










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.27 no.2 Chapingo may./ago. 2021 Epub 26-Ene-2024
https://doi.org/10.5154/r.rchscfa.2020.06.046
Artículo científico
Injerto interespecífico como alternativa para la propagación asexual de Pinus rzedowskii Madrigal & Caball. Del. en riesgo de extinción
1 Universidad Michoacana de San Nicolás de Hidalgo (UMSNH), Facultad de Agrobiología “Presidente Juárez”. Av. Revolución esquina con Berlín s/n, col. Viveros. C. P. 60170. Uruapan, Michoacán, México.
2 Universidad Veracruzana, Instituto de Investigaciones Forestales. Parque Ecológico “El Haya”, Carretera antigua a Coatepec s/n. C. P. 91070. Xalapa, Veracruz, México.
Introducción:
Pinus rzedowskii Madrigal & Caball. Del. es endémica de México, en peligro de extinción y con baja reproducción natural. Una alternativa para su propagación es el injerto interespecífico, bajo la premisa de que las especies filogenéticamente más relacionadas influyen positivamente en la supervivencia.
Objetivos:
Evaluar la supervivencia de injertos de P. rzedowskii con plantas-patrón de cinco especies de Pinus.
Materiales y métodos:
Se realizaron injertos interespecíficos de P. rzedowskii con P. pinceana Gordon & Glend., P. maximartinezii Rzed., P. ayacahuite var. veitchii (Roezl) Shaw, P. pseudostrobus Lindl. y P. rzedowskii (testigo). El método de injerto fue el enchapado lateral; el prendimiento se cuantificó durante seis meses y la supervivencia se evaluó con el método no paramétrico de Kaplan-Meier.
Resultados y discusión:
La supervivencia final más alta de los injertos y la función esperada de supervivencia se obtuvieron con plantas-patrón de P. rzedowskii (85.7 ± 0.21 %; m = 0.97), seguido de P. pinceana (80 ± 0.31 %; m = 0.95); P. pseudostrobus obtuvo los valores más bajos (20 ± 0.89 %; m = 0.66). Se obtuvieron diferencias significativas (P < 0.0001) entre P. pseudostrobus y P. maximartinezii con relación a P. rzedowskii y P. pinceana.
Conclusión:
P. pinceana, especie filogenéticamente más cercana con P. rzedowskii y P. maximartinezii, fue la de mayor supervivencia, por lo que puede ser una alternativa para el rescate y conservación ex situ de P. rzedowskii.
Palabras clave: supervivencia; relaciones filogenéticas; enchapado lateral; prendimiento de injerto; Pinus pinceana
Introduction
: Pinus rzedowskii Madrigal & Caball. Del. is endemic to Mexico, in danger of extinction and low natural reproduction. An alternative for propagation is interspecific grafting, under the assumption that the phylogenetically more related species positively influence survival.
Materials and methods:
Interspecific grafting of P. rzedowskii with P. pinceana Gordon & Glend, P. maximartinezii Rzed., P. ayacahuite var. veitchii (Roezl) Shaw, P. pseudostrobus Lindl. and P. rzedowskii (control treatment) was carried out. The grafting method was side-veneer graft; grafting was quantified for six months and survival was evaluated using the nonparametric Kaplan-Meier method.
Results and discussion:
The highest final grafting survival and expected survival function were obtained with rootstocks of P. rzedowskii (85.7 ± 0.21 %; m = 0.97), followed by P. pinceana (80 ± 0.31 %; m = 0.95); P. pseudostrobus had the lowest values (20 ± 0.89 %; m = 0.66). Significant differences (P < 0.0001) were observed between P. pseudostrobus and P. maximartinezii regarding P. rzedowskii a P. pinceana.
Conclusion:
P. pinceana, a species phylogenetically closer to P. rzedowskii and P. maximartinezii, was the one with the highest survival rate, which may be an alternative for the rescue and ex situ conservation of P. rzedowskii.
Keywords: survival; phylogenetic relationships; side-veneer grafting; grafting success; Pinus pinceana
Ideas destacadas
Injertos interespecíficos de P. rzedowskii con P. pinceana, P. maximartinezii, P. ayacahuite y P. pseudostrobus.
La supervivencia se evaluó con el método no paramétrico de Kaplan-Meier.
P. pinceana, especie emparentada con P. rzedowskii, fue la de mayor supervivencia (80 ± 0.3 %).
P. pseudostrobus obtuvo la función de supervivencia más baja (m = 0.66; 20 ± 0.8 %).
Las relaciones filogenéticas deben considerarse para el éxito de injertos interespecíficos de Pinus.
Introducción
El injerto es uno de los métodos de propagación asexual más utilizados en especies de pino (Aparicio-Rentería, Viveros-Viveros, & Rebolledo-Camacho 2013; Muñoz-Flores, Prieto-Ruiz, Flores-García, Pineda-Ojeda, & Morales-González, 2013; Pérez-Luna et al., 2019). Este método acelera el ciclo reproductivo con el fin de obtener mayor producción de planta con mejor calidad genética en menor tiempo; la planta es utilizada para las actividades de reforestación y conservación, establecimiento de huertos semilleros clonales o desarrollo de plantaciones comerciales (Aparicio-Rentería et al., 2013; Guadaño & Mutke, 2016; Muñoz-Flores et al., 2013; Pérez-Luna et al., 2019, 2020a). No obstante, la mayoría de los ensayos se han realizado con tutores de la misma especie y los estudios reportados entre especies diferentes son escasos (Climent, Prada, Gil, & Pardos, 1997; Darikova, Vaganov, Kuznetsova, & Grachev, 2013; Villaseñor & Carrera, 1980; Pérez-Luna et al., 2020b).
Algunas de las ventajas más destacadas del injerto interespecífico son: la capacidad de adaptación en ambientes diferentes al de la especie original y la producción de mayor cantidad de estructuras reproductivas con incremento en la disponibilidad de semillas (Climent et al., 1997). Estas ventajas se han observado en especies de distribución amplia y de importancia económica; en contraste, los ensayos en pinos de distribución restringida y en peligro de extinción son nulos. La aplicación de injertos en este tipo de especies puede ser una herramienta de gran valor para su conservación, ya que mitiga la degradación de sus áreas naturales a través de la propagación para ayudar a su repoblación y reduce la pérdida de variantes genéticas que son de gran importancia adaptativa y evolutiva para el género.
Pinus rzedowskii Madrigal & Caball. Del. es endémica de México y está en peligro de extinción (Secretaría de Medio Ambiente y Recursos Naturales [SEMARNAT], 2010). La especie se distribuye en la Sierra de Coalcomán, Michoacán, con poblaciones fragmentadas (15) y representadas por pocos individuos (9 a 3 500 por población; Castilleja, Delgado, Sáenz-Romero, & Herrerías, 2016; Delgado, Piñero, Chaos, Pérez‐Nasser, & Alvarez‐Buylla, 1999). Su hábitat es característico de suelos calizos (Cambisol) pobres en nutrientes, con poblaciones conformadas en su mayoría por individuos adultos senescentes, lo cual reduce su capacidad de regeneración (Delgado et al., 1999). No obstante, la variación genética es relativamente alta (>0.275), aunque con niveles importantes de endogamia (F = 0.270; Delgado et al., 1999). Esta condición afecta el éxito reproductivo con una producción escasa de semillas viables (17 % con embrión) y germinación baja (3 %; Castilleja et al., 2016).
Una de las alternativas para el incremento de la reproducción de P. rzedowskii es la implementación de injertos, que desafortunadamente son difíciles de realizar entre individuos de la misma especie, debido a la disponibilidad escasa de semillas viables (Castilleja et al., 2016). Por lo anterior, es necesario explorar la posibilidad del uso de otras especies-patrón de pino, bajo el supuesto de que la selección de especies filogenéticamente cercanas, donde las características anatómicas probablemente han evolucionado de manera similar, permitirá mejor acoplamiento de los tejidos y, en consecuencia, mayor éxito de los injertos. Este supuesto filogenético no se ha documentado en pinos, pero se han explorado otros factores, aunque también de manera escasa, como son: el estado fenológico y viabilidad de la vareta y el patrón, la época del injertado, la habilidad del injertador, el control de las condiciones ambientales y la compatibilidad taxonómica y anatómica de las especies (Climent et al., 1997; Hartmann, Kester, Davies, & Geneve, 2002; Pérez-Luna et al., 2019, 2020a, 2020 b).
Las reconstrucciones filogenéticas con datos moleculares de regiones de ADN de cloroplasto y núcleo (Gernandt, Geada, Ortiz, & Liston, 2005; Montes et al., 2019) ubican a P. rzedowskii dentro de la sección Parrya, subsección Cembroides, cuyas relaciones filogenéticas con P. pinceana Gordon & Glend. y P. maximartinezii Rzed. forman un clado monofilético. Por ello, el uso de plantas-patrón con estas especies de pino sustentan la posibilidad de obtener injertos compatibles con P. rzedowskii.
En el presente estudio se hicieron injertos interespecíficos de P. rzedowskii con plantas-patrón de P. pinceana, P. maximartinezii, P. ayacahuite var. veitchii (Roezl) Shaw y P. pseudostrobus Lindl. con el objetivo de evaluar la supervivencia. La hipótesis plantea que las especies filogenéticamente más cercanas a P. rzedowskii presentan mayor supervivencia que las especies con relación filogenética más distante.
Materiales y métodos
Material biológico e injertación
Las plantas-patrón se obtuvieron por germinación de semillas de cinco especies: P. rzedowskii; P. maximartinezii, P. pinceana y P. ayacahuite var. veitchii, tres especies filogenéticamente cercanas del subgénero Strobus; así como P. pseudostrobus del subgénero Pinus (Cuadro 1).
Cuadro 1 Procedencias de las semillas utilizadas para la obtención de las plantas-patrón de las especies de Pinus en estudio.
| Especie | Procedencia | Latitud (N) | Longitud (O) | Altitud (m) |
|---|---|---|---|---|
| P. pinceana | Guadalcázar, San Luis Potosí | 22° 40’ 10’’ | 100° 29’ 30’’ | 1 600 |
| P. maximartinezii | Pueblo Viejo, Jalisco | 21° 21’ 15’’ | 103° 21’ 22’’ | 2 380 |
| P. ayacahuite var. veitchii | La Palma, Michoacán | 19° 36’ 39’’ | 102° 20’ 24’’ | 2 150 |
| P. pseudostrobus | San Juan Nuevo Parangaricutiro, Michoacán | 19° 27’ 06’’ | 102° 12’ 57’’ | 2 685 |
| P. rzedowskii | El Varaloso, Michoacán | 18° 40’ 59’’ | 102° 59’ 34’’ | 2 800 |
Las semillas se trataron con hipoclorito de sodio al 0.2 % por 5 min; posteriormente, se enjuagaron con agua corriente y se embebieron en una solución de agua destilada con peróxido de hidrógeno al 2 % por 24 h, seleccionando las semillas con embrión por la técnica de flotación (Castilleja et al., 2016). Finalmente, las semillas se trataron con 3 g∙L-1 de fungicida Captán® por 5 min y se plantaron en charola de plástico transparente (30 x 30 x 5 cm) con sustrato vermiculita al 100 %. Posterior a los 40 días, las plántulas se cambiaron a un sustrato compuesto de 50 % de turba de musgo, 25 % de agrolita, 25 % de vermiculita y 5.0 g∙planta-1 de fertilizante de liberación controlada (Multicote®), en envases de plástico negro de 5 L de capacidad (30 x 20 cm); los riegos se aplicaron cada tercer día. Las plantas-patrón se seleccionaron de tres años acorde con las características recomendadas; edad entre 2 a 4 años, lo más rectas posibles y con diámetro entre 1.5 a 2.5 cm (Muñoz-Flores et al., 2013). Debido a los problemas reproductivos que P. rzedowskii presenta (Castilleja et al., 2016), solo se obtuvieron siete plantas, por ello, el número de plantas para el resto de las especies se aproximó a 10 con la intención de evitar sesgo en los resultados, a causa de las diferencias en el tamaño de muestra.
Las varetas de P. rzedowskii se obtuvieron en la localidad de Chiqueritos, Dos Aguas, municipio de Aguililla, Michoacán (18° 04’ 17’’ N y 102° 59’ 36’’ O). Se recolectaron 70 varetas con yemas terminales de tamaños similares (5 a 8 cm de longitud), provenientes de un árbol adulto reproductivo (140 años; diámetro de 30 cm y altura de 25 m). La recolecta se hizo de la parte media baja del árbol que contenía la mayor cantidad de yemas. El trabajo de recolecta e injerto se realizó en un periodo de 24 h (18 y 19 de febrero de 2017), con la finalidad de que los tejidos continuaran vivos y se lograra un injerto exitoso (Aparicio-Rentería et al., 2013; Muñoz-Flores et al., 2013).
Los injertos se hicieron en un vivero rústico o jardín común con dimensiones de 4 x 8 x 3 m y malla al 50 % de sombra en la Facultad de Agrobiología “Presidente Juárez” de la Universidad Michoacana de San Nicolás de Hidalgo. Las etapas del injertado se muestran en la Figura 1. El método de injerto fue el enchapado lateral, recomendado para especies de pinos (Muñoz-Flores et al., 2013; Pérez-Luna et al., 2019; Villaseñor & Carrera 1980). Durante el experimento, las condiciones de temperatura fueron constantes entre 23 a 24 °C y la humedad relativa varió de 60 a 80 %. Los injertos se mantuvieron cubiertos con bolsa de papel estraza para disminuir la incidencia de luz y bolsa plástica con agua para mantenerlos hidratados (Muñoz-Flores et al., 2013); la bolsa de papel se retiró a los 30 días, momento en que se apreció el prendimiento de los injertos por el crecimiento de las yemas y la bolsa plástica se retiró a los 60 días. Para evitar problemas por hongos se aplicó fungicida Ridomil Gold® (3 g∙L-1) en el follaje y en el área del injerto una vez por semana.
El 20 de mayo del 2017 se realizó la primera poda en el total del follaje y ramas debajo de la zona del injerto. El 3 de junio del 2017 se realizó la segunda poda del follaje y ramas en la zona continua a la parte superior del injerto, sin podar el extremo terminal del patrón. Esta poda se hizo con la finalidad de que la savia no alimentara su yema principal, dándole prioridad al injerto (Goldschmidt, 2014; Muñoz-Flores et al., 2013). El 17 de junio del 2017 se observó crecimiento avanzado de las yemas de los injertos y se optó por la poda total de las plantas-patrón y de su liberación.

Figura 1 Secuencia de las etapas de injertado. a) vareta de Pinus rzedowskii, b) corte en el tallo de la planta-patrón, c) acoplamiento de tamaños, d) injertado de enchapado lateral, e) amarrado con cinta plástica para injerto (marca MD), f) bolsa plástica con agua para evitar deshidratación, g) cubierta de papel estraza para disminuir la incidencia de luz, h) injerto con plantas-patrón de P. ayacahuite var. veitchii, con estróbilos masculinos en desarrollo, i) segunda poda de P. pinceana en la parte central-superior del injerto con el extremo terminal sin podar y estróbilos masculinos, j) injerto de P. rzedowskii con estróbilos masculinos de mayor tamaño en la etapa de liberación del polen, k) tercera poda y liberación del injerto, e l) injerto con acículas y ramillas nuevas de P. rzedowskii.
Obtención de datos y análisis
Los injertos vivos y muertos se cuantificaron cada siete días hasta los seis meses posteriores al injertado (marzo-agosto, 2017). En cada evaluación se asignó un valor de 0 a las plantas con pérdida de la turgencia de las yemas y amarillamiento en la zona del injerto, y 1 a las plantas con yemas vivas (Barchuk & Díaz, 2000; Sigala, González, & Prieto-Ruíz, 2015). Cada especie se consideró como tratamiento (cinco tratamientos) con 10 repeticiones, a excepción de P. rzedowskii (tratamiento testigo), ya que solo se obtuvieron siete plantas.
Se realizaron curvas de supervivencia con el modelo no paramétrico de Kaplan-Meier (Sigala et al., 2015), el cual permite hacer pruebas de significancia estadística sobre la supervivencia de cada tratamiento a través del tiempo; es decir, analiza el tiempo transcurrido para que ocurra un suceso (en este caso la muerte de las plantas) considerando también los datos censurados, los cuales corresponden a los individuos que, hasta el último momento de medición, no habían presentado el suceso (plantas vivas) (García, 2012). Las mediciones realizadas fueron 12 con un intervalo de 15 días durante seis meses. Las diferencias entre tratamientos se evaluaron con la prueba Log-Rank, fundamentada en la comparación de eventos observados versus los esperados a través de la función de supervivencia (Le, 1997) definida como: S (t) = P (T > t); donde, S (t) es la probabilidad de que al menos una muerte ocurra en un tiempo T, tan grande como el tiempo t12. Asimismo, con esta aproximación no paramétrica se obtuvo el valor medio de la función de supervivencia (m) de las muestras de cada tratamiento. Al dar seguimiento a cada muestra que presentó o no el evento (muerte de la planta), se estimó el valor medio de supervivencia en el tiempo, obtenido del número de días (md) transcurridos de las observaciones realizadas en seis meses (180 días). Finalmente se hicieron comparaciones pareadas de Chi-cuadrado (X2), para determinar la contribución de los tratamientos. Los datos se analizaron en la plataforma R (R Development Core Team, 2016) con la paquetería Survival (2.41.3) y Survminer (0.4.0).
Resultados y discusión
El valor más alto de supervivencia final se obtuvo con injertos de plantas-patrón de la misma especie: P. rzedowskii con seis de siete injertos (85.7 ± 0.21 %), seguido de P. pinceana con ocho de 10 (80 ± 0.316 %), ambas con los mayores valores medios de la función esperada de supervivencia (m = 0.97 y m = 0.95) y de los valores medios de supervivencia estimados en número de días (md = 163.4 y md = 162.2) ((Cuadro 2; Figura 2).
Un estudio genómico reciente de la subsección Cembroides indica que P. pinceana es una de las dos especies más cercanas filogenéticamente a P. rzedowskii (Montes et al., 2019); sus características anatómicas y modificaciones a través del tiempo han sido similares probablemente, permitiendo mejor acoplamiento y funcionalidad de los tejidos del cámbium (Castro-Garibay, Villegas-Monter, & López-Upton, 2017; Pérez-Luna et al., 2020a). Se ha documentado que las características del xilema y la forma de los haces vasculares también deben ser similares para que el contacto sea lo más homogéneo posible (Castro-Garibay et al., 2017; Daricova et al., 2013; Pérez-Luna et al., 2019). En este sentido, y de acuerdo con los resultados obtenidos, se recomienda a P. pinceana como planta-patrón para el injerto exitoso con P. rzedowskii.
No obstante que P. pinceana está catalogada en el estatus de protección especial (NOM-59-SEMARNAT-2010), a diferencia de P. rzedowskii, aún no presenta problemas reproductivos extremos con 54 a 55 % de eficiencia de semillas (Quiroz-Vázquez, López-Upton, Cetina-Alcalá, & Ángeles-Pérez, 2017) y algunas de las poblaciones muestran regeneración adecuada, representada por 22 a 59 % de plantas jóvenes (1 a 25 años) con una tasa de reclutamiento superior a un individuo por árbol (Martínez-Ávalos et al., 2015; Molina-Freaner, Delgado, Pérez, Piñero, & Alvarez-Buylla, 2001). Con base en lo anterior, P. pinceana puede ser una fuente de semillas segura para la producción de plantas-patrón para el injerto con P. rzedowskii, sin afectar el equilibrio demográfico y genético de la especie.
Cuadro 2 Función de supervivencia de Kaplan-Meier en injertos de Pinus rzedowskii con cinco tratamientos (especies-patrón) y 12 evaluaciones en intervalos de 15 días durante seis meses.
| Especie | m | DE | IC (95 %) | md | DE | IC (95 %) | ||
|---|---|---|---|---|---|---|---|---|
| Límite inferior | Límite superior | Límite inferior | Límite superior | |||||
| P. pinceana | 0.951 | 0.035 | 0.886 | 1 | 162.203 | 2.962 | 156.291 | 167.616 |
| P. maximartinezii | 0.758 | 0.085 | 0.609 | 0.944 | 130.993 | 6.15 | 116.975 | 145.01 |
| P. ayacahuite var. veitchii | 0.852 | 0.065 | 0.834 | 0.988 | 145.796 | 4.69 | 145.211 | 164.381 |
| P. pseudostrobus | 0.666 | 0.106 | 0.558 | 0.873 | 121.509 | 8.523 | 102.844 | 140.174 |
| P. rzedowskii | 0.973 | 0.027 | 0.922 | 1 | 163.421 | 0 | - | - |
m = valores medios de la función esperada de supervivencia, md = valores medios de supervivencia respecto al número de días, IC = intervalo de confianza (95 %), DE = desviación estándar de la media.
P. maximartinezii, la segunda especie cercanamente emparentada con P. rzedowskii, tuvo una supervivencia final baja con tres de 10 injertos (30 ± 0.91 %) y menor que la esperada de acuerdo con la función de supervivencia (m = 0.758; md = 130.9). Este resultado posiblemente se debió a que las plantas-patrón tenían mayor tamaño (diámetro y altura del tallo) en comparación con las otras especies utilizadas, aunque eran de la misma edad, de acuerdo con lo recomendado (Aparicio-Rentería et al., 2013). Probablemente, debido a las diferencias de crecimiento natural con estructuras anatómicas de tamaño distinto (v. g., haces vasculares de mayor diámetro), no se logró un acoplamiento adecuado de los tejidos. Para corroborar esta aseveración es necesario probar injertos con plantas-patrón menores de tres años y ver si es posible el ajuste de los tejidos que incrementen la tasa de supervivencia. Cabe mencionar que P. maximartinezii está considerada como microendémica y en estatus de protección especial (NOM-59-SEMARNAT-2010). Aunque no presenta problemas de estabilidad demográfica (López-Mata, 2013), su semilla se usa para la producción y comercialización de planta ornamental sin regulación. Debido a lo anterior, la regeneración natural de P. maximartinezii podría verse afectada si las plantas producidas en los viveros no se reintroducen en sus poblaciones, por tanto, el desarrollo de un programa de repoblación para su protección podría ser requerido.
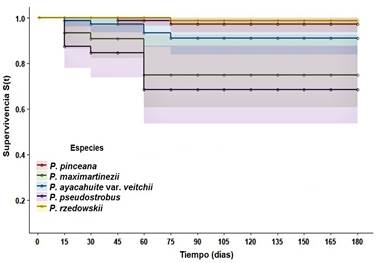
Figura 2 Distribución de la función de supervivencia de Kaplan-Meier para injertos de Pinus rzedowskii con el uso de cinco especies-patrón. Los colores en tonos claros indican los intervalos de confianza (95 %) para cada especie.
Una de las especies promisorias para el éxito de los injertos es P. ayacahuite var. veitchii, que también forma parte del subgénero Strobus con un haz vascular como P. rzedowskii; aunque está agrupada en otra sección (Quinquefoliae) y subsección (Strobus) (Gernandt et al., 2005) se ha observado que los árboles de ambas especies tienen características fisionómicas muy similares cuando son jóvenes (2 a 15 años), por lo que es muy probable obtener buen acoplamiento de los tejidos. Al ser la segunda especie con el mayor porcentaje de supervivencia final (50 ± 0.628) y con un tiempo medio de supervivencia esperada aceptable (m = 0.85, md = 154.7), se propone como una especie-patrón apta para su injerto con P. rzedowskii. Además, la obtención de semillas de P. ayacahuite, para la producción de planta, no afecta el equilibrio demográfico de sus poblaciones naturales, ya que no presenta problemas reproductivos con 54.9 % de eficiencia de semillas y 71.5 % de germinación (Castilleja et al., 2016).
La especie menos indicada fue P. pseudostrobus con supervivencia final (20 ± 0.89 %) y tiempo medio de supervivencia esperada (m = 0.66; md = 121.5) más bajos. Uno de los principales factores que probablemente influyó en este resultado fue su relación filogenética distante con P. rzedowskii, debido a que forma parte de otro subgénero (Pinus) con características morfológicas y anatómicas diferentes que posiblemente impidieron la unión completa de los tejidos (Darikova et al., 2013; Goldschmidt, 2014).
La heterogeneidad observada entre las especies fue altamente significativa con la prueba de Log-Rank (X2 = 25.24, P < 0.0001), la cual se debe a diferencias entre P. rzedowskii con P. pseudostrobus (X2 = 13.480, P < 0.0001) y P. maximartienzii (X2 = 8.390, P < 0.004), y estas dos últimas especies con relación a P. pinceana (P. pseudostrobus, X2 = 15.357, P < 0.0001; P. maximartinezii, X2 = 9.105, P < 0.003) ambas con la menor supervivencia (Figura 2). También se presentaron diferencias entre P. ayacahuite var. veitchii y P. pseudostrobus (X2 = 5.344) con un soporte estadístico menor pero significativo (P < 0.021). Los resultados sugieren nuevamente que las relaciones filogenéticas son un factor importante para el éxito de los injertos, mostrando que la especie más distante (P. pseudostrobus) fue la menos exitosa y que generó la mayor diferenciación.
A la fecha, los estudios sobre injertos heteroespecíficos son escasos y no se ha logrado determinar algún patrón claro de su comportamiento. No obstante, algunos de ellos muestran tendencias similares a los resultados obtenidos en este estudio; el ensayo de P. patula Schiede ex Schltdl. et Cham. en P. radiata D. Don. con 96.7 % de supervivencia (Dyson, 1975), ambas pertenecientes a la subsección Australes (Gernandt et al., 2005); P. patula (subsección Australes) en P. pseudostrobus y P. douglasiana Mtz. (ambas en la subsección Ponderosa) con 80 % y 69 % de supervivencia, respectivamente (Villaseñor & Carrera 1980), las cuales pertenecen a la sección Trifoliae (Gernandt et al., 2005), lo que probablemente posibilitó su supervivencia. Otro estudio es el de P. brutia Ten. en P. nigra Arnold. con 56 % de supervivencia (Climent et al., 1997), ambas de la subsección Pinus (Eckert & Hall, 2006).
Un estudio reciente sobre injertos de P. engelmannii Carr. con la misma especie y con un híbrido de P. engelmanni x P. arizonica Engelm. var. arizonica indicó mayor supervivencia con el híbrido (83 %) que con la especie pura (25 %) (Pérez-Luna et al., 2020b). Los autores concluyen que el bajo prendimiento se debió a la alta densidad de canales resiníferos en las varetas de P. engelmannii (Pérez-Luna et al., 2019); mientras que, para el caso del éxito de los injertos con el híbrido, los autores indican que la presencia de pocos estomas en las acículas de P. arizonica hace que los injertos sean más resistentes al estrés ambiental con un incremento en la supervivencia. Estas diferencias existen a pesar de que ambos taxones están relacionados cercanamente (subsección Ponderosae; Eckert & Hall, 2006), lo que también puede considerarse como otro factor importante para los resultados obtenidos con los injertos de las plantas híbridas.
En un estudio sobre la anatomía de plantas-patrón y varetas para injertos de cuatro especies de pino (P. patula, P. greggii Engelm. ex Parl., P. teocote Schiede ex Schltdl. et Cham. y P. leiophylla Schiede ex Schltdl. et Cham.), Castro-Garibay et al. (2017) demostraron que un cámbium circular y continuo es el más adecuado para una conexión completa de los tejidos del injerto; dichos autores observaron estas características en P. teocote y P. leiophylla, consideradas como especies-patrón candidatas para el injerto del resto de las especies que evaluaron. En el mismo trabajo, los resultados anatómicos sugirieron la influencia de la cercanía filogenética, donde las especies más promisorias (P. teocote y P. leiophylla) fueron incluidas en un estudio filogenético de Pinus (Gernandt et al., 2005), dentro de la subsección Australes y todas (las cuatro especies) dentro de la sección Trifoliate.
En general, en los estudios mencionados se observa que la proximidad filogenética es un factor común para el éxito de los injertos. Por tanto, la heterogeneidad de los resultados obtenidos en este trabajo está asociada claramente a las diferencias filogenéticas y evolutivas de las especies. Actualmente, no existe información sobre la anatomía de los tallos de las especies estudiadas; sin embargo, algunos trabajos taxonómicos reportan diferencias anatómicas en acículas (Farjon & Styles, 1997; Madrigal & Caballero, 1969; Martínez, 1948). Por lo anterior, se requiere de trabajos futuros sobre los tejidos vasculares de las plantas-patrón y varetas que de manera sistemática y rigurosa corroboren la compatibilidad de los tejidos vasculares de las especies.
Conclusiones
La estrategia de propagación por injerto enchapado lateral es viable para P. rzedowskii, ya que se logra supervivencia mayor de 80 %. De igual manera se demuestra que el injerto interespecífico es posible y que el factor especie afecta la supervivencia a través del tiempo, la cual está asociada a la cercanía filogenética de las especies. La mayor supervivencia se obtuvo con plantas-patrón de P. pinceana, que forma un grupo monofilético con P. rzedowskii. Esta información es una iniciativa con una sólida base científica y biotecnológica para la propagación de la especie a mayor escala, de gran utilidad para su conservación ex situ y para ensayos futuros de mejoramiento genético.
Agradecimientos
Los autores agradecen al Consejo de la Investigación Científica de la UMSNH, por el financiamiento otorgado dentro del Programa de Investigación 2018-2019. A la comunidad de Dos Aguas, Michoacán, por las facilidades otorgadas durante el trabajo de campo y a los revisores anónimos que con sus comentarios enriquecieron el manuscrito sustancialmente.
REFERENCIAS
Aparicio-Rentería, A., Viveros-Viveros, H., & Rebolledo-Camacho, V. (2013). Huertos semilleros clonales: una alternativa para los programas de reforestación en Veracruz. Revista Mexicana de Ciencias Forestales, 4(20), 90‒97. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-11322013000600009 [ Links ]
Barchuk, A. H., & Díaz, M. P. (2000). Vigor de crecimiento y supervivencia de plantaciones de Aspidosperma quebracho-blanco y de Prosopis chilensis en el Chaco árido. Quebracho, 8, 17‒29. Retrieved from https://fcf.unse.edu.ar/archivos/quebracho/q8_02-Barchuk.pdf [ Links ]
Castro-Garibay, S. L., Villegas-Monter, A., & López-Upton, J. (2017). Anatomy of rootstocks and scions in four pine species. Forest Research, 6(3), 1‒6. doi: 10.4172/2168-9776.1000211 [ Links ]
Castilleja, S. P., Delgado, P., Sáenz-Romero, C., & Herrerías, D. Y. (2016). Reproductive success and inbreeding differ in fragmented populations of Pinus rzedowskii and Pinus ayacahuite var. veitchii, two endemic Mexican pines under threat. Forests, 7(8), 1‒17. doi: 10.3390/f7080178 [ Links ]
Climent, J. M., Prada, M. A., Gil, L. A., & Pardos, J. A. (1997). Increase of flowering in Pinus nigra Arn subsp salzmannii (Dunal) Franco by means of heteroplastic grafts. Annals of Forest Science, 54(2), 145‒153. doi: 10.1051/forest:19970202 [ Links ]
Darikova, Y. A., Vaganov, E. A., Kuznetsova, G. V., & Grachev, A. M. (2013). Changes in the anatomical structure of tree rings of rootstock and scion in the heterografts of Siberian pine. Trees, 27, 1621‒1631. doi: 10.1007/s00468-013-0909-6 [ Links ]
Delgado, P., Piñero, D., Chaos, A., Pérez‐Nasser, N., & Alvarez‐Buylla, E. R. (1999). High population differentiation and genetic variation in the endangered Mexican pine: Pinus rzedowskii (Pinaceae). American Journal of Botany, 86(5), 669‒676. doi: 10.2307/2656576 [ Links ]
Dyson, W. G. (1975). A note on dwarfing of Pinus patula grafts. Silvae Genetica, 24, 60‒61. Retrieved from https://www.thuenen.de/media/institute/fg/PDF/Silvae_Genetica/1975/Vol._24_Heft_2-3/24_2-3_60.pdf [ Links ]
Eckert, J. A., & Hall, D. B. (2006). Phylogeny, historical biogeography, and patterns of diversification for Pinus (Pinaceae): Phylogenetic tests of fossil-based hypotheses. Molecular Phylogenetics and Evolution, 40(1), 166‒182. doi: 10.1016/j.ympev.2006.03.009 [ Links ]
Farjon, A., & Styles, B. T. (1997). Pinus (Pinaceae). Flora Neotropica, 75, 1‒291. Retrieved from http://www.jstor.org/stable/4393881 [ Links ]
García, B. J. J. (2012). Análisis de supervivencia aplicado al estudio de la mortalidad en injertos de inchi (Caryodendron orinocense Karsten). Revista Científica UDO Agrícola, 12(4), 759‒769. Retrieved from https://dialnet.unirioja.es/servlet/articulo?codigo=6104314 [ Links ]
Gernandt, D. S., Geada, L. G., Ortiz, G. S., & Liston, A. (2005). Phylogeny and classification of Pinus. Taxon, 54(1), 29‒42. doi: 10.2307/25065300 [ Links ]
Goldschmidt, E. E. (2014). Plant grafting: new mechanisms, evolutionary implications. Frontiers in Plant Science, 5, 1‒9. doi: 10.3389/fpls.2014.00727 [ Links ]
Guadaño, C., & Mutke, S. (2016). Establecimiento de plantaciones clonales de Pinus pinea para la producción de piñón mediterráneo. Madrid, España: INIA. [ Links ]
Hartmann, H. T., Kester, D. E., Davies, F. T., & Geneve, R. L. (2002). Plant propagation. principles and practices (7th ed.). New Jersey, USA: Prentice Hall. [ Links ]
Le, C. T. (1997). Applied survival analysis. New York, USA: John Wiley and Sons. [ Links ]
López-Mata, L. (2013). The impact of seed extraction on the population dynamics of Pinus maximartinezii. Acta Oecologica, 49, 39‒44. doi: 10.1016/j.actao.2013.02.010 [ Links ]
Madrigal, X. S., & Caballero, D. M. (1969). Una nueva especie mexicana de Pinus. México: Instituto Nacional de Investigaciones Forestales. [ Links ]
Martínez-Ávalos, J., Sánchez-Castillo, C. G., Martínez-Gallegos, R., Sánchez-Ramos, G., Trejo, I., Mora-Olivo, A., & Alanis, E. (2015). Primer registro de Pinus pinceana (Pinaceae) para Tamaulipas: aspectos ecológicos y estructurales. Botanical Sciences, 93(2), 357‒360. doi: 10.17129/botsci.87 [ Links ]
Molina-Freaner, F., Delgado, P., Perez, N. N., Piñero, D., & Alvarez-Buylla, E. R. (2001). Do rare pines need different conservation strategies? Evidence from three Mexican species. Canadian Journal of Botany, 79(2), 131‒138. doi: 10.1139/b00-155 [ Links ]
Montes, J. R., Peláez, P., Willyard, A., Moreno-Letelier, A., Piñero, D., & Gernandt, D. S. (2019). Phylogenetics of Pinus subsection Cembroides Engelm. (Pinaceae) inferred from low-copy nuclear gene sequences. Systematic Botany, 44(3), 501‒518. doi: 10.1600/036364419X15620113920563 [ Links ]
Muñoz-Flores, H. J., Prieto-Ruiz, J. Á., Flores-García, A., Pineda-Ojeda, T., & Morales-González, E. (2013). Técnicas de injertado "enchapado lateral" y "fisura terminal" en Pinus pseudostrobus Lindl. México: INIFAP. Retrieved from https://www.academia.edu/31278206/T%C3%A9cnicas_de_injertado_enchapado_lateral_y_fisura_terminal_en_Pinus_pseudostrobus_Lindl [ Links ]
Pérez-Luna, A., Prieto-Ruíz, J. A., López-Upton, J., Carrillo-Parra, A., Wehenkel, Ch., Chávez-Simental, J. A., & Hernández-Díaz, J. C. (2019). Some factors involved in the success of side veneer grafting of Pinus engelmannii Carr. Forests, 10(2), 112. doi: 10.3390/f10020112 [ Links ]
Pérez-Luna, A., Wehenkel, C., Prieto-Ruíz, J. A., López-Upton, J., Solís-González, S., Chávez-Simental, J. A., & Hernández, J. C. (2020a). Grafting in conifers: a review. Pakistan Journal of Botany, 52(4), 1‒10. doi: 10.30848/PJB2020-4(10) [ Links ]
Pérez-Luna, A., Wehenkel, C., Prieto-Ruíz, J. A., López-Upton, J., Solís-González, S., & Hernández-Díaz, J. C. (2020b). Survival of side grafts with scions from pure species Pinus engelmannii Carr. and the P. engelmannii x P. arizonica Engelm. var. arizonica hybrid. PeerJ, 8:e8468. doi: 10.7717/peerj.8468 [ Links ]
Quiroz-Vázquez, R. I., López-Upton, J., Cetina-Alcalá, V. M., & Ángeles-Pérez, G. (2017). Capacidad reproductiva de Pinus pinceana Gordon en el límite sur de su distribución natural. Agrociencia, 51(1), 91‒104. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-31952017000100091 [ Links ]
R Development Core Team. (2016). R: A language and environment for statistical computing. Vienna, Austria: R Foundation fos Statistical Computing. [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). (2010). Norma Oficial Mexicana NOM-059, Protección ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-lista de especies en riesgo. México: Diario Oficial de la Federación. Retrieved from https://www.gob.mx/cms/uploads/attachment/file/134778/35.-_NORMA_OFICIAL_MEXICANA_NOM-059-SEMARNAT-2010.pdf [ Links ]
Sigala, R. J. A., González, T. M. A., & Prieto-Ruíz, J. A. (2015). Supervivencia en plantaciones de Pinus pseudostrobus Lindl. en función del sistema de producción y preacondicionamiento en vivero. Revista Mexicana de Ciencias Forestales, 6(30), 20‒31. doi: 10.29298/rmcf.v6i30.205 [ Links ]
Villaseñor, R. R., & Carrera, G. M. (1980). Tres ensayos de injertado en Pinus patula Schl. et Cham. Ciencia Forestal, 5(23), 21‒36. Retrieved from https://www.cabdirect.org/cabdirect/abstract/19820685496 [ Links ]
Recibido: 17 de Junio de 2020; Aprobado: 22 de Marzo de 2021
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
