ± 2.68 b |
± Desviación estándar de la media (n = 3). En cada columna, los valores medios seguidos por las mismas letras no son significativamente diferentes entre sí, según la prueba de Tukey (P = 0.05).
Discusión
Rendimiento de nuez
Las condiciones ambientales favorables y un manejo adecuado durante el ciclo productivo del nogal, particularmente en cada fase fenológica, propician una buena producción (Reyes Vázquez & Morales Landa, 2019). Asimismo, la humedad disponible en el suelo (Grauke, Wood, & Harris, 2016; Thompson, 2005), asociada con el suministro de fertilizaciones, es importante para que el árbol manifieste su potencial productivo (Walworth, White, & Comeau, 2017). Por ello, es fundamental evitar condiciones de estrés hídrico, durante el periodo de crecimiento y llenado de la nuez (Godoy & López, 2000). Los resultados de los dos años de evaluación indican que el mayor nivel de humedad en el suelo, a partir del endurecimiento de cáscara del fruto ocurrido a finales de julio, tuvo un efecto positivo en la producción de nuez por árbol. Esta superó la producción media de nuez (1.1 t∙ha-1) en la Comarca Lagunera, reportada por Santamaría, Medina Morales, Rivera-González, y Faz Contreras (2002), particularmente en los dos tratamientos con mayor contenido de humedad. El incremento del rendimiento de nuez, al aumentar la humedad en el suelo, también ha sido observado por Marco et al. (2021) y soporta lo establecido por Sparks (2005) con relación a la necesidad hídrica elevada de la especie (Walls, 2017). Se deduce que el requerimiento hídrico elevado fue heredado por sus ancestros, los cuales se desarrollaron en los márgenes de ríos y suelos profundos y fértiles (Babuin, Echeverría, Menedez, & Maiale 2016; Poletto, Poletto, Morales, Briao, Silveira, & Richards 2020). Al respecto, Orona Castillo, Sangerman-Jarquín, Cervantes Vázquez, Espinoza Arellano, y Núñez Moreno (2019) determinaron que los rendimientos de nuez por unidad de superficie, en las huertas de nogales del norte de México, están directamente relacionados con la disponibilidad de riegos y la atención técnica. Considerando lo anterior, se puede explicar la baja producción en los árboles que tuvieron menor volumen de agua en el suelo, reflejada en la menor cantidad de frutos producidos y de menor tamaño, como se ha observado en otras especies (Wiegand & Swanson, 1982).
Germinación prematura de nuez
Diversos estudios indican que existe una serie de plantas que presentan viviparidad (Duermeyer et al., 2018); sin embargo, son pocos los reportes que se tienen sobre este fenómeno en las especies frutícolas comerciales (Taylor, Kunene, & Pandor, 2020). Farnsworth (2000) plantea que la viviparidad es más común en plantas con un perfil morfológico particular, que incluye semillas grandes observadas solo en una fruta; semillas con endocarpio duro rodeado de tejido carnoso que mantiene el contenido de agua interno; especies de dispersión dirigida (estrategia que les permite alcanzar hábitats específicos favorables para su supervivencia); especies con larga vida adulta, así como, aquellas que están menos especializadas para microhábitats; y las adaptadas a los hábitats húmedos. Según Wood (2015), la viviparidad también es favorecida por una humedad elevada en el fruto, aunada a la presencia de temperaturas de 30 a 35 °C durante la etapa final de su desarrollo y baja concentración de ácido abscísico. Dado que la nuez pecanera presenta la mayoría de estas características, la probabilidad de que presente germinación prematura, aun permaneciendo en el árbol, es elevada, cuando las condiciones de temperatura y humedad son propicias (Gonçalves Bilharva et al., 2018; Wood, 2015). La región de estudio presenta temperatura máxima promedio de 33 °C y precipitación de 131 mm de julio a septiembre, condiciones que pueden ser propicias para que ocurra este fenómeno (Sifuentes-Ibarra et al., 2015).
La viviparidad es considerada una adaptación de la planta para tomar ventaja ante ciertas condiciones y así incrementar la sobrevivencia en el medio silvestre; en nogales cultivados, es una causa de pérdidas económicas (Wood, 2015). El proceso de la germinación usa carbohidratos almacenados para el crecimiento, y en el proceso causa ennegrecimiento de la región del embrión que une a las dos secciones de almendra; las nueces con esta característica no son comercializables (Wood, 2015). Taylor et al. (2020) agregan que, además de las condiciones mencionadas, el vigor de los árboles, la condición de luz y la fertilización en exceso en las últimas etapas del desarrollo de la nuez, podrían ser los factores que desencadenan la viviparidad en C. illinoinensis.
En el presente estudio, la mayor cantidad de nueces germinadas se observó en los árboles con mayor contenido de humedad en el suelo. La tendencia hacia mayor viviparidad, conforme el nivel de humedad incrementó en el suelo, confirma los resultados obtenidos por Godoy y López (2000), León (2014), y Wood (2015). Previo a la maduración, las nueces no están fisiológicamente en dormancia, sino que tienen una restricción mecánica para germinar (Wood, 2015). En esta restricción, la cáscara de la nuez, que parece ser una barrera formidable (Wells, 2017), no impide la transferencia de agua o gases requeridos para la germinación. Como resultado, la cantidad elevada de humedad entre la cáscara y el ruezno de la nuez, asociadas con temperaturas altas, podrían consecuentemente ser un factor que neutraliza el efecto mecánico de la cáscara, permitiendo que la nuez germine prematuramente, aun estando en el árbol, como sucede en algunas variedades susceptibles bajo condiciones climáticas adecuadas (Aguilar et al., 2015). En 2016, las temperaturas máxima, mínima y media en septiembre fueron 32.7, 19.3 y 26 °C, respectivamente, y en octubre fueron 20.2, 16.2 y 23.3 °C. En 2017, las temperaturas máxima, mínima y media en septiembre fueron 32.4, 19.0 y 25.7 °C y en octubre fueron 30.2, 16.9 y 23.6 °C. La presencia de temperaturas superiores a los 30 °C durante el periodo de maduración de la nuez, asociadas con la humedad elevada, pudieron ser un factor importante sobre la inducción de la germinación prematura de nueces en el árbol (García-Moreno et al., 2020; Nonogaki, Barrero, & Li, 2018).
Por otra parte, la cantidad de nueces germinadas en el árbol incrementa en los años de alta producción (Sparks, 2005; Stein, 1985). En este estudio, la tendencia hacia una mayor presencia de nueces germinadas en 2016 (año de mayor producción) no se manifestó en el 2017, año con menor producción y viviparidad.
Tamaño de nuez y porcentaje de almendra
El crecimiento y llenado de la nuez dependen fundamentalmente del contenido de humedad; por lo tanto, es importante su disponibilidad adecuada en el suelo, tanto en la primera fase de desarrollo, que se caracteriza por el crecimiento de la nuez, como durante la segunda fase que corresponde al llenado (Ferreyra, Selles, & Lemus, 2002; Godoy & Huitrón, 1998; Godoy & López, 2000). El estrés de humedad durante el crecimiento del fruto conduce a la producción de nueces pequeñas (Godoy & Huitrón, 1998; Herrera, 1990). Por el contrario, un déficit hídrico en el llenado del fruto induce un bajo porcentaje de almendra (Herrera, 1990; Godoy & Huitrón, 1998). En el presente estudio, el tamaño de la nuez incrementó a medida que la disposición de humedad aumentó, sin demeritar el porcentaje de almendra, cuya tendencia hacia el incremento fue mayor en árboles con mejor condición de humedad. Los valores de crecimiento de la nuez son similares a los reportados por Marco et al. (2021), coincidiendo en que la cantidad de agua aplicada durante el llenado de fruto influye en el ancho y longitud, obteniendo nueces significativamente más grandes conforme el riego aumenta.
Según el USDA (United States Department of Agriculture, 2020), la longitud promedio de las nueces es de 36.8 mm y el ancho de 25.07 mm. En este estudio, los valores de longitud y ancho de nuez fueron mayores que los referidos e incrementaron en relación con la disponibilidad hídrica. En este sentido, durante el periodo del llenado de almendra cuando ocurre el desarrollo del embrión, es evidente la magnitud de la respuesta de nogal ante el aumento del agua de riego; sin embargo, la viviparidad aumenta y el porcentaje de nuez comercial disminuye. La respuesta favorable hacia el tamaño mayor y porcentaje elevado de almendra en los tratamientos de mayor humedad no coincide con el supuesto de que, en el nogal, las nueces pequeñas llenan mejor o producen almendra mejor desarrollada (Thompson, 2005). En este estudio, el porcentaje de almendra varió de 56 % a 58 % en los árboles con menor y mayor humedad durante los dos ciclos productivos. Estos registros superan los valores de referencia para exportación (55 %) (Godoy & López, 2000). El porcentaje de almendra es usado en la industria nogalera para determinar el precio por unidad de peso de la nuez en cáscara (Thompson, 2005). Igualmente, se ha observado que los rendimientos elevados afectan el porcentaje de almendra (Smith, 2012; Smith, Reid, Carroll, & Cheary, 1993). En el presente trabajo, el rendimiento por árbol, particularmente en el año 2016, no afectó el tamaño y el porcentaje de almendra, los cuales incrementaron dentro de los niveles de humedad evaluados.
Conclusiones
El contenido de humedad de 0.257 m3∙m-3 de suelo fue el mejor tratamiento para reducir la germinación prematura de nuez en el árbol. La mayor producción de nuez por árbol se obtuvo con el tratamiento de 0.380 m3∙m-3; sin embargo, la viviparidad también incrementó y afectó el porcentaje de nuez comercial; el riego aplicado por el productor (0.327 m3∙m-3) tuvo el mismo efecto en dichas variables. Por tanto, en este estudio se comprobó que la humedad elevada del suelo induce la viviparidad y que la disminución de riego podría reducir dicho fenómeno.
References
Aguilar-Pérez, H., Arreola-Ávila, J. G., Morales-Olais, E., Cuéllar-Villarreal, E., Lagarda-Murrieta, A., Tarango-Rivero, H., …Lombardini, L. (2015). ‘Norteña’ Pecan. HortScience, 50(9), 1399-1400. doi: 10.21273/HORTSCI.50.9.1399 [ Links ]
Babuin, M. F., Echeverría, M., Menedez, A. B., & Maiale, S. J. (2016). Arbuscular mycorryhizal pecan seedlings alleviate effect of restricted water suly. HortScience, 5(3), 212-215. doi: 10.21273/HORTSCI.51.3.212 [ Links ]
Cohen, M., Valancogne, C., Dayau, S., Ameglio, T., Cruiziant., P., & Archer, P. (1997). Yield and physiological responses of walnut trees in semiarid conditions: application to irrigation scheduling. Acta de Horticultura, 449, 273-280. doi: 10.17660/ActaHortic.1997.449.39 [ Links ]
Duermeyer, L., Khodaanahi, E., Yan, D., Krapp, A., Rothstein, S. J., & Nambara, E. (2018). Regulation of seed dormancy and germination by nitrate. Seed Science Research, 28(3), 150-157. doi: 10.1017/S096025851800020X [ Links ]
Farnsworth, E. (2000). The ecology and physiology of viviparous and recalcitrant seeds. Annual Review of Ecology and Systematics, 31, 107-138. doi: 10.1146/annurev.ecolsys.31.1.107 [ Links ]
Ferreyra, E. R., Selles, V. G., & Lemus, S. G. (2002). Efecto del estrés hídrico durante la fase II de crecimiento del fruto del duraznero cv. Kakamas en el rendimiento y estado hídrico de las plantas. Agricultura Técnica, 62(4), 565-573. doi: 10.4067/S0365-28072002000400008 [ Links ]
García-Moreno, B. Y., Báez-Sañudo, R., Mercado-Ruiz, J. N., García-Robles, J. M., & Núñez-Moreno, J. H. (2020). Bioregulación de la germinación prematura de nuez pecanera mediante aplicaciones precosecha con ácido 2-hidroxibenzoico. Revista Iberoamericana de Tecnología Postcosecha, 21(2), 1-12. Retrieved from http://www.redalyc.org/articulo.oa?id=81365122005 [ Links ]
Garrot, D., Kilbby, M., Fangmeier, D., Husman, S., & Ralowics, A. (1993). Production, growth, and nut quality under water stress based on crop water stress index. Journal American Society for Horticultural Science, 118(6), 694-698. doi: 10.21273/JASHS.118.6.694 [ Links ]
Grauke, L. J., Wood, B. W., & Harris, M. K. (2016). Crop vulnerability: Carya. HortScience, 51(6), 653-663. doi: 10.21273/HORTSCI.51.6.653 [ Links ]
Godoy, A. C., & Huitrón, R. M. (1998). Relaciones hídricas de hojas y frutos de nogal pecanero durante el crecimiento y desarrollo de la nuez. Agrociencia, 32(4), 331-337. Retrieved from https://www.agrociencia-colpos.mx/index.php/agrociencia/article/view/1564 [ Links ]
Godoy Avila, C., & López Montoya, I. (2000). Desarrollo de la almendra y germinación del fruto del nogal pecanero bajo cuatro calendarios de riego. Terra Latinoamericana, 18(4), 305-311. Retrieved from https://www.redalyc.org/pdf/573/57318404.pdf [ Links ]
Gonçalves Bilharva, M., Roberto Martins, C., Janer Hamann, J., Fronza, D., De Marco, R. and Barbosa Malgarim, M. (2018). Pecan: from research to the Brazilian reality. Journal of Experimental Agriculture International, 23(6), 1-16. doi: 10.9734/JEAI/2018/41899 [ Links ]
Herrera, E. (1990). Fruit growth and development of Ideal and Western pecans. Journal of the American Society for Horticultural Science, 115(6), 915-923. [ Links ]
León, G. V. (2014). Ácido abscísico-giberelinas como indicador de viviparidad en nogal pecanero (Carya illinoinensis K.). Hermosillo, Sonora, México: CIAD. [ Links ]
Marco, R. D., Goldschmidt, R. J., Herter, F. G., Martins, C. R., Mello-Farias, P. C., & Uberti, A. (2021). The irrigation effect on nuts’ growth and yield of Carya illinoinensis. Anais da Academia Brasileira de Ciencias, 93(1), 1-8. doi: 10.1590/0001-3765202120181351 [ Links ]
Nonogaki, H., Barrero, J. M., & Li, C. (2018). Seed dormancy, germination, and pre-harvest sprouting. Frontiers in Plant Science, 9, 1783. doi: 10.3389/fpls.2018.01783 [ Links ]
Orona Castillo, I., Sangerman-Jarquín, D. M., Cervantes Vázquez, M. G., Espinoza Arellano, J. de J., & Núñez Moreno, J. H. (2019). La producción y comercialización de nuez pecanera en México. Revista Mexicana de Ciencias Agrícolas, 10(8), 1797-1808. doi: 10.29312/remexca.v10i8.1833 [ Links ]
Ou, S. K., Storey, J. B., & Thompson, T. E. (1994). A northern pecan pollen source delays germination of nuts from a southern pecan cultivar. HortScience, 29(11), 1290-1291. doi: 10.21273/HORTSCI.29.11.1290 [ Links ]
Poletto, T., Poletto, I., Moraes Silva, L. M., Briao Muñiz, M. F., Silveira Reiniger, L. R., Richards, N., & Marco, S. V. (2020). Morphological, chemical and genetic analysis of southern Brazilian pecan (Carya illinoinensis) accessions. Acta Horticulturae, 261, 1-7 doi: 10.1016/j.scienta.2019.108863 [ Links ]
Prodan, M. (1968). Forest bimetrics. Oxford, Inglaterra: Pergamon Pres. [ Links ]
Reyes Vázquez, N. C., & Morales Landa, J. L. (2019). Agronomía sustentable y aprovechamiento alternativo de la nuez. Agronomía sustentable y aprovechamiento alternativo de la nuez. México: Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco CIATEJ. Retrieved from http://ciatej.repositorioinstitucional.mx/jspui/handle/1023/671 [ Links ]
Santamaría, J., Medina Morales, M. C., Rivera-González, M., & Faz Contreras, R. (2002). Algunos factores de suelo, agua y planta que afectan la producción y alternancia del nogal pecanero. Revista Fitotecnia Mexicana, 25(2), 119-125. Retrieved from https://www.revistafitotecniamexicana.org/documentos/25-2/1a.pdf [ Links ]
SAS Institute. (2002). Statistical analysis system. The SAS system for Windows version 9.0. Cary, North Carolina, USA: Author. [ Links ]
Servicio Meteorológico Nacional (SMN). (2010). Normales climatológicas por estado. Retrieved from https://smn.conagua.gob.mx/es/climatologia/informacion-climatologica/normales-climatologicas-por-estado [ Links ]
Sifuentes-Ibarra, E., Samaniego-Gaxiola, J. A., Anaya-Salgado, A., Núñez-Moreno, J. H., Valdez-Gascón, B., Gutiérrez-Soto, R. G., … Macías-Cervantes, J. (2015). Programación del riego en nogal pecanero (Carya illinoinensis), mediante un modelo integral basado en tiempo térmico. Revista Mexicana de Ciencias Agrícolas, 6(8), 1893-1902. doi: 10.29312/remexca.v6i8.527 [ Links ]
Smith, M. W., Reid, W., Carroll, B., & Cheary, B. (1993). Mechanical fruit thinning influences fruit quality, yield, return fruit set, and cold injury of pecan. HortScience, 28(11), 1081-1084. doi: 10.21273/HORTSCI.28.11.1081 [ Links ]
Smith, M. W. (2012). Fruit production characteristics in ‘Pawnee’ Pecan. HortScience, 47(4), 489-496. doi: 10.21273/HORTSCI.47.4.489 [ Links ]
Sparks, D. (2005). Adaptability of pecan as a species. HortScience, 40(5), 1175-1189. doi: 10.21273/HORTSCI.40.5.1175 [ Links ]
Sparks, D., Reid, W., Yates, I., Smith, M. W., & Stevenson, T. G. (1995). Fruiting stress induces shuck decline and premature germination in pecan. Journal of the American Society for Horticultural Science, 120(1), 43-53. doi: 10.21273/JASHS.120.1.43 [ Links ]
Stein, L. A. (1985). The influence of fertilization, phloem restriction, irrigation, microbes, and ethephon on pecan shuck disorders in Texas (Ph. D. Diss.), Texas A&M University, Texas, USA. [ Links ]
Taylor, N., Kunene, S., & Pandor, M. (2020). Stick-tights and vivipary in pecans. Retrieved from https://www.sappa.za.org/wp-content/uploads/docs/2020/07/STICK-TIGHTS-AND-VIVIPARY-IN-PECANS_v3.pdf [ Links ]
Thompson, T. E. (2005). Pecan fruit shuck thickness is related to nut quality. HortScience, 40(6), 1664-1666. doi: 10.21273/HORTSCI.40.6.1664 [ Links ]
United States Department of Agriculture (USDA). (2020). NCGR Pecan/Hickory Database (Pecan Cultivars). Retrieved from https://aggiehorticulture.tamu.edu/usda_pecan/plant_details.php?pid=Success&acno=518130 [ Links ]
Walworth, J. L., White, S. A., & Comeau, M. (2017). Soil - applied Zn EDTA: vegetative growth, nut production, and nutrient acquisition of immature pecan trees grown in alkaline calcareous soils. HortScience, 2(2), 300-305. doi: 10.21273/HORTSCI11467-16 [ Links ]
Wells, L. (2017). Southeastern pecan growers' handbook. Georgia, USA: Cooperative Extension Service, University of Georgia College of Agricultural & Environmental Sciences. [ Links ]
White, C. N., Proebsting, W. M., Hedden, P., & Rivin, C. (2000). Gibberellins and seed development in maize. I. Evidence that gibberellin/abscisic acid balance governs germination versus maturation pathways. Plant Physiology, 122(4), 1081-1088. doi: 10.1104/pp.122.4.1081 [ Links ]
Wiegand, C., & Swanson, W. (1982). Citrus responses to irrigation: II Fruit yield, size and number. Journal of the Rio Grande Valley Horticultural Society, 35, 87-95. [ Links ]
Wood, B. (2015). Regulation of vivipary in pecan. Acta Horticulturae, 1070, 33-42. doi: 10.17660/ActaHortic.2015.1070.3 [ Links ]
Recibido: 11 de Julio de 2021; Aprobado: 09 de Diciembre de 2021
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
